
2022 年鄂西地区PRRSV ORF5 基因遗传进化分析
2022-12-09 10:07:04欧阳艳刘发志王孝忠李祚丹熊同舟杜建丰
中国动物检疫 2022年12期
欧阳艳,刘发志,王孝忠,李祚丹,熊同舟,杜建丰
(1.湖北三峡职业技术学院,湖北宜昌 443000;2.宜昌市动物疫病预防控制中心,湖北宜昌 443000;3.宜昌正大畜牧有限公司,湖北宜昌 443000)
猪繁殖与呼吸综合征病毒(PRRSV)是引发猪繁殖与呼吸综合征(PRRS)的病原。目前,在全球范围内,除瑞士等少数国家没有暴发PRRS 疫情外,其他国家的养猪业都深受其害[1]。猪感染PRRSV 后表现厌食、发热、耳发绀、流鼻涕等症状,母猪感染后在怀孕110 d 左右可发生流产,产弱胎、死胎或木乃伊胎,产出的弱胎也会很快死亡[2-4]。
PRRSV 属于RNA 病毒,其结构蛋白GP5 是重要的保护性抗原蛋白,具有高度变异性,因此GP5 蛋白编码基因开放阅读框ORF5 常被用于遗传变异分析[5]。为了解鄂西地区PRRSV 的遗传变异情况,本研究对2022年采自该地区的672份猪血液、肺脏组织等样品进行实时荧光RT-PCR 检测,并对部分阳性样品进行ORF5 基因序列遗传进化分析,以期为鄂西地区PRRS 防控提供参考。
1 材料和方法
1.1 材料
1.1.1 主要试剂 PRRSV 通用型实时荧光RTPCR 检测试剂盒,购自元亨生物药业有限公司;DNA/RNA核酸提取试剂盒,购自天隆科技有限公司。
1.1.2 主要仪器设备 高速组织匀浆机(TissueLyser LT),德国凯杰公司产品;涡旋震荡仪(Mx-S)、赛洛捷克公司产品;高速冷冻离心机(Centriguge 5417R),德国艾本德公司产品;Ⅱ级生物安全柜(AC2-4S1),新加坡艺思高科技有限公司产品;荧光定量 PCR 扩增仪(LightCycler 96),德国罗氏诊断公司产品;生化培养箱(SPX-250B-Z),上海博讯实验有限公司医疗设备厂产品;医用冷藏箱(HYC-940),青岛海尔生物医疗股份有限公司产品;核酸提取仪(NP968),西安天隆科技有限公司产品。
1.2 方法
1.2.1 病料采集和处理 在鄂西地区采集疑似PRRS 发病猪血液、肺脏组织等样品共672 份,其中伍家岗区、夷陵区、兴山县、长阳自治县(简称长治县)、猇亭区、五峰自治县(简称五峰县)、恩施市7 个县(市、区)的血液、肺组织样品采自小规模育肥猪场,枝江市的血液、肺组织样品采自规模化种猪场。组织样品剪碎混匀后,加生理盐水充分匀浆,高速离心后,取上清液用于RNA 提取;血液样品采集后先放置在37 ℃生化培养箱中2 h,然后转移至4 ℃冰箱过夜,待血清析出后无菌分装,用于RNA 提取;抗凝血样品混匀后直接用于RNA提取。
1.2.2 引物设计与合成 使用欧阳艳等[6]设计的针对Nsp2基因的引物和1 对检测并扩增完整ORF5 基因序列的引物(表1)进行扩增。

表1 本研究使用的引物
1.2.3 RNA 提取与实时荧光RT-PCR 检测 用DNA/RNA 核酸提取试剂盒提取病毒总RNA,再使用实时荧光RT-PCR 检测试剂盒进行反转录。PCR 反应体系25.0 μL:无菌无核酸酶水5.0 μL、RT-PCR 反应液12.5 μL、酶混合液1.0 μL、荧光探针4.5 μL、模板RNA 2.5 μL。PCR 反应程序:42 ℃5 min,95 ℃变性10 s;95 ℃ 5 s,60 ℃ 35 s,40个循环,在每个循环第二步(60 ℃ 35 s)收集荧光信号。报告基团FAM,淬灭基团NONE。结束后根据样品Ct 大小及扩增曲线形成情况判定结果[7-8]。
1.2.4 ORF5 基因扩增与测序 将阳性样品用病毒核酸提取试剂盒提取RNA 后进行RT-PCR 扩增,将反应产物送擎科生物工程有限公司进行序列测定。
1.2.5 ORF5 基因遗传进化分析 应用DNAstar软件对测序获取的ORF5 基因序列与参考毒株序列进行比对分析,利用Clustal 2.1 和Mega-7.0 软件,基于Neighbor-Joining 运算法,绘制遗传进化树[6]。参考毒株的背景信息见表2。
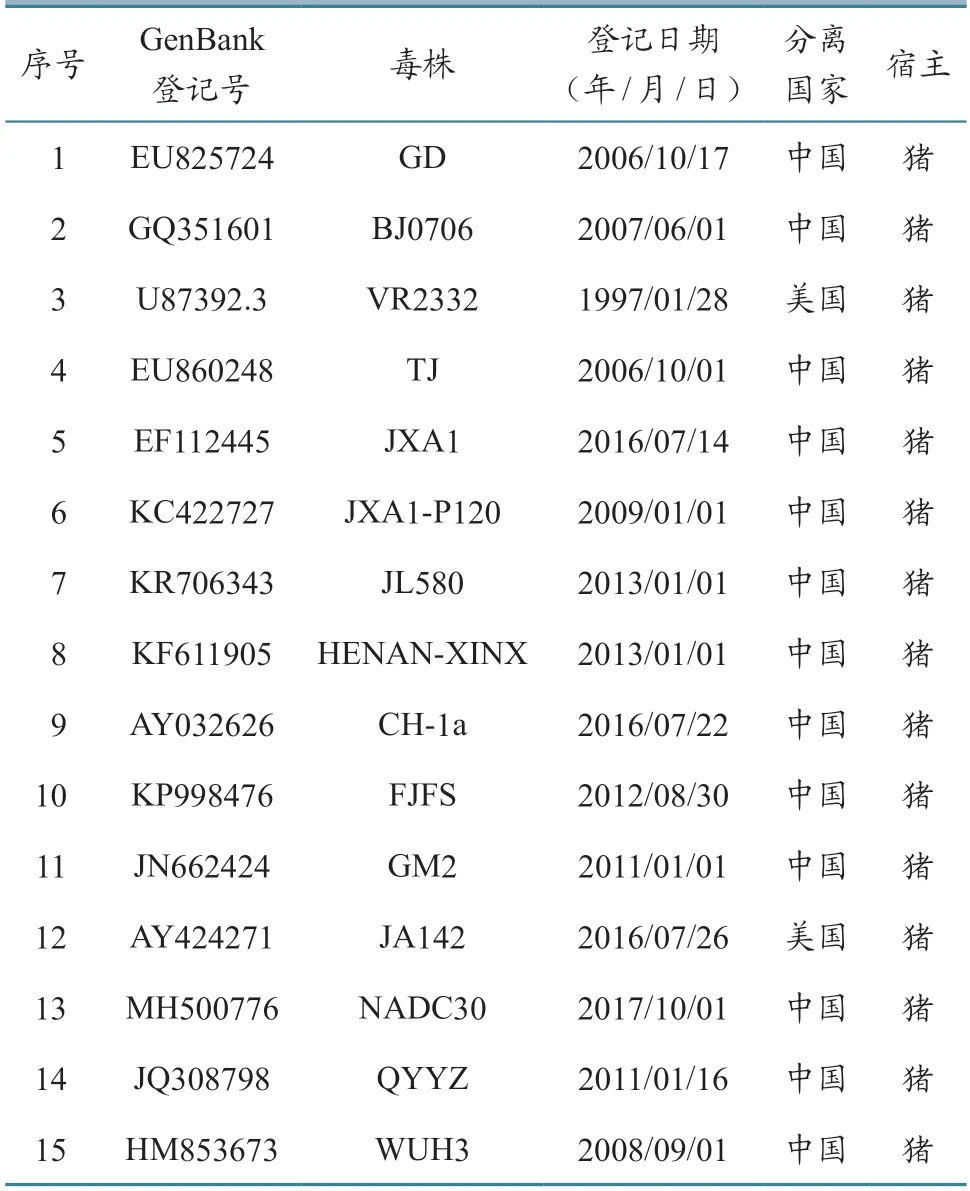
表2 参考毒株信息
2 结果与分析
2.1 临床样品RT-PCR 检测
2022 年,共收集检测来自鄂西地区8 个县(市、区)的血液、肺组织等样品672 份,使用PRRSV 实时荧光 RT-PCR 试剂盒检测。结果(表3、图1)显示:33 份样品为PRRSV 阳性,阳性检出率为5%;使用欧阳艳等[6]设计的针对Nsp2基因引物进行分类,其中高致病性毒株(HP-PRRSV)23 份,占69.7%,分布于伍家岗区、夷陵区、兴山县、长阳县、猇亭区等5 个县(区),是鄂西地区主要流行毒株;类NADC30 毒株(NADC30-like PRRSV)10 份,占30.3%,分布于兴山县、长阳县、枝江市、猇亭区等4 个县(市、区),是次要流行毒株。
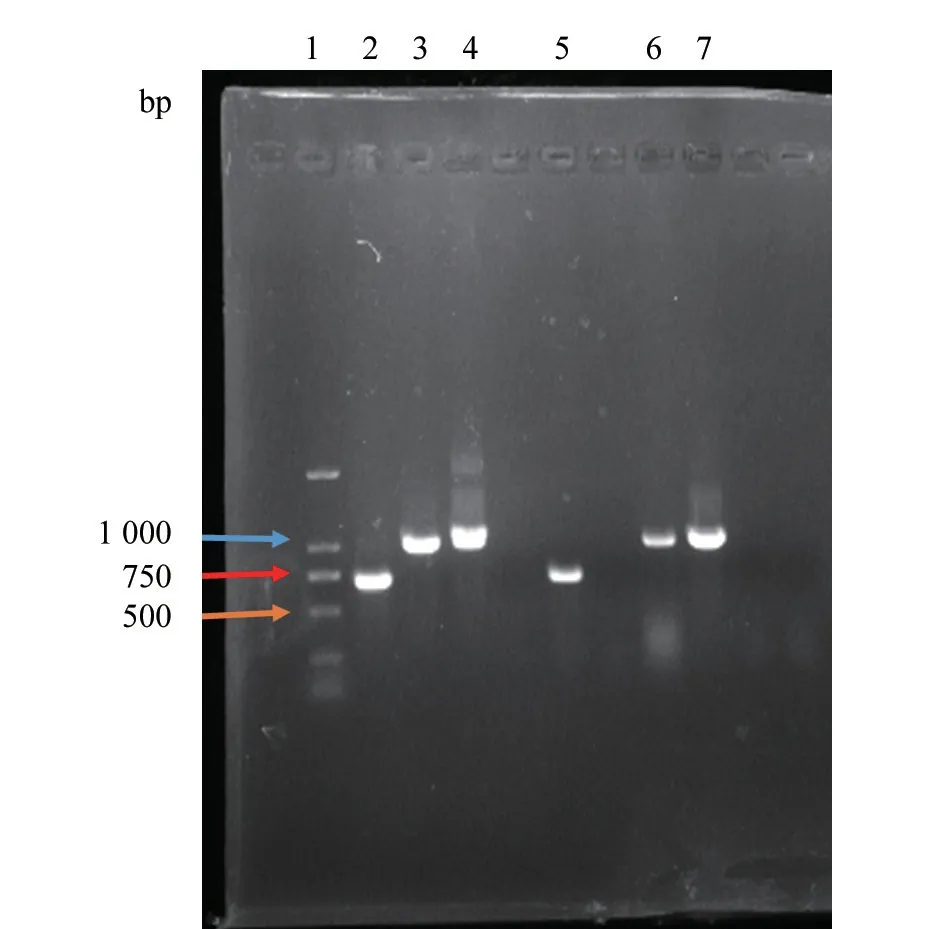
图1 PRRSV Nsp2 基因部分片段扩增结果
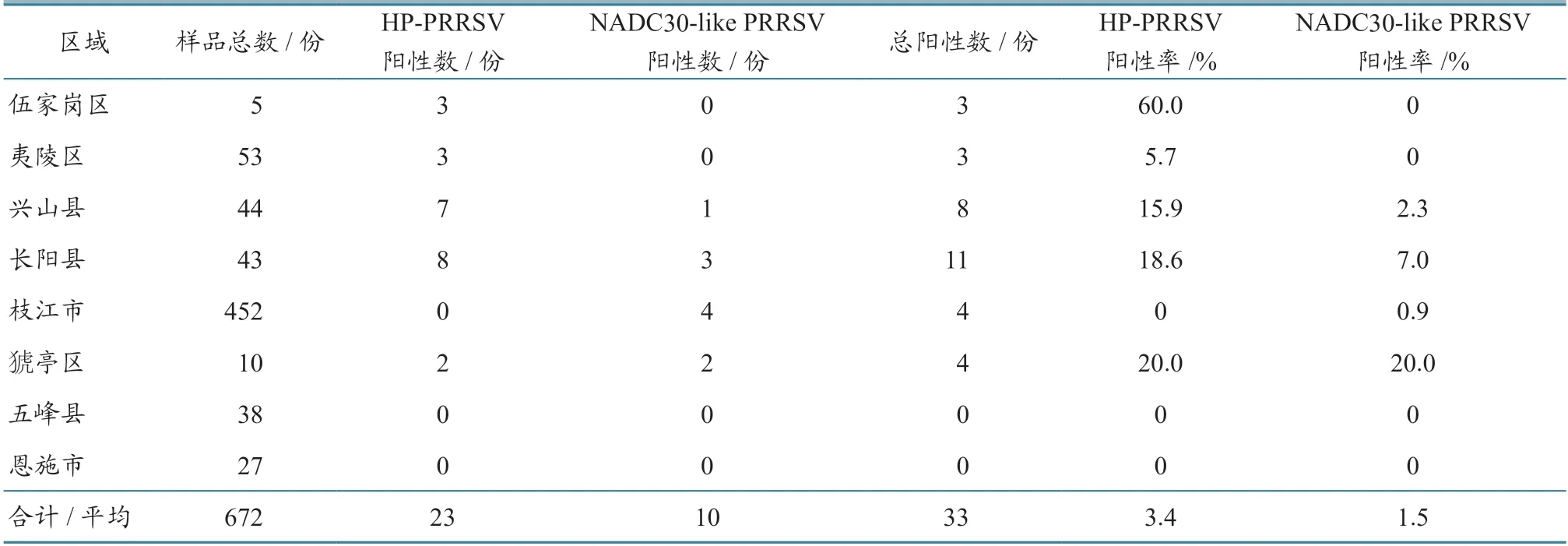
表3 2022 年病料检测结果
2.2 ORF5 基因同源性分析
本研究共扩增获得4 份阳性样品(2022YCLPFA、2022YCLPFA1、2022YCYZD、2022 YCZDBLZ)ORF5基因序列,其中2022YCLPFA、2022YCLPFA1采自长阳县,2022YCYZD采自夷陵区,2022YCZDBLZ 采 自枝江市。与参考毒株序列经同源性比对分析发现:4 份阳性样品间的ORF5 基因序列同源性为81.4%~98.5%,其与HP-PRRSV代表株JXA1同源性为80.3%~95.9%,与NADC30-like PRRSV代表株HENAN-XINX 同源性为80.8%~81.6%,与经典PRRSV 代表株VR-2332 同源性为80.3%~84.7%(图2)。

图2 ORF5 基因序列之间同源性分析结果
2.3 ORF5 序列遗传进化树分析
由图3可知:2022YCLPFA(长阳)、2022YCLPFA1(长阳)、2022YCYZD(夷陵)3份阳性样品ORF5 序列与JXA1、TJ-M 在同一分支(Lineage 8.7),属于HP-PRRSV;2022YCZDBLZ(枝江)样 品 与ZJ1407、JL580、HENAN-XINX、NADC30LIKEXD-HN-004 遗传关系较近,在同一分支(Lineage 1.9),属于类NADC30 毒株。

图3 PRRSV ORF5 基因进化树
2.4 GP5 蛋白氨基酸序列分析
经序列比对分析,GP5 蛋白氨基酸序列高变区,在诱骗表位(27~30 aa)出现29G →29C;在中和表位,HP-PRRSV 毒株出现氨基酸替换39A →39T,类NADC30 毒株出现38A →38T(图4)。
3 讨论
本研究共检测了来自鄂西地区8 个县(市、区)的672 份病料样品,发现PRRSV 阳性检出率为5%。其中:HP-PRRSV 阳性样品占69.7%,来自于伍家岗区、夷陵区、兴山县、长阳县、猇亭区等5 个县(市、区),可能是鄂西地区的主要流行毒株;类NADC30 阳性样品占30.3%,来自于兴山县、长阳县、枝江市、猇亭区等4 个县(市、区),可能是鄂西地区的次要流行毒株。从上述结果可以看出,长阳县、兴山县、猇亭区等地存在2 个毒株的混合流行,因此各场疫苗接种时要先进行检测,一定要选择与本场流行毒株匹配的疫苗,否则会导致免疫失败。
本研究获得4 份阳性样品的ORF5 基因序列同源性为81.4%~98.5%,其与HP-PRRSV代表株JXA1同源性为80.3%~95.9%,与NADC30-like PRRSV代表株HENAN-XINX同源性为80.8%~81.6%,与经典PRRSV 代表株VR-2332 同源性为80.3%~84.7%,说明鄂西地区流行毒株的ORF5 基因序列正在发生一些新的变化。
自1996 年郭宝清等[9]首次分离到PRRSV CH-1a毒株以来,Lineage8.7分支PRRSV 在我国已流行20 多年。通过遗传进化分析可知,本研究获得的2022YCLPFA、2022YCLPFA1、2022YCYZD 3 份阳性样品ORF5 序列与毒株JXA1、TJ-M在同一分支(Lineage 8.7),属于HP-PRRSV 毒 株;2022YCZDBLZ 与JL580、HENAN-XINX 遗传关系较近,在同一分支(Lineage 1.9),属于NADC30-like PRRSV 毒株。Lineage 1.9 分支PRRSV 自2013 年以来也在我国广泛流行,并不断发生变异和重组[10]。我国先后研制了经典株灭活疫苗、高致病性毒株减毒活疫苗等商品化疫苗,虽然在控制PRRS 大规模暴发方面发挥了一定作用,但PRRSV 易变异,类NADC30 毒株也开始广泛流行,因而期待更为安全有效,特别是针对类NADC30 毒株的疫苗尽早问世。
研究[11]表明,GP5 蛋白上存在的中和抗原位点与免疫保护有关。本研究获得的4 份PRRSV GP5 氨基酸序列与参考毒株比对,在诱骗表位(27~30 aa)发现氨基酸替换29G →29C,在中和表位,HP-PRRSV 毒株出现氨基酸替换39A →39T,类NADC30 毒株出现38A →38T。这些变异有可能会影响机体中和抗体的产生,但需进一步研究。
综上所述,鄂西地区存在HP-PRRSV 和NADC30-like PRRSV 两种毒株的流行,并且其GP5 蛋白氨基酸序列已发生了多个变异,有可能会影响疫苗的免疫保护效果。建议持续加强监测,一定要选择与本场流行毒株匹配的疫苗进行免疫。
猜你喜欢
科学大观园(2022年2期)2022-01-23 11:05:15
湖北畜牧兽医(2017年12期)2017-12-28 16:03:13
法医学杂志(2015年4期)2016-01-06 12:36:36
法医学杂志(2015年4期)2016-01-06 12:36:36
动物医学进展(2015年10期)2015-12-07 05:46:18
湖北教育·综合资讯(2015年8期)2015-11-16 19:55:46
吉林农业·下半月(2014年11期)2014-11-25 12:28:56
卷宗(2014年9期)2014-10-20 07:36:20
特产研究(2014年4期)2014-04-10 12:54:12
郑州大学学报(理学版)(2014年3期)2014-03-01 04:21:08
