
香蕉UEV基因家族鉴定及其表达分析
2022-12-22 11:51武爱龙沈一跃郑佳明吴建阳
中国南方果树 2022年6期
何 冰,武爱龙,沈一跃,郑佳明,吴建阳
(1 湛江幼儿师范专科学校,广东湛江,524037;2 岭南师范学院基础教育学院,广东湛江524037)
香蕉是世界四大水果之一[1],主要产区为非洲、拉丁美洲和亚洲,全球117个香蕉生产国家和地区中,中国香蕉种植面积排名第二,仅次于印度[2-3]。中国香蕉主产区为台湾、福建、海南、云南、广西、广东等省(区),其中广东省产量最大。香蕉是呼吸跃变型果实,果实一般绿硬期采收,后熟食用[4]。
泛素是一种高度保守的蛋白,通过和其他蛋白共价结合发挥作用。泛素结合酶(Ubiquitin-conjugating enzymes,E2s)在DNA修复、核糖体合成、细胞周期调控等多种真核生物代谢过程中发挥重要作用[5-6]。所有E2s蛋白都有UBC保守结构域,里面含有1个半胱氨酸活性位点[7-8]。泛素结合酶变体蛋白(Ubiquitin-conjugating E2 enzyme variant,UEV)[9],又被称为类泛素结合酶蛋白家族,其结构和序列都与E2s相似,但没有半胱氨酸催化位点,所以该蛋白没有结合泛素的活性,但由于UBC可以彼此相互作用,形成同二聚体或异二聚体[10],进而影响这些UBC的活性和底物特异性,而UEV与UBC有非常高的同源性,因此UEV可以与某些UBC形成异二聚体,并对这些特定的UBC的泛素化作用进行负性或正性调控,因此UEV蛋白也可能参与对蛋白泛素化的调控[11]。
UEV参与激素信号、DNA 损伤和非生物胁迫等多种生物学功能[12],MMS2是酵母芽细胞中第一个被鉴定的UEV1基因,参与酵母DNA损伤[13]。拟南芥中4个UEV1基因都可以补充酵母mms2缺失突变体,UEV1D 缺失减弱了植株对DNA损伤剂的耐受性[14]。水稻OsUEV1s的表达可以让酵母mms2突变体不被DNA损伤剂致死[15]。UEV基因与果实成熟的关系研究在香蕉中未见报道。本研究从香蕉基因组中首次鉴定出VUEV基因家族所有成员,分析其理化性质、亚细胞定位、基因结构、保守基序和进化关系,探析其在香蕉后熟过程的表达模式,以期为研究UEV基因在香蕉后熟过程中的作用奠定基础。
1 材料与方法
1.1 材料
在中国热带农业科学院南亚热带作物研究所香蕉园内,采摘七至八成熟香蕉,带回实验室分成单果指,用清水冲洗干净,挑选外观一致的果实用500 mg/L施保功处理3 min后晾干,待其自然成熟,采摘当天标记为0 d,分别在采后0、1、4、7、17 d等5个时间点取样备用(见图1)。

图1 香蕉采后不同时间果实外观变化
1.2 拟南芥和香蕉UEV基因序列获取
先利用TAIR数据库(http://www.arabidopsis.org/index.jsp)获得拟南芥所有UEV基因序列,接着通过拟南芥UEV序列在香蕉基因组数据库(https://banana-genome-hub.southgreen.fr/)中进行BLAST比对,再将获得的序列放在NCBI中进行BLAST比对,最终获得香蕉UEV基因序列。
1.3 香蕉UEV基因家族蛋白基本理化性质
利用ExPASy在线软件获得UEV蛋白的氨基酸数目、分子量、等电点、亲水指数和不稳定系数,利用SOPMA在线分析软件对UEV蛋白的二级结构进行分析,利用CELLO在线软件对UEV蛋白亚细胞定位进行分析。
1.4 香蕉UEV家族基因结构、保守基序和进化树分析
利用MEME在线软件对UEV蛋白的保守基序进行分析,基序的最大数目设置为10。利用TBtools软件[16]对UEV基因结构进行分析。利用ClustalX进行氨基酸序列比对,利用MEGA 6.0的邻接法对UEV蛋白家族的进化树进行构建。
1.5 香蕉MaUEV荧光定量表达分析
香蕉总RNA利用植物RNA提取试剂盒提取(华越洋),cDNA利用TaKaRa公司的M-MLV 逆转录酶合成。实时荧光定量PCR在RoChe Light Cycler 480Ⅱ仪器上进行,利用TaRaKa公司荧光定量PCR试剂盒进行反应,反应体系20 μL,反应程序按说明书进行,每个反应重复3 次。根据转录组测序结果查找在采后成熟过程中有表达差异的UEV基因,共计10个,依据其基因序列,利用Primer Premier 5.0软件设计荧光定量引物,引物序列见表1,以香蕉Actin基因为内参,香蕉UEV基因的相对表达量采用2-ΔΔCT方法计算。

表1 香蕉UEV引物
2 结果与分析
2.1UEV基因家族成员鉴定及理化性质分析
试验结果看出,在香蕉基因组中共鉴定UEV基因13个,根据它们在染色体上的位置分别命名为MaUEV1~MaUEV13,MaUEV基因分布在1、3、4、5、7、9、10、11号染色体上,其中9号染色体上分布的UEV基因数量最多,为3个,1、3、4号染色体上分布的UEV基因数量次之,为2个。MaUEV基因开放阅读框介于471~1 637 bp,编码氨基酸数量127 ~495个,分子量14 212.34~56 209.83 Da,13个MaUEV基因的等电点均小于7;8个MaUEV基因为稳定蛋白,其不稳定指数均小于40,5个MaUEV基因为不稳定蛋白,其不稳定指数均大于40;13个MaUEV基因的总平均疏水指数均小于0,为亲水性蛋白(见表2)。

表2 香蕉UEV基因家族成员理化性质
2.2 UEV蛋白家族二级结构预测及亚细胞定位分析
试验结果看出,13个MaUEV蛋白均由无规则卷曲、扩展链结构、β-转角、α-螺旋组成,但不同基因编码蛋白的二级结构所占比例差异较大,β-转角在所有基因占比最低。其中MaUEV1、MaUEV2、MaUEV7、MaUEV13等4个蛋白中无规则卷曲所占比例最大,扩展链结构次之,α-螺旋再次之;MaUEV3、MaUEV4、MaUEV8、MaUEV12等4个蛋白中α-螺旋所占比例最大,无规则卷曲次之,扩展链结构再次之;MaUEV5、MaUEV6、MaUEV9、MaUEV10、MaUEV11等5个蛋白中无规则卷曲所占比例最大,α-螺旋次之,扩展链结构再次之。除MaUEV3蛋白定位在叶绿体、细胞质和细胞核外,其他基因编码蛋白都定位在细胞核(见表3)。
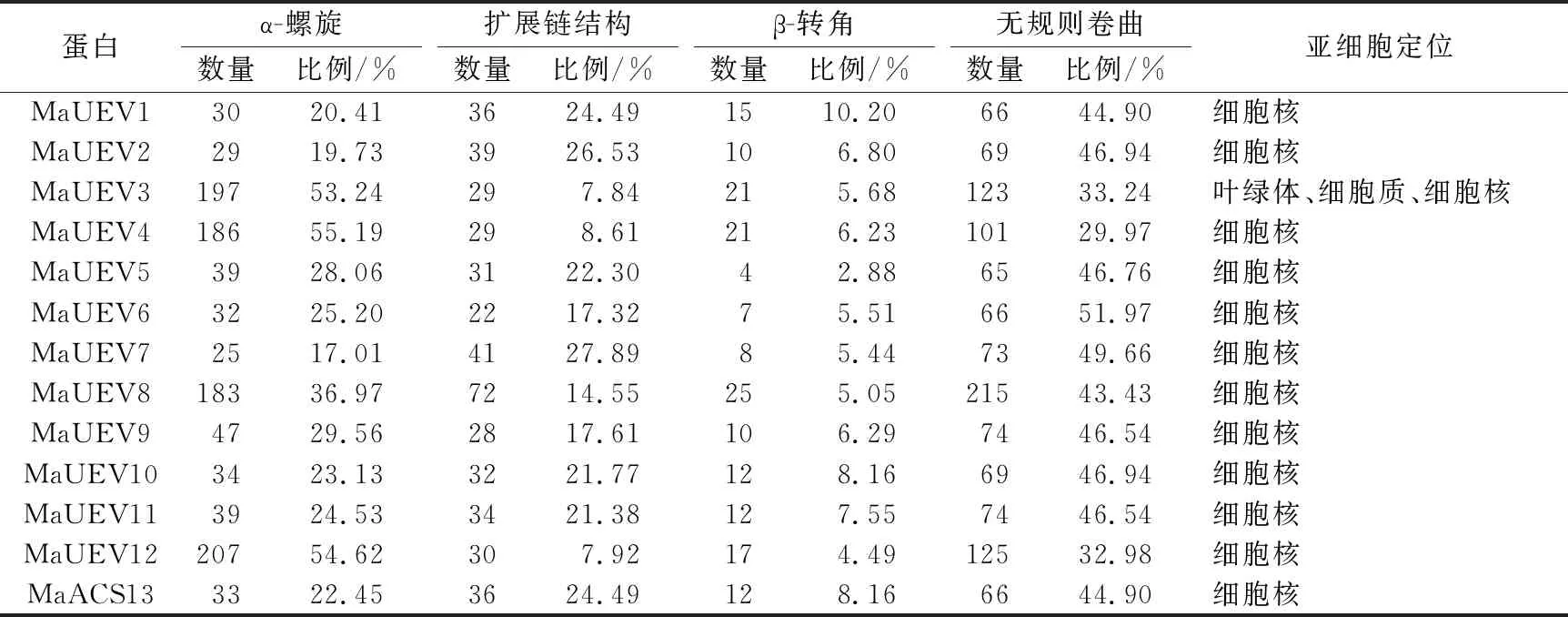
表3 香蕉UEV蛋白家族二级结构及亚细胞定位
2.3UEV基因家族结构、保守基序和进化分析
试验结果看出,除了MaUEV4无内含子外,其余12个MaUEV基因均由内含子和外显子组成,内含子数量介于1~12之间,外显子数量介于1~13之间。其中,MaUEV8基因内含子和外显子数量最多,有12个内含子和13个外显子(见图2)。利用MEME软件对UEV基因保守基序分析发现,MaUEV8只有1个保守基序,MaUEV1、MaUEV2、MaUEV7、MaUEV13具有3个相同的基序,MaUEV3和MaUEV12具有6个相同的基序,MaUEV5和MaUEV6具有1个相同的基序,MaUEV9和MaUEV11具有4个相同的基序(见图3)。构建了香蕉、拟南芥、水稻UEV的进化树,香蕉UEV与水稻的亲缘关系更近,且MaUEV1和MaUEV13、MaUEV9和MaUEV11、MaUEV3和MaUEV12聚类在一起,这与它们基因结构和保守基序数量相同(见图4)。
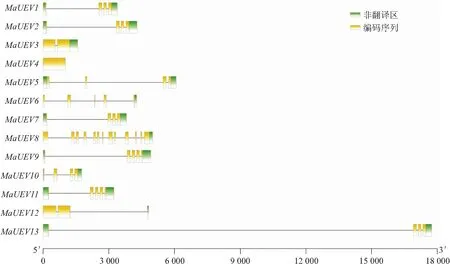
图2 香蕉UEV家族基因结构

图3 香蕉UEV基因家族保守基序

图4 香蕉、拟南芥和水稻UEV蛋白系统进化树
2.4 香蕉后熟过程中UEV基因表达分析
试验结果看出,香蕉后熟过程中,UEV基因家族成员表达趋势差异较大。MaUEV2、MaUEV5、MaUEV7、MaUEV10、MaUEV12等5个基因在采后0~7 d表达差异不大且稍有下调,但是在采后17 d显著上调。MaUEV3在采后0~4 d表达量稍有增加,在采后4~17 d稍有下降。MaUEV6和MaUEV9在后熟过程中逐渐下降,MaUEV6表达量变化不大,采后17 d的表达量下降至最低,仅为采后0 d的63%;而MaUEV9显著下降,采后17 d的表达量下降至最低,是0 d的32%。MaUEV11在采后0~4 d表达量逐渐下降,采后4~17 d表达量逐渐增加。MaUEV13基因在后熟过程中逐渐增加,采后17 d表达量达到峰值,约为采后0 d表达量的3倍(见图5和图6)。
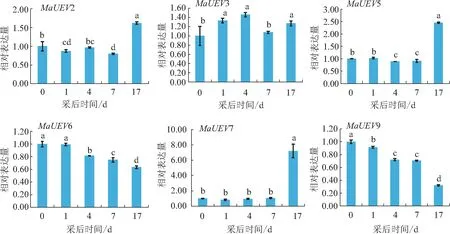
注:不同小写字母表示差异显著,p<0.05。图6同。

图6 香蕉后熟过程中4个MaUEV基因荧光定量表达比较
3 结论与讨论
UEV基因参与激素信号、DNA损伤和非生物胁迫等多种生物学功能,在番茄、藻类、苔藓、山楂等植物中都有报道[17],玉米[18]和可可[19]中有UEV基因6个,拟南芥中有8个[20]。本研究在香蕉中共鉴定UEV基因13个,MaUEV基因编码区域471~1 637 bp,编码氨基酸数量127~495个,分子量14 212.34~56 209.83 Da,等电点均小于7;8个MaUEV基因编码蛋白为稳定蛋白,5个MaUEV基因编码蛋白为不稳定蛋白;13个MaUEV基因编码蛋白均为亲水性蛋白,其编码蛋白均由α-螺旋、β-转角、扩展链结构、无规则卷曲组成;除MaUEV3定位在叶绿体、细胞质和细胞核外,其他基因都定位在细胞核。前人对UEV基因结构分析发现,番茄UEV基因外显子数量1~5个[12],可可UEV基因外显子数量1~9个[19],本研究发现香蕉中MaUEV基因外显子数量1~13个。
UEV是一类在结构和功能上都非常保守的蛋白,尽管没有半胱氨酸催化位点,其结构和序列都与E2s相似,UEV蛋白也可能参与对蛋白泛素化的调控。在拟南芥中COP10属于UEV蛋白,该蛋白在植物光形态建成中发挥重要作用[21]。UEV蛋白在香蕉中的作用还未见报道。本研究在香蕉后熟过程中分析了UEV基因家族的表达情况,根据其表达模式可以分为5种类型。第一类基因在采后0~7 d表达差异不大且稍有下降,采后17 d显著上调,包括MaUEV2、MaUEV5、MaUEV7、MaUEV10、MaUEV12等5个基因。第二类基因表达量先增加后下降,但变化幅度不大,MaUEV3属于这种类型。第三类基因在整个后熟过程中呈下调表达,MaUEV6和MaUEV9属于这种类型,但MaUEV6表达量变化不大,MaUEV9显著下调。第四类基因表达量先下降后增加,但变化幅度不大,MaUEV11属于这种类型。第五类基因表达量在整个后熟过程中逐渐增加,并在成熟时显著上调,MaUEV13属于这种类型。鉴于MaUEV9 和MaUEV13在香蕉后熟过程中表达差异显著且表达趋势相反,推测不同MaUEV基因在香蕉后熟过程中作用不同。
猜你喜欢
福建农林大学学报(自然科学版)(2022年4期)2022-11-01
电子科技大学学报(2022年5期)2022-10-29
中国生物化学与分子生物学报(2022年4期)2022-09-07
生物化学与生物物理进展(2022年8期)2022-08-20
南方医科大学学报(2022年3期)2022-04-13
南京医科大学学报(自然科学版)(2021年8期)2021-10-19
心血管病学进展(2021年8期)2021-09-13
湖北农业科学(2020年24期)2021-01-21
中国生殖健康(2020年4期)2021-01-18
三农资讯半月报(2020年15期)2020-08-25
