
传染性法氏囊病毒VP2蛋白嵌合新城疫病毒样颗粒的构建和鉴定
2022-11-15 06:20:22邵亚男冯嘉轩李金斗郭春红丁佳欣陈铭桦单春晖
中国兽医学报 2022年10期
邵亚男,冯嘉轩,李金斗,郭春红,丁佳欣,陈铭桦,丁 伟,单春晖,张 頔,丁 壮
(吉林大学 动物医学学院 人兽共患病研究教育部重点实验室,吉林 长春 130062)
传染性法氏囊病(infectious bursal disease,IBD)是由传染性法氏囊病毒(infectious bursal disease virus,IBDV)引起的一种急性、高度接触性传染病,主要侵害3~6周龄雏鸡,其属于免疫抑制性疾病[1]。自1962年被首次报道后在全球范围内广泛传播流行,给养禽业造成了严重的经济损失[2]。我国于1979年由周蛟[3]首次发现并分离该病毒,而后在我国各地陆续有强毒株、超强毒株和变异株被分离,目前IBDV几乎散布于我国所有养禽地区。临床上感染IBDV后,会对法氏囊造成不可逆的损伤,其破坏鸡群的体液免疫,造成免疫抑制,可提高其他疾病的易感率与病死率[4],特别是致死率极高的新城疫(Newcastle disease,ND),一旦感染将造成不可估量的损失。大量研究表明VP2蛋白是IBDV重要的保护性蛋白[5-6],能够诱导机体产生中和抗体,是研制IBDV的主要候选抗原。IBDV通过VP2蛋白与受体细胞膜吸附结合,侵入细胞[7],在病毒的感染复制中起到关键作用,因此本研究利用VP2蛋白作为疫苗的主要抗原蛋白。
ND是由新城疫病毒(Newcastle disease virus,NDV)引起的一种急性、热性、高度接触性禽类传染病[8],被我国农业农村部列为一类动物疫病。NDV在感染宿主细胞时,首先由血凝素-神经氨酸酶(hemagglutinin-neuraminidase,HN)蛋白与细胞膜上的唾液酸受体结合,进一步激活F蛋白(fusion protein),由F蛋白参与促进病毒粒子与细胞膜融合过程[9],这表明NDV的感染复制过程与HN、F蛋白密切相关。研究表明,HN、F蛋白作为抗原蛋白构建疫苗能够诱导产生很高的抗体水平[10-11]。20世纪初,PANTUA等[12]在研究中发现,M蛋白(matrix protein)是NDV的驱动力蛋白,单独表达M蛋白即可形成颗粒状结构。因此本研究将IBDV强毒株保护性蛋白VP2替换NDV HN蛋白胞外域部分,与NDV的M、F和HN蛋白共转染至昆虫细胞,组装成含有IBDV候选抗原表位的NDV病毒样颗粒(virus-like particles,VLPs)。因窗口期的存在,现有的IBDV疫苗与NDV疫苗不能同时使用,同时增加了人力物力和时间成本,因此研制一针多防、安全有效的新型疫苗刻不容缓。
自投射电子显微镜出现以来,VLPs也逐渐进入人们的视野[13]。VLPs具有类似于在有限空间内显示密集重复表位特征的天然病毒粒子的形态[14],属于更结构化的重组疫苗,在刺激免疫系统方面具有“强激活剂”的美称。此外,VLPs不具有病毒基因组,缺乏复制能力,是非感染性的重组疫苗,在生物安全性、免疫原性和平台丰富性方面展示了极高的优势。近年来,一些VLPs产品获得了人们的广泛认可,并取得了显著的临床效果和经济价值。目前,市场上的乳头瘤病毒(HPV)[14-16]、乙型肝炎病毒(HBV)[17-19]、戊型肝炎病毒(HEV)[20]、流感病毒(influenza virus)和甲型肝炎病毒(HAV)[21-22]几种重组疫苗都基于高度纯化的VLPs。此外,针对疟疾[23]、诺沃克病毒[24]和基孔肯亚病毒(CHIK)[25]的VLPs疫苗目前正在临床试验中。这些有效的临床应用和极高的市场认可度鼓励了更多的疾病利用VLPs平台开展临床前和临床上相关疫苗的开发和测试。
1 材料与方法
1.1 细胞、病毒及载体Sf9昆虫细胞(CRL-1711)、E.coliDH10Bac、E.coliDH5α感受态细胞、rBV NA-1 M、rBV NA-1 HN、rBV NA-1 F、杆状病毒表达载体pFastBac1均由本实验室保存。
1.2 主要试剂pEASY®-Basic Seamless Cloning and Assembly Kit、X-gal购自北京全式金生物技术有限公司;SalⅠ、KpnⅠ限制性内切酶购自TaKaRa公司;X-treme GENE HP DNA Transfection Reagent购自ROCHE公司;Sf-900TMⅡ SFM无血清无蛋白昆虫细胞培养基购自Gibco公司;胎牛血清购自BI公司;VP2多抗由本实验室保存。
1.3 目的基因的优化及合成根据昆虫细胞密码子偏好性和pFastbac-1载体信息对GenBank:SH99(LM651365.1)VP2核酸序列进行密码子优化,并依次引入蜂素信号肽序列、His标签序列、TEV切割序列,委托生工生物工程(上海)股份有限公司合成,命名为PUC-VP2。
1.4 引物设计根据生物公司合成的PUC-VP2与NA-1(DQ659677.1)HN核酸序列和pFastBac1载体信息分别设计含pFastBac1载体序列和柔性肽的VP2、含有柔性肽和pFastBac1载体信息且去除胞外域的HN引物及M13通用引物,详细引物信息见表1。

表1 详细引物信息
1.5 重组穿梭质粒的构建和鉴定以本实验室保存的pCI-NA-1为模板,通过特异性引物rdHN+pFastBac1-F、rdHN+linker-R扩增出去除胞外域且含有柔性肽和pFastBac1载体信息的片段uniseamless-rdHN-linker,uniseamless-rdHN-linker片段在与pFastBac1同源臂部分引入酶切位点BamHⅠ;利用生物公司合成的PUC-VP2质粒为模板,以rdVP2+linker-F、rdVP2+pFastBac1-R为引物特异性扩增出含pFastBac1载体序列和柔性肽的VP2片段,并将pFastBac1载体信息引入添加到MCS区的KpnⅠ酶切位点处,命名为uniseamless-rdVP2-linker。将pFastBac1空载体信息利用BamHⅠ、KpnⅠ进行双酶切,酶切后的pFastBac1与经过PCR扩增的uniseamless-rdHN-linker、uniseamless-rdVP2-linker利用1%的琼脂糖凝胶鉴定正确后,胶回收其目的片段。将线性化的pFastBac1载体片段、uniseamless-rdHN-linker、uniseamless-rdVP2-linker与无缝克隆盒子中的Assembly Mix混匀,在同源重组酶的作用下进行无缝克隆。作用完成后转化至E.coliDH5α感受态细胞,次日挑取单菌落接种到LB中进行菌液PCR鉴定。利用特异性引物和pFastBac1通用引物鉴定正确后的菌株再提取质粒,双酶切鉴定,送去生物公司测序。将测序正确的重组穿梭质粒命名为pFastBac-cHN+VP2。
1.6 重组杆粒的构建和鉴定将构建好的重组穿梭质粒pFastBac-cHN+VP2转化入E.coliDH10Bac感受态细胞中,通过蓝白斑筛选的方法,筛选转座完成的阳性菌落。挑取大小均一的单一白色菌落加入LB培养,利用特异性引物与通用引物进行菌液PCR鉴定。阳性菌液在3次涂板筛选获取阳性菌落后,送至生物公司测序,测序正确的阳性质粒通过无内毒素提取试剂盒提取杆粒,再利用PCR方法鉴定,鉴定正确得到的杆粒即为rBacmid-cHN+VP2。
1.7 重组杆状病毒的拯救和基因组鉴定将重组杆粒利用转染试剂X-tre006De GENE HP DNA Transfection Reagent转染至Sf9细胞,持续观察。第5天出现细胞病变,收取细胞上清,即为第一代杆状病毒,继续盲传3代,收获细胞上清得到P3代杆状病毒。采用病毒基因组提取试剂盒提取重组杆状病毒基因组,利用特异性引物和通用引物进行PCR鉴定,以确保重组杆状病毒构建正确,将鉴定正确的重组杆状病毒命名为rBv-cHN+VP2。
1.8 IFA检测鉴定重组杆状病毒将细胞爬片放入细胞培养板中,在爬片上培养Sf9细胞至细胞密度约为80%。将rBv-cHN+VP2、rBv-M、rBv-HN、rBv-F以MOI=5共感染Sf9细胞,同时设置未感染组作为阴性对照,在细胞培养箱中培养36 h,使用4%多聚甲醛溶液固定后,再通过Triton通透处理,一抗为NDV M、NDV F、NDV HN以及IBDV VP2的兔源多抗,二抗为FITC标记的猪抗兔IgG抗体,操作时严格避光。用Hoechst室温避光进行染核后处理封片,利用荧光显微镜观察。
1.9 IBD-ND cVLPs的产生与鉴定将Sf9细胞悬浮培养至2×106/mL,以MOI=5接种P4代rBv-cHN+VP2、rBv-M、rBv-HN、rBv-F,27℃、200 r/min环境下培养3 d,收集细胞上清经高速离心机离心处理,继续用超速离心机过20%,30%,60%蔗糖纯化VLPs。将纯化后的VLPs制样通过Western blot鉴定,鉴定正确后再利用透射电镜观察VLPs的形态。
2 结果
2.1 目的基因的扩增以实验室保存的pCI-NA-1为模板,通过特异性引物rdHN+pFastBac1-F、rdHN+linker-R扩增出去除胞外域且含有柔性肽和pFastBac1载体信息的片段uniseamless-rdHN-linker(图1),利用生物公司合成的PUC-VP2质粒为模板,以rdVP2+linker-F、rdVP2+pFastBac1-R为引物特异性扩增出含pFastBac1载体序列和柔性肽的VP2片段(图2),1%琼脂糖凝胶电泳显示条带大小与预期相符。

M.DL2000 DNA Marker;1.特异性引物PCR扩增cVP2基因产物

M.DL2000 DNA Marker;1.特异性引物PCR扩增cHN基因产物
2.2 重组穿梭质粒的鉴定线性化的pFastBac1载体片段与uniseamless-rdHN-linker、uniseamless-rdVP2-linker通过无缝克隆方法连接形成重组穿梭质粒pFastBac-cHN+VP2,重组穿梭质粒pFastBac-cHN+VP2利用pFastBac1通用引物鉴定以及BamHⅠ、KpnⅠ双酶切鉴定,由pFastBac1通用引物鉴定结果(图3)以及BamHⅠ、KpnⅠ双酶切鉴定结果(图4)可知目的条带与理论值相符。
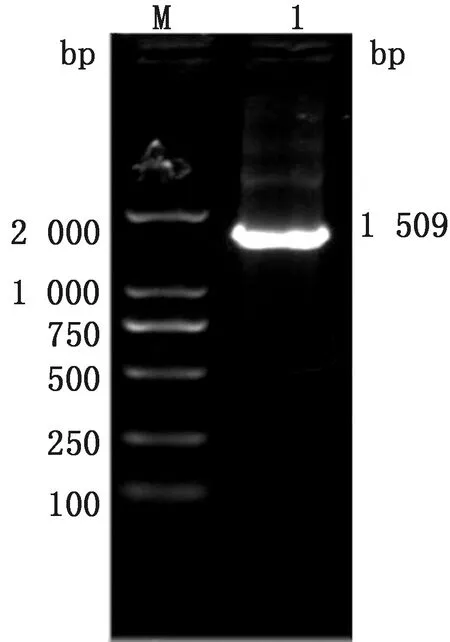
M.DL2000 DNA Marker;1.pFastBac通用引物鉴定
2.3 重组杆粒的鉴定将构建好的重组穿梭质粒转化到E.coliDH10Bac菌中完成转座过程,通过3次划线涂板实现蓝白斑筛选后,挑取完全的白斑接种到液体LB中,用M13通用引物和特异性引物进行菌液PCR。菌液PCR鉴定结果显示目的条带与理论值相符,重组杆粒rBacmid-cHN+VP2构建正确(图5)。

M.DL5000 DNA Marker;1.cHN+VP2-pFastBac酶切鉴定

M.DL5000 DNA Marker;1.rBacmid-cHN+VP2 M13鉴定;2.rBacmid-cHN+VP2特异性引物鉴定
2.4 重组杆状病毒的鉴定
2.4.1Sf9细胞病变 状态良好的Sf9细胞在感染拯救的杆状病毒后盲传,持续用光学显微镜观察,结果出现肿大变圆甚至破裂的状态(图6),且生长速度较正常细胞缓慢。

A.正常Sf9细胞;B.rBv cHN+VP2感染后Sf9细胞
2.4.2基因组鉴定 将重组杆状病毒rBv-cHN+VP2感染Sf9细胞后盲传3代,收获Sf9细胞培养上清提取重组杆状病毒基因组,将获得的重组杆状病毒基因组通过PCR方法鉴定,鉴定引物包括特异性引物与M13通用引物,1%琼脂糖凝胶电泳结果显示条带大小与预期值相符(图7)。

M.DL5000 DNA Marker;1.rBv-cHN+VP2基因组M13鉴定;2.rBv-cHN+VP2基因组特异性引物鉴定
2.4.3IFA鉴定 将处理好的杆状病毒细胞爬片使用荧光倒置显微镜观察,可见各试验组均出现绿色荧光信号,而阴性对照组无荧光信号出现(图8),显示各组分蛋白表达状况正确。
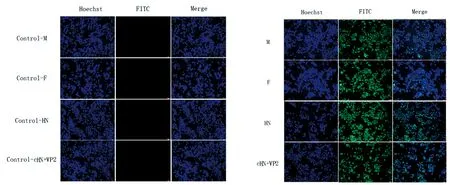
图8 IFA鉴定蛋白表达
2.5 IBD-ND cVLPs的产生与鉴定
2.5.1Western blot鉴定 纯化后的IBD-ND cVLPs制样后,分别用多克隆抗体NDV M、NDV F、NDV HN和IBDV VP2为一抗,FITC标记的猪抗兔IgG抗体为二抗,通过Western blot鉴定,结果显示各组分蛋白表达良好,cVLPs构建正确(图9)。

图9 IBD-ND cVLPs的Western blot鉴定
2.5.2电镜观察 取适量样品置于铜网上处理观察,发现纯化后的IBD-ND cVLPs具有囊膜和纤突结构(图10)。

图10 IBD-ND cVLPs的透射电镜观察
3 讨论
自IBD流行肆虐以来,给养殖业造成了无可估量的经济损失,因其给鸡群带来的病理变化和致病特点,它有“禽肾病”、“鸡艾滋病”这些别名[26]。近20年来,IBD防控态势逐年升级,变异株和超强毒株出现频率增加[27],且具有多样的血清亚型和抗原变异,现已成为危害养禽业最棘手最严重的病毒性传染病之一,引起了我国兽医领域的广泛重视。一旦感染IBDV,对禽类的免疫系统造成的是不可逆转的损伤,进一步提高继发感染NDV、大肠杆菌和支原体的可能性,致使病死率提高,造成严重的经济损失。NDV、IBD这2种传染病预防都使用弱毒疫苗防控,存在接种窗口期,大大增加了患病的风险。我国预防和防制IBD主要使用弱毒疫苗B87株,与目前我国出现频率越来越高的超强毒株和变异株同源性降低,而且存在着变异和散毒的风险。这与我国NDV防制出现的问题相仿,LaSota弱毒疫苗株与目前国内流行株NA-1株同源性仅82%左右。这些问题都限制着我国动物传染病的防制进程,因此本研究中NDV的主要保护性抗原和基质蛋白来自国内流行株NA-1株,IBD的主要保护性抗原部分来自于当前流行的IBDV强毒株SH99株,这为我国这两类禽病防控提供高度同源的疫苗选择。综合以上原因研制无核酸、绿色安全有效、无需等待免疫窗口期的二联疫苗相当有必要。
IBDV的VP2蛋白作为病毒外衣壳的唯一成分,位于病毒粒子的最外层[28]。有学者研究发现,在IBDV感染早期,VP2蛋白可以通过抑制AKT-MTOR通路的活性,诱导自噬的发生,进而抑制IBDV的复制[29]。VP2蛋白具有的20个独立的VP2亚单位,可以作为中和抗体结合的靶标,同时,成熟的VP2蛋白能够诱导产生保护宿主的保护性抗原[30]。综合以上原因,针对VP2蛋白的疫苗广泛应用于预防IBD。因此本试验利用杆状病毒表达系统构建了含有IBDV候选抗原表位的VP2蛋白,与NDV-HN、NDV-F、NDV-M组装成cVLPs,形成含有IBDV候选抗原表位的IBD-ND二联病毒样颗粒。
病毒样颗粒(VLPs)被认为是疫苗开发的有用工具,因为它们能诱导免疫反应并且是安全的。VLPs保留了免疫原性蛋白的天然抗原构象和原始病毒颗粒的重复结构,可以被抗原呈递细胞(APC),尤其是树突状细胞(DC)有效识别,然后被MHCⅡ类分子加工和呈递以刺激CD4+T辅助细胞[31],也能在经过DC处理后通过交叉呈递机制由MHCⅠ类分子呈递给细胞毒性CD8+T细胞,有效激活体液免疫、黏膜免疫、细胞免疫。VLPs技术的灵活性允许VLPs成为在结构内或颗粒表面掺入和显示异源抗原或特定表位的平台[32-33]。NDV VLPs因其极高的释放率是极好的载体选择。因此,嵌合IBD-ND VLPs是一个很有前景的疫苗平台。此外,某些VLPs还被用作潜在的药物载体和生物医学成像剂。它们缺乏病毒遗传物质,由具有固有自组装能力的结构蛋白形成,可作为比全病毒疫苗更安全的展示外来抗原的疫苗平台[34]。除GPI锚定策略外,NDV VLPs的F蛋白和HN蛋白跨膜蛋白的特点允许通过胞外域替换的方式构建嵌合VLPs,因此本研究将IBDV强毒株保护性蛋白VP2替换NDV HN蛋白胞外域部分,与NDV-HN、NDV-F、NDV-M组装成cVLPs,利用Western blot和透射电镜鉴定cVLPs各组分组装正确,充实了NDV VLPs作为载体平台的使用方向,为ND和IBD的防制提供了疫苗储备。
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06 03:52:34
当代水产(2021年7期)2021-11-04 08:17:54
食品科学(2018年10期)2018-05-23 01:27:28
中国兽医杂志(2016年5期)2016-06-27 01:44:58
广东海洋大学学报(2015年4期)2016-01-13 08:39:33
西南医科大学学报(2015年1期)2015-08-22 13:01:46
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
中国当代医药(2015年9期)2015-03-01 02:01:59
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
