
miR-26对TGF-β2诱导的人Tenon氏囊成纤维细胞迁移、细胞外基质蛋白表达的调控作用及其机制
2022-12-26 15:47李燕孟凯杜允宏陈斐刘文静鲍慧婧
山东医药 2022年35期
李燕,孟凯,杜允宏,陈斐,刘文静,鲍慧婧
1青岛大学附属泰安市中心医院眼科,山东泰安 271000;
2青岛大学附属泰安市中心医院肛肠外科
青光眼滤过性手术是目前治疗青光眼尤其是终末期青光眼的主要手段,但该手术方式存在较高的远期失败率,手术区滤过泡瘢痕形成是导致青光眼滤过性手术失败的主要原因[1]。转化生长因子β2(TGF-β2)是创伤后组织修复过程中关键的调控分子,可通过介导人Tenon氏囊成纤维细胞(HTFs)增殖、迁移并分泌大量细胞外基质,促进细胞纤维化进程,加速青光眼术后滤过泡瘢痕化,从而造成手术失败。miR-26是微小RNA(miRNA)的一种,位于人3号染色体3P21.3,由21~23个核苷酸组成,被证实能够影响多种肿瘤的发生发展[2-4]。近年来多项研究表明,miR-26在肝脏纤维化、心肌细胞纤维化、肾脏纤维化以及晶状体纤维化等多种纤维化疾病中发挥重要调控作用,已经成为进一步开发抗纤维增生药物的新靶点[5-8]。但是,目前关于miR-26在青光眼术后滤过泡瘢痕化中的研究比较少。2018年9月—2021年12月,本研究观察了miR-26对TGF-β2诱导后HTFs细胞迁移及细胞外基质(ECM)蛋白分泌的影响,并对其可能的作用机制进行探讨,为青光眼术后抗瘢痕化治疗提供新的靶点和理论基础。
1 材料与方法
1.1 材料DMEM陪养基、胎牛血清均购自美国Gibco公司,青—链霉素双抗购自美国Hyclone公司,胰蛋白酶购自上海实生生物技术有限公司,Lipo⁃fectamineTM2000试剂购自美国Invitrogen公司,逆转录试剂盒、qPCR试剂盒均购自大连宝生物科技有限公司,TGF-β2购自美国R&D Systems公司;角蛋白一抗、波形蛋白一抗、FITC荧光二抗、纤维连接蛋白(Fibronectin)一抗、Ⅰ型胶原蛋白(COLⅠ)一抗、CTGF一抗 均购 自美 国Santa Cruz公 司。miR-26的模拟物和2'甲氧基修饰的反义寡核苷酸序列根据Elbashir的设计原则进行设计,由上海吉玛制药有限公司合成,经BLAST查询,排除与其他基因同源。
1.2 HTFs的提取、培养及鉴定
1.2.1 HTFs的提取选择2018年9月—2021年10月于本院行青光眼滤过性手术的青光眼患者10例,平均年龄50.12岁,男女比例为4∶6。术中沿患者角巩缘剪开球结膜,取结膜下适当体积的Tenon氏囊组织,注意避免混入结膜上皮组织,置于无菌的生理盐水瓶中保存。本研究通过医院伦理委员会审核,患者均签署知情同意书。
1.2.2 HTFs的原代培养将无菌取材的Tenon氏囊组织置于培养皿中,PBS反复冲洗,加入少量含20%胎牛血清的DMEM培养液;用眼科剪将其剪成大小约0.5 mm³的组织块,置于35 mm2培养皿中,接种后加入含10%胎牛血清的DMEM培养基,置于恒温培养箱中培养。每天换液1次,组织块贴壁培养3~8 d后,倒置显微镜下可见细胞从组织边缘游离出,细胞形态不一,多呈长梭形、多边形或星状,可有2~5个细长状突起,细胞核大而居中;20 d左右细胞生长渐渐达融合状态。倒置显微镜下观察当细胞融合达到90%时,即可进行传代培养,细胞传代3次后,细胞生长趋于一致,细胞呈长梭形,大小不一,呈旋涡状或鱼群样分布,有一定的方向性;细胞体透亮,细胞核位于细胞中央,呈椭圆形,符合HTFs的形态学特征,见OSID码图1。
1.2.3 HTFs表型鉴定采用免疫荧光染色。取传至第4代的HTFs进行后续实验,6孔板中加入培养液,放入爬片,以4×104/mL进行接种,培养48 h后取出爬片,每孔加入PBS 1~2 mL漂洗3次;吸除PBS,用乙醇与冰醋酸(99∶1)的混合液固定细胞15 min,PBS漂洗。加入0.5% Triton X-100 1 mL,室温下孵育10 min,PBS漂洗;加入10%羊血清1 mL,室温下封闭30 min,吸除羊血清。滴加0.5%BSA稀释的单克隆兔抗人波形蛋白、角蛋白一抗(稀释比例均为1∶100),置于4℃湿盒孵育过夜,PBS漂洗;滴加FITC标记的0.5%BSA稀释二抗,室温孵育1 h,PBS漂洗。加入碘化丙啶(PI),37℃孵育10 min,封片剂封片,荧光显微镜下观察拍照。免疫荧光染色结果显示,波形蛋白在HTFs细胞质内呈阳性表达,表现为与细胞生长方向一致的束状绿色高荧光,角蛋白在HTFs细胞质内呈阴性表达,见OSID码图2;证实培养的细胞为成纤维细胞。
1.3 细胞分组及处理方法取对数期传代的HT⁃Fs,调整细胞密度为4×104/mL,每孔板2 mL接种到6孔板。将细胞随机分为空白组(不予转染)、TGFβ2组(不予转染)、TGF-β2+miR-26 mimics组(转染miR-26类似物)、TGF-β2+mimics NC组(转染miR-26阴性对照物)、TGF-β2+miR-26 inhibitors组(转染miR-26抑制物)、TGF-β2+inhibitors NC组(转染miR-26抑制物阴性对照物),按照LipofectamineTM2000转染试剂盒说明书进行操作;转染24 h后,空白组加入等体积含10%FBS的DMEM培养基,其余各组更换为含5 μg/L TGF-β2及10%FBS的DMEM培养基,培养48 h进行后续检测。
1.4 细胞迁移能力观察采用划痕实验。取各组细胞,吸除培养基,用200 μL无菌枪头在融合的细胞层上沿直尺作一划痕,PBS冲洗,置于37℃、5%CO2细胞培养箱中,培养48 h后将细胞置于倒置显微镜下拍照。采用Image J软件测量划痕宽度,细胞迁移率=(初始划痕宽度-48 h划痕宽度)/初始划痕宽度×100%。
1.5 细胞COLⅠ、Fibronectin蛋白检测采用Western blotting法。取各组细胞,使用RIPA试剂提取总蛋白,BCA法定量。加入5×蛋白上样缓冲液,100℃恒温水浴5 min,使蛋白充分变性。取变性蛋白40 μg行SDS⁃PAGE电泳,转至PVDF膜,5%脱脂奶粉封闭2 h。分别加入COLⅠ、Fibronectin、GAP⁃DH一抗(稀释比例均为1∶1 000),4℃孵育过夜;加入二抗(稀释比例均为1∶1 000),室温条件孵育1 h。采用Odyssey红外图像成像系统扫描成像,以GAPDH为内参,计算COLⅠ、Fibronectin蛋白相对表达量。
1.6 miR-26和CTGF的靶向作用观察采用生物信息学软件Target Scan筛选发现,CTGF基因下游的3′非翻译区存在miR-26的结合位点(见图1)。采用荧光素酶报告基因实验进行验证,CTGF的野生型(WT)和突变型(MUT)3'-UTR均由上海吉玛制药有限公司克隆到报告基因监测系统pmiR-RBReportTM质粒载体中。取对数期传代的HTFs,分为四部分,分别共转染WT-CTGF、MUT-CTGF 100 ng与miR-26 mimics、miR-26 NC载体50 nmol/L,严格按照LipofectamineTM2000说明书进行操作,共转染48 h后按照双荧光素酶活性检测试剂盒说明书操作,记录海肾荧光素酶与萤火虫萤光素酶的荧光值,以二者的比值表示荧光素酶相对活性。共转染miR-26 mimics+WT-CTGF的HTFs荧光素酶相对活性明显低于共转染miR-26 NC+WT-CTGF的HTFs(P<0.05),共 转 染miR-26 mimics+MUT-CTGF的HTFs与 共 转 染miR-26 NC+MUT-CTGF的HTFs荧光素酶相对活性比较差异无统计学意义(P>0.05)。
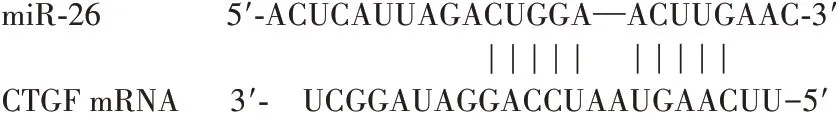
图1 Target Scan软件显示miR-26和CTGF存在结合位点
1.7 各组细胞CTGF mRNA及蛋白检测①CTGF mRNA:采用实时荧光定量PCR法。取各组细胞,采用TRIzol法提取总RNA,行逆转录反应。CTGF引物序列:上游引物5′-GCGGCTTACCGACTGGA-3′、下 游 引 物5′-AGGCGGCTCTGCTTCTC-3′,内 参GAPDH引 物 序 列:上 游 引 物5′-GTCGATGGC⁃TAGTCGTAGCATCGAT-3′、下 游 引 物5′-TGC⁃TAGCTGGCATGCCCGATCGAT-3′。PCR反应体系:模板1.0 μL,上、下游引物各1.0 μL,Master 12.5 μL,加双蒸水补充至25 μL。PCR反应过程:95℃预变性2 min,95℃变性15 s,60℃退火、延伸60 s,共40个循环,最后72℃延伸3 min。PCR完成后,采用2-ΔΔCt法计算CTGF mRNA相对表达量。②CTGF蛋白:参照1.5采用Western blotting法检测各组细胞CTGF蛋白相对表达量。
1.8 CTGF、miR-26共同作用对TGF-β2诱导的HTFs细胞迁移及COLⅠ、Fibronectin蛋白表达影响的观察
1.8.1 细胞分组处理参照Gen Bank数据库提供的CTGF基因序列,由上海吉玛制药技术有限公司设计合成CTGF-siRNA和阴性对照siRNA。将对数生长期的HTFs分为空白转染组(不予转染)、miR-26 inhibitor组(转染miR-26 inhibitor)、miR-26 inhibi⁃tors+si-NC组(转 染miR-26 inhibitor+阴 性 对 照siRNA)、miR-26 inhibitors+si-CTGF组(转染miR-26 inhibitor+CTGF-siRNA),严格按照LipofectamineTM2000说明书操作;转染24 h后,空白转染组加入等体积含10%FBS的DMEM培养基,其余各组更换为含5 μg/L TGF-β2及10%FBS的DMEM培养基,培养48 h进行后续检测。
1.8.2 细胞迁移能力参照“1.4”采用划痕实验检测各组细胞迁移率。
1.8.3 细胞COLⅠ、Fibronectin蛋白表达参照“1.5”采用Western blotting法检测各组细胞COLⅠ、Fibronectin蛋白相对表达量。
1.9 统计学方法采用SPSS17.0统计软件。计量资料采用Shapiro-Wilk正态性检验,呈正态分布以±s表示,多组间比较采用方差分析,组间比较采用独立样本t检验,重复测量数据采用重复测量的方差分析;非正态分布以M(P25,P75)表示,组间比较采用秩和检验。P<0.05为差异有统计学意义。
2 结果
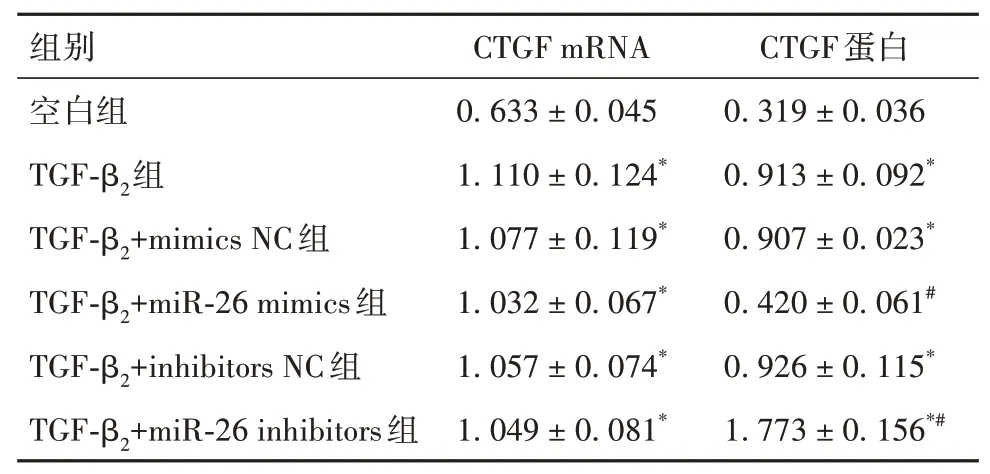
2.1 六组细胞迁移率及COLⅠ、Fibronectin蛋白表达比较空白组、TGF-β2+miR-26 mimics组 表1 六组细胞迁移率及COLⅠ、Fibronectin蛋白相对表达量比较(±s) 表1 六组细胞迁移率及COLⅠ、Fibronectin蛋白相对表达量比较(±s) 注:与空白组、TGF-β2+miR-26 mimics组比较,*P<0.05;与TGF-β2组、TGF-β2+mimics NC组、TGF-β2+inhibitors NC组比较,#P<0.05。 ? 2.2 六组细胞CTGF mRNA和蛋白表达比较CT⁃GF mRNA相对表达量:空白组 表2 六组细胞CTGF mRNA和蛋白相对表达量比较(±s) 表2 六组细胞CTGF mRNA和蛋白相对表达量比较(±s) 注:与空白组比较,*P<0.05;与TGF-β2组、TGF-β2+mimics NC组、TGF-β2+inhibitors NC组比较,#P<0.05。 ? 2.3 四组细胞迁移率及COLⅠ、Fibronectin蛋白表达比较TGF-β2+miR-26 inhibitors+si-CTGF组<空白 转染 组 表3 四组细胞迁移率及COLⅠ、Fibronectin蛋白相对表达量比较(±s) 表3 四组细胞迁移率及COLⅠ、Fibronectin蛋白相对表达量比较(±s) 注:与空白转染组比较,*P<0.05;与TGF-β2+miR-26 inhibitors组、TGF-β2+miR-26 inhibitors+si-NC组比较,#P<0.05。 ? 青光眼滤过性手术后瘢痕过度增生导致的滤过泡包裹是青光手术失败的主要原因,目前临床上通常采用的5-FU、MMC、环孢霉素A(CsA)等均是通过抑制成纤维细胞增殖而发挥作用,取得了一定疗效,但是由于这些药物对眼部组织的作用具有非特异性,常常导致滤过泡渗漏、低眼压性黄斑病变等一系列并发症的发生。因此,阐明青光眼术后滤过通道瘢痕化的机制,并积极探索特异性更强、更安全有效的新靶点一直是眼科青光眼领域研究的热点。 TGF-β2是创伤后组织修复过程中关键的调控分子,与青光眼术后滤过泡瘢痕形成关系密切,主要通过介导HTFs细胞增殖、迁移并分泌大量细胞外基质以促进细胞纤维化进程。过去曾认为,抗TGF抗体是青光眼术后抗纤维化治疗的方法之一,有多项研究尝试通过阻断TGF-β2表达来有效抑制滤过泡瘢痕,从而提高手术成功率,如ilomastat、Decorin、ASON等。但事实证实这种方法具有双刃剑的作用,阻断TGF在缓解了其促纤维化效应的同时,也抑制了其正面的抗炎效应。因此,临床亟需寻找一种靶向作用更强、疗效更高、不良反应更小的药物用于抑制青光眼滤过性手术后瘢痕的形成。 miRNA是真核生物体内的一类长22~24个核苷酸的单链小分子RNA,是非编码RNA的一种,具有高度的基因保守性。miRNA对基因表达的调控位于转录后水平,通过完全或者不完全互补配对,特异性结合到靶mRNA的3′非翻译区,引起靶mRNA的降解或者翻译抑制,从而抑制靶蛋白表达。近年多项研究表明,miRNA与青光眼的发病、进展有关[9-12]。miR-26作为miRNA的一种,最早被发现在多种肿瘤的发生和转移方面具有重要作用,近年来越多的研究证实miR-26在多种器官纤维化过程中发挥重要作用。ZHANG等[7]研究发现,在单侧输尿管梗阻(UUO)小鼠的肾脏、肌肉和外泌体中,miR-26a表达减少,小鼠骨骼肌注射miR-26a可减弱UUO小鼠的肾纤维化。CHEN等[8]研究认为,miR-26a和miR-26b在体内外均能显著抑制晶状体上皮细胞的增殖、迁移和晶状体纤维化。但是,关于miR-26在青光眼术后滤过通道瘢痕化过程中的作用目前尚无相关研究。 成纤维细胞向创面的迁移移行是创面愈合的重要步骤,而细胞外基质蛋白的过量分泌和合成是创伤后瘢痕形成的关键因素。本研究通过检测COLⅠ和Fibronectin蛋白表达来评价miR-26对ECM蛋白分泌的影响;结果显示,与对照组比较,TGF-β2组细胞迁移率及COLⅠ、Fibronectin蛋白表达均升高,提示TGF-β2可诱导HTFs细胞迁移及ECM蛋白分泌;与TGF-β2组比较,miR-26 mimics组COLⅠ和Fibronectin蛋白表达下降,miR-26 inhibitors组COLⅠ和Fibronectin蛋白表达提高,提示miR-26能抑制TGF-β2诱导的ECM蛋白分泌,而敲低miR-26可以促进ECM分泌。KOGA等[13]研究表明,在糖尿病肾病模型中,miR-26可以抑制TGF-β诱导的ECM蛋白表达,与本研究结果一致。 目前已有多项研究表明,miR-26的抗纤维化作用是通过抑制CTGF表达而实现的。YANO等[14]研究发现,细胞暴露于电离辐射后,miR-26a表达下调,miR-26a在转录后水平负向调控CTGF表达,可减轻辐射后引起的纤维化作用。LI等[15]研究发现,miR-26a-5p过表达可通过降低支气管肺泡灌洗液中总蛋白、中性粒细胞、淋巴细胞计数以及肿瘤坏死因子α(TNF-α)、白细胞介素1β(IL-1β)、IL-6、丙二醛(MDA)和髓过氧化物酶(MPO)表达,缓解脂多糖诱导的急性肺损伤小鼠的炎症反应,而CTGF过表达可以逆转miR-26a-5p对细胞凋亡和炎症反应的影响。本研究采用生物信息学软件Target Scan对CT⁃GF基因的3′非翻译区进行了潜在靶点的筛选,发现CTGF基因下游的3′非翻译区存在miR-26的结合位点,此结合位点位于人CTGF基因mRNA的3′非翻译区652~659 bp处;双荧光素酶报告基因实验也证实miR-26可与CTGF靶向结合,证实CTGF是miR-26的下游靶点。本研究结果显示,miR-26过表达后HTFs中的CTGF蛋白表达降低,抑制miR-26表达后细胞内CTGF蛋白表达增高,但miR-26过表达/抑制对细胞CTGF mRNA表达无明显影响。由此我们推测,miR-26可能在转录后水平直接靶向结合CTGF mRNA而负调控CTGF蛋白表达。本研究在转染miR-26 inhibitors质粒的基础上采用RNA干扰技术抑制细胞内CTGF基因表达,结果发现与TGF-β2+miR-26 inhibitors组比较,TGF-β2+miR-26 inhibitors+si-CTGF组细胞迁移能力以及促ECM蛋白分泌能力降低,表明敲除CTGF基因能够逆转miR-26下调对TGF-β2诱导的HTFs细胞迁移和ECM蛋白表达的影响。 综 上 所 述,miR-26可 以 抑 制TGF-β2诱 导 后HFTs细胞迁移及ECM蛋白合成,该作用可能是通过转录后水平靶向CTGF来实现的,调控miR-26表达有望为青光眼术后抗瘢痕化治疗提供新的思路。

3 讨论
猜你喜欢
中老年保健(2022年3期)2022-08-24天津医科大学学报(2021年4期)2021-08-21眼科学报(2021年6期)2021-07-18中日友好医院学报(2021年1期)2021-04-14山东医药(2020年9期)2020-05-20中医眼耳鼻喉杂志(2019年3期)2019-04-13浙江大学学报(工学版)(2016年2期)2016-06-05烟草科技(2015年8期)2015-12-20中国医药导报(2015年27期)2015-02-28郑州大学学报(医学版)(2015年1期)2015-02-27
