
Metabolomic and proteomic responses of Phaeodactylum tricornutum to hypoxia*
2022-10-08 02:22PeipeiZHAOQinghuaWUXuekuiXIAShiyiGUOSizhongSHENYujueWANGAiyouHUANG
Peipei ZHAO , Qinghua WU , Xuekui XIA , Shiyi GUO , Sizhong SHEN ,Yujue WANG , , Aiyou HUANG , ,**
1 Key Biosensor Laboratory of Shandong Province, Biology Institute, Qilu University of Technology (Shandong Academy of Sciences), Jinan 250103, China
2 State Key Laboratory of Marine Resource Utilization in the South China Sea, Hainan University, Haikou 570228, China
3 Laboratory of Development and Utilization of Marine Microbial Resource, Hainan University, Haikou 570228, China
4 Key Laboratory of Tropical Hydrobiology and Biotechnology of Hainan Province, Haikou 570228, China
5 College of Marine Sciences, Hainan University, Haikou 570228, China
6 Department of Chemistry, University of Washington, Seattle WA 98195, USA
Abstract Diatoms are important contributors to global net primary productivity, and play a crucial role in the biogeochemical cycles of carbon, phosphorus, nitrogen, iron, and silicon. Currently in some regions in the ocean, there’s a trend that carbon content is high while oxygen concentration is low, and the underlying mechanisms of diatoms’ response to low oxygen environments are worth investigating. Phaeodactylum tricornutum is a model diatom whose genome has been sequenced; it provides a universal molecular toolbox and a stable transgenic expression system. Therefore, the study of the responses of P. tricornutum to low oxygen has not only fundamental research signif icance but also important ecological signif icance. In this study,growth rates were determined and proteomic analysis and metabolomic analysis were performed to examine P. tricornutum responses under diff erent oxygen concentrations (2% oxygen concentration for hypoxic condition and 21% oxygen concentration for the normal condition (sterilized air)). Results show that the hypoxic environment inhibited the growth of P. tricornutum. In the hypoxic conditions, P. tricornutum could reset its metabolism pathways, including enhancement in lipid utilization, replenishment of tricarboxylic acid (TCA) cycle through the glyoxylic acid cycle, and down-regulation of photorespiration to reduce energy waste. Additionally, the stress resistance mechanism was activated to facilitate the adaptation to low oxygen conditions. This study helps to reveal the diff erent metabolic changes to hypoxia of diatom from that of higher plants, which might be ascribed to their diff erent habitats and needs further exploration in the future.
Keyword: Phaeodactylum tricornutum; hypoxia; metabolomics; proteomics* Supported by the National Natural Science Foundation of China (Nos.41876158, 31770024), the Natural Science Foundation of Hainan Province(No. 420QN219), the Biology and Biochemistry ESI Cultivation Discipline Open Project of Qilu University of Technology (No. ESIBBC202004), the Innovation and Development Joint Fund of Natural Science Foundation from Shandong Province (No. ZR2021LSW022), the Young Taishan Scholarship to Xuekui XIA (No. tsqn202103100), and the Start-up Fund Project of Hainan University (No. KYQD(ZR)20060)** Corresponding author: huangaiyou08@163.com# Peipei ZHAO and Qinghua WU contributed equally to this work and should be regarded as co-f irst authors.
1 INTRODUCTION
Plants and algae can respond to low oxygen by remodeling their metabolism pathways. The transition from mitochondrial respiration to fermentation is representative of anaerobic metabolism in most organisms (Geigenberger, 2003; Banti et al., 2013).Rapid activation of lactate dehydrogenase (LDH)catalyzing the reduction of pyruvate to lactate occurs in almost all plant species under hypoxic conditions(Rivoal et al., 1991; Sweetlove et al., 2000).Chlamydomonasproduces hydrogen (H2) in the absence of oxygen. It is speculated that H2may have evolved as a component of photoprotection, but the precise physiological functions of hydrogenase and H2release inChlamydomonasremains unclear(Grossman et al., 2011).
Additionally, hypoxia may lead to decreased rate of photorespiration, which can aff ect cellular carbon and nitrogen metabolism pathways. For example, in cyanobacteria, the enhanced photorespiration due to low carbon availability leads to an accumulation of 2-phosphoglycolate, a sign of carbon def iciency(Jiang et al., 2018). 2-phosphoglycolate also promotes the expression of carbon assimilation-related genes and thus regulates carbon and nitrogen balance(Zhang et al., 2018). Photorespiration is associated with intracellular (near ribulose-1,5-bisphosphate carboxylase/oxygenase, RuBisCO) carbon-oxygen levels (Jiang et al., 2018). Thereby, if hypoxia can lead to a decrease in photorespiration and 2-phosphoglycolate levels, and in turn, reduce carbon assimilation, remains unknown. Hypoxia may also aff ect oxidative phosphorylation (Solaini et al., 2010),and thus alter the cellular energy state and induce changes in cellular carbon metabolism (glycolysis,TCA cycle, lactic acid metabolism, and glyoxylic acid cycle) to maintain an intracellular balance between matter and energy.
Diatoms are important unicellular photoautotrophic eukaryotes that play a crucial role in ecology; they contribute to one-f ifth of the total primary productivity on the Earth (Falkowski et al., 1998; Field et al., 1998;Kroth, 2007). The success of diatoms in marine environments is attributed to their rapid response to nitrogen f luctuations. In addition, in some regions in modern oceans, high carbon and low oxygen trends are prevalent (Khangaonkar et al., 2021).PhaeodactylumtricornutumBohlin is the f irst diatom whose complete genome is sequenced, which provides a universal molecular toolbox and a stable transgenic expression system (Bowler et al., 2008). Thus, it is a model organism for studying physiology, evolution,and biochemistry. In addition,P.tricornutumis widely distributed across coastal and inland waters(including but not limited to the Great Salt Lake),where the environment is unstable and temperature,light quality, salinity, and dissolved oxygen levels f luctuate rapidly (Rushforth et al., 1988; Yang and Zhang, 2020). Therefore, it is of great ecological and physiological signif icance to study the underlying response mechanism of diatoms in a low oxygen environment usingP.tricornutummodel. In this study, we compared the growth rates and pigment contents ofP.tricornutumunder diff erent oxygen concentrations. Proteomic and metabolomic analyses were conducted. The results help to reveal the diff erent metabolic changes to hypoxia of diatom from that of higher plants, which might be ascribed to their diff erent habitats and needs further exploration in the future.
2 MATERIAL AND METHOD
2.1 Strain, culture conditions, and growth measurements
Phaeodactylumtricornutumwere cultured axenically in f/2 medium supplemented with steamsterilized NaHCO3-free artif icial seawater, f/2 vitamins(0.5-μg/mL Biotin, 100-μg/mL Vitamin B1, 0.5-μg/mL Vitamin B12, f ilter sterilized), and inorganic nutrients(Guillard, 1975; Harrison et al., 1980). Cultures were incubated in cool-white-f luorescent light at 100 μmol/(m2·s) in 12 h∶12 h light∶dark cycle at 20 °C for approximately seven days. In low oxygen condition(LO), cultures were incubated in 2-L f lasks containing 1.5-L medium, continuously aerated with a sterilized gas mixture of 2% O2, 0.04% CO2, and 97.96% N2,bubbled at a constant f low rate of 500 mL/min. In normal conditions (NC), sterilized air was used instead of the hypoxia gas mixture. Each treatment was conducted in triplicates. Cell growth was measured at 730 nm using a ultraviolet-visible (UV-VIS)spectrophotometer (UV-1800, Shimadzu, Japan)(Zhao et al., 2018). Cells were harvested after eight days of cultivation by centrifugation for 5 min at 4 000×g. Cell pellets were frozen instantly in liquid nitrogen and stored at -80 °C until protein extraction and further use.
2.2 Metabolomics analysis
Samples were freeze-dried and suspended in 1 mL of 50% v/v methanol-water buff er (vortexed for 1 min). Mixtures were centrifuged at 12 000×gfor 5 min; the supernatant was lyophilized and redissolved in 0.45-mL water. Subsequently, a 50-μL 2,2-dimethyl-2-silapentane-5-sulfonate (DSS)standard solution (Anachro, Canada) was added.Samples were mixed well before transferring into 5-mm nuclear magnetic resonance (NMR) tube(Norell, USA). Spectra were collected on the Bruker AV III 600-MHz spectrometer. The f irst increment of a 2D-1H,1H-NOESY pulse sequence was utilized for the acquisition of1H-NMR data and for suppressing the solvent signal. The mixing time was set to 100 ms along with a 990-ms pre-saturation time (~80-Hz gammaB1). A total of 128 scans for 15 min; spectra were collected at 25 °C.
2.3 P. tricornutum pigment analysis
Pigment extraction and analysis fromP.tricornutumwere performed according to the previously described methods of Thayer and Björkman (1990), and Enriquez et al. (2010) with slight modif ications. Brief ly, the collected cells were freeze-dried, suspended in acetone/methanol (1/1,v/v) on ice for 1 h, and then centrifuged at 6 000×gfor 5 min. The extraction was repeated thrice and the supernatant was pooled. After f iltration, the supernatant was injected into the Agilent 1260 high performance liquid chromatography (HPLC)equipped with a reversed-phase column (YMC-Pack ODS-A, 10×250 mm). Analysis of the mobile phase,consisting of water, methanol, and acetonitrile, was programmed as follows: 0-15 min, linear gradient from I (15% water, 30% methanol, and 55%acetonitrile) to II (0% water, 15% methanol, and 85%acetonitrile); 15-30 min, linear gradient from II to III(100% methanol); and 30-40 min, III. The column temperature was set at 50 ℃ and the f low rate was 0.75 mL/min. Fucoxanthin was detected at 450 nm.
2.4 Preparation of P. tricornutum water-soluble proteins
Cell pellets were ground in liquid nitrogen using a mortar and pestle. Pulverized cells were suspended in 10-mL phenol extraction buff er (Solarbio, T0250,containing 10-mmol/L dithiothreitol (DTT) and 1%protease inhibitor cocktail). Mixtures were subject to ultrasonic lysis. One volume of tris-phenol buff er was added to the mixtures. The cell lysate was centrifuged at 5 500×gfor 10 min at 4 °C and the phenol supernatant was collected. Five volumes of 0.1 mol/L cold ammonium acetate-methanol buff er were added to the supernatant and precipitated overnight at-20 °C. The precipitate was washed using cold methanol and acetone and dissolved in 8-mol/L urea.Protein concentration was determined by the Bradford assay (Bradford, 1976).
2.5 Label-free quantitative proteomics
Trichloroacetic acid at a f inal concentration of 20%was added to the samples, eddy mixed and precipitated at 4 ℃ for 2 h. The supernatant was discarded after centrifugation at 4 500×gfor 5 min at 4 ℃. The supernatant was washed twice and precipitated with pre-chilled acetone. After the precipitate was dried,tetraethylammonium bromide (TEAB) at a f inal concentration of 200 mmol/L was added, the precipitate was dispersed by ultrasound, and trypsinized at a ratio of 1∶50 (protease∶protein, m/m)overnight (enzymatic hydrolysis). DTT was added to a f inal concentration of 5 mmol/L and reduced at 56 ℃ for 30 min. Iodoacetamide (IAA) was added to make up to the f inal concentration to 11 mmol/L and incubated at room temperature for 15 min in dark(Wiśniewski et al., 2009).
The peptides were dissolved in mobile phase A(0.1% formic acid, 2% acetonitrile) and separated using a NanoElute ultra-high performance liquid phase system. Liquid phase gradient settings were as follows: 0-70 min, 6%-24% mobile phase B (0.1%formic acid in acetonitrile); 70-84 min, 24%-35% B;84-87 min, 35%-80% B; 87-90 min, 80% B, and the f low rate was maintained at 450 nL/min. After separation on the ultra-high performance liquid phase platform, the peptide was injected into the capillary ion source for ionization and subsequently analyzed using timsTOF Pro instrument. The voltage of the ion source was set at 1.7 kV. High resolution time-off light (TOF) was used to detect and analyze the parent ion of the peptide and its secondary fragments.Maxquant software (v1.6.15.0) was used to search the secondary mass spectrum data. Search parameter settings and the process were as follows: The database isPhaeodactylum_tricornutum_strain_CCAP_1055_556484_PR_20200709.fasta (10 465 sequences). Database searches were performed with trypsin digestion specif icity. The Maxquant database search results show the label-free quantif ication(LFQ) intensity of each protein in diff erent samples(the original intensity value of the protein is corrected between samples). After centralizing the LFQ intensity (I) of the protein in diff erent samples, the relative quantitative value (R) of the protein in diff erent samples was calculated by the formula ofRij=Iij/mean(Ij) (irepresents the sample andjrepresents the protein). For multiple replicate sample experiments, calculate the average value of the relative quantitative value of each protein in multiple replicates, and then calculate the ratio of the average of the two samples, and this ratio was used as the f inal diff erential expression of the protein between the two samples ratio. Studentt-test was conducted on the triplicates in each group. Signif icantly diff erentially expressed proteins were def ined by the criteria of a fold change (FC) ≥1.5 or ≤0.67 (P<0.05).Functional annotation of the proteins was performed using non-redundant protein (NR), Swiss Protein(SwissProt), Gene Ontology (GO), and Kyoto Encyclopedia of Genes and Genomes (KEGG)databases.
2.6 Protein validation by parallel reaction monitoring (PRM)
To verify the protein expression levels in label-free quantitative proteomics, the expression levels of six selected proteins were quantif ied by PRM analyses.The method for protein extraction and tryptic digestion were the same as described above for the label-free experiment. The tryptic peptides were dissolved in 0.1% formic acid and directly loaded onto a homemade reversed-phase analytical column using the EASY-nLC 1000 ultra performance liquid chromatography (UPLC) system. Mobile phase A was 0.1% formic acid and 2% acetonitrile in water,and mobile phase C was 0.1% formic acid in 90%acetonitrile. Liquid phase gradient settings were as follows: 0-16 min, 7%-25% C; 16-22 min, 25%-35% C; 22-26 min, 35%-80% C. The f low rate was set at 500 nL/min. The peptides were examined using the Q Exactive Plus tandem mass spectrometry(Thermo Fisher Scientif ic) platform where the nanoelectrospray ionization (NSI) source was coupled online with the UPLC. The electrospray voltage was set to 2.1 kV. The primarym/zscan range was between 430 to 1 200, and the scanning resolution was 70 000.The scanning resolution of secondary mass spectrometry Orbitrap was 17 500. Data independent scanning (DIA) program was used and the fragmentation energy of higher-energy collisional dissociation was 28. The automatic gain control(AGC) of primary mass spectrometry was 3E6, and the maximum injection time was 50 ms. The AGC of secondary mass spectrometry was 1E5, the maximum injection time was 200 ms, and the isolation window was 1.4m/z.
2.7 Statistical analysis
There were three replicates for each condition. The studentt-test was used to determine signif icant diff erences. All data were represented as the mean of three independent experiments. The mean value was used to calculate the ratio (fold changes, FC). For metabolomics analysis, metabolites withP-value<0.05 andP-value <0.01 were marked with an asterisk and two asterisks, respectively. For proteomics analysis, theP-value of the diff erentially expressed proteins shown in the results were all less than 0.01 or 0.05, as indicated.
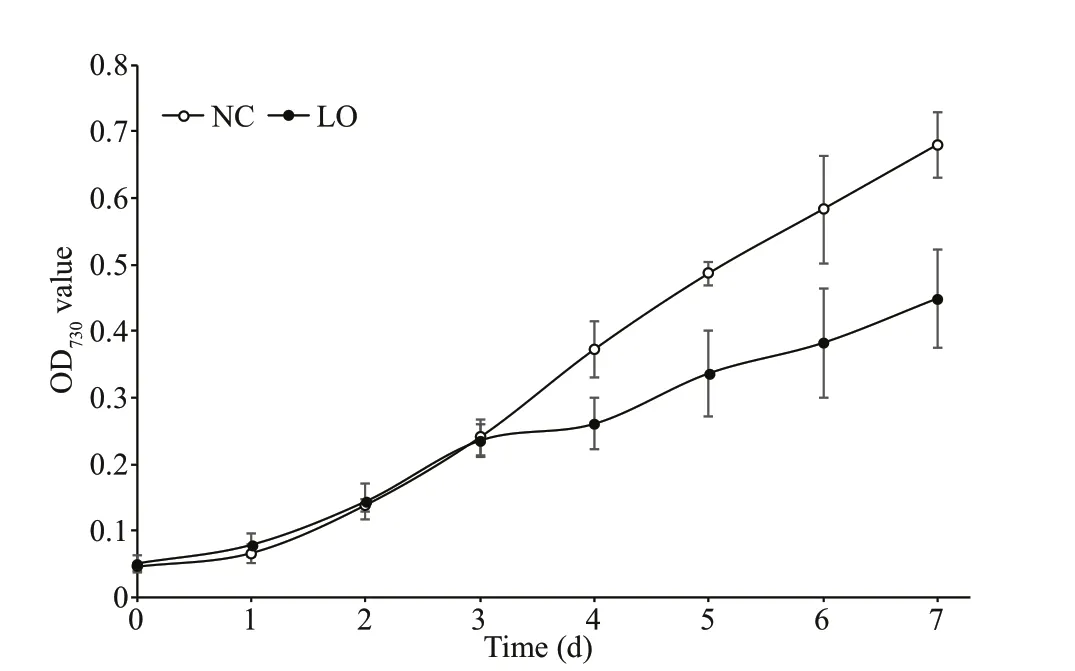
Fig.1 O diDff e73 r0 evnat lcuuel tfuorre tchoen gdri toiwo n th s of P. tricornutum under
3 RESULT
3.1 Growth of P. tricornutum
We compared the growth rates ofP.tricornutumunder hypoxic and normal conditions. The growth rate ofP.tricornutumunder hypoxic condition was signif icantly lower as compared to that under normal conditions (Fig.1), which indicated that hypoxia condition could inhibit the growth ofP.tricornutum.
3.2 Metabolomic analysis
Metabolite prof iling showed that the levels of amino acids such as serine, leucine, phenylalanine,and tyrosine, whose biosynthesis was the derivative of glycolysis were signif icantly reduced in LO culture(P.tricornutumcultured under hypoxic condition)(Table 1). Level of threonine, derivative from TCA cycle, was also reduced. However, levels of other amino acids derivative from the TCA cycle, such as glutamine, increased in LO culture. Ornithine level was also signif icantly increased in LO culture.Intermediates of the TCA cycle, including fumarate and succinate, were down-regulated in LO culture.For amines and ammonium compounds, the levels of dimethylamine and sn-glycero-3-phosphocholine were signif icantly altered between the two conditions(LO and NC).
3.3 P. tricornutum pigment analysis
The fucoxanthin content ofP.tricornutumincreased signif icantly (P<0.01) under hypoxic condition, which was 30.85±0.84 mg/g dry weight(DW) and 25.46±1.61 mg/g DW under hypoxic condition and normal conditions, respectively (Fig.2).
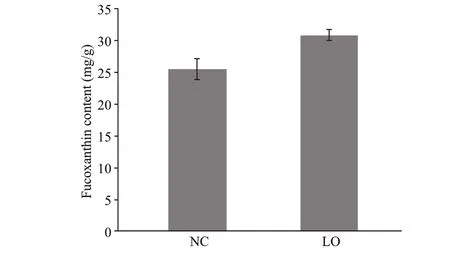
Fig.2 Fucoxanthin content of P. tricornutum under diff erent culture conditions
3.4 Protein expression and identif ication

Table 1 Ratio of abundances of key cellular metabolites under diff erent culture conditions determined by NMR
The proteomic analysis yielded 271 diff erential expression proteins with 129 was up-regulated and 142 was down-regulated (LO/NC:P.tricornutumcultured under hypoxia condition vs.P.tricornutumcultured under normal condition) (data are available via ProteomeXchange with project ID: PXD028642).The potential cellular functions of the diff erentially expressed proteins identif ied were searched for in UniProt (http://www.uniprot.org/) and classif ied into several categories (Figs.3-4). More than half of the proteins had no notation in the database. Annotated proteins were classed into categories and described in Fig.3. Here annotated proteins were proteins with annotation in UniProt (http://www.uniprot.org/).Predicted proteins were proteins which had no notation and only were named as ‘Predicted protein’or ‘Hypothetical protein’ in the database.
3.4.1 Central carbon metabolism
A fructose-bisphosphate aldolase (UniProt accession number, B7GE67) related to the glycolysis pathway was down-regulated in LO culture (Fig.4a).Aconitate hydratase 2/2-methylisocitrate dehydratase(B7FUR4), which catalyzes the interconversion between citric acid and isocitrate in the TCA cycle,was signif icantly increased in LO culture. Propionyl-CoA carboxylase (B7GCL6) and methylmalonyl-CoA mutase (B7FX73), responsible for the f irst and third step reactions of propionyl-CoA to succinyl-CoA conversions, were up-regulated by 2.42 fold and 1.97 fold respectively in LO culture. Propionyl-CoA is an end-product of β-oxidized odd chain fatty acids and it enters the TCA cycle after its conversion to succinyl-CoA. Serine hydroxymethyltransferase(B5Y594), an enzyme that participates in photorespiration, was decreased in LO culture.
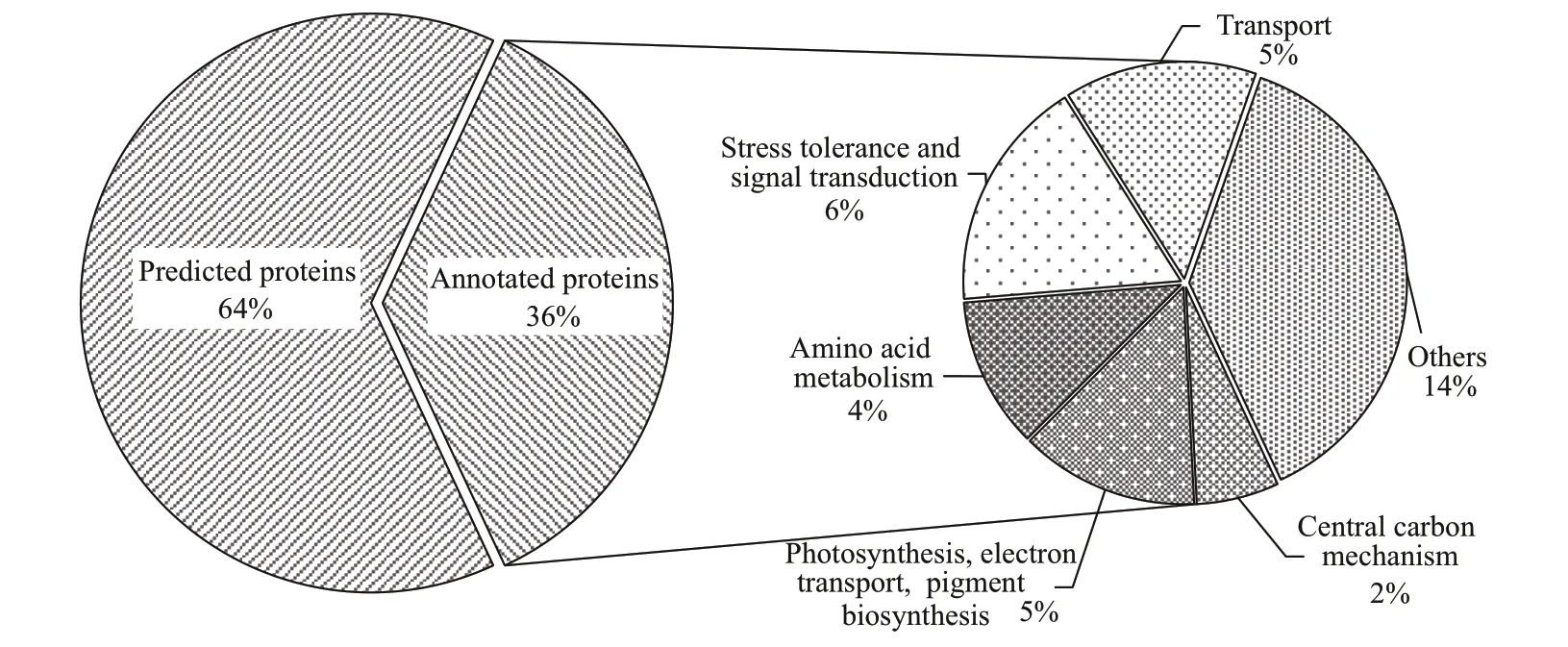
Fig.3 Functional categories of diff erentially expressed proteins
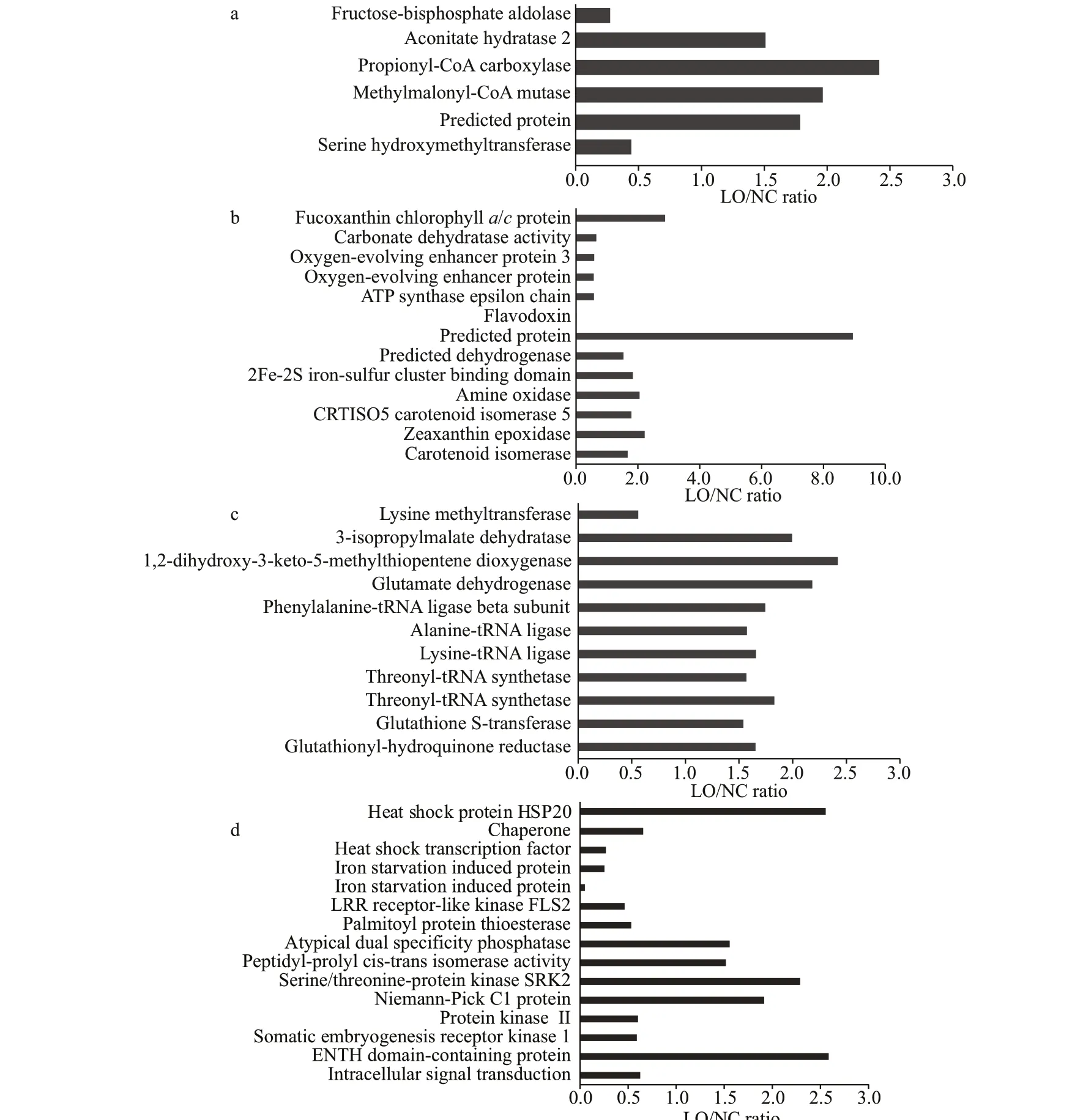
Fig.4 Diff erentially expressed proteins in LO/NC group
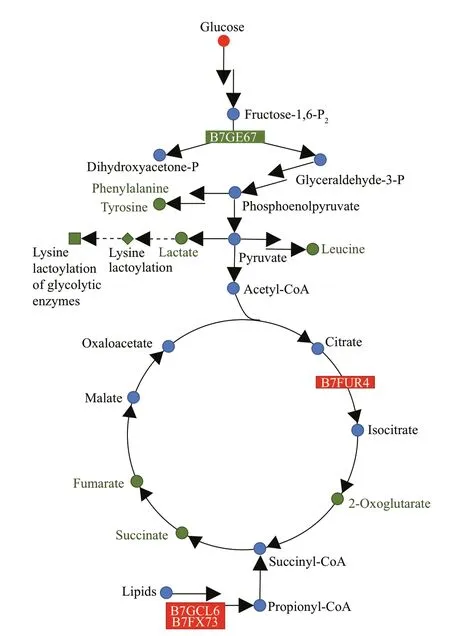
Fig.5 Central carbon metabolism regulation network of P. tricornutum under hypoxic condition
3.4.2 Photosynthesis, electron transport, and biosynthesis of photosynthetic pigments
Fucoxanthin chlorophylla/cprotein (B7G4U8),the antenna protein in diatoms, was up-regulated by 2.88-fold in LO culture (Fig.4b). Two oxygenevolving enhancer proteins (B7FZ94, B7FZ96) were down-regulated in LO culture. An amine oxidase(B7GBV4) and a zeaxanthin epoxidase (B7FUR7)were up-regulated by 2.05-fold and 2.22-fold,respectively, in LO culture. These two up-regulated proteins indicated that the production of photosynthetic pigments could be enhanced in LO culture.
3.4.3 Amino acid metabolism
A lysine methyltransferase (B5Y4M4) was downregulated, while a 3-isopropylmalate dehydratase(B7FPZ8), a 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase (B7FQT4), and a glutamate dehydrogenase(B7G3X3) were up-regulated in LO culture (Fig.4c). A phenylalanine-tRNA ligase beta subunit (A0T0H4), an alanine-tRNA ligase (B7G7J0), and two threonyl-tRNA synthetases (B5Y5F7, B7S423) were also up-regulated in LO culture.
3.4.4 Stress tolerance and signal transduction
A heat shock protein HSP20 (B7G195) was upregulated in LO culture (Fig.4d), while a heat shock transcription factor (B7FWU3) was down-regulated in LO culture. Two iron starvation induced proteins(B7FYL2, B7G4H8) were down-regulated in LO culture. The abundance of other proteins involved in stress tolerance and signal transduction was also altered, which implied that stress response mechanisms ofP.tricornutumwere turned on under hypoxic conditions. The target protein levels detected by PRM were consistent with those in label-free quantitative proteomics, which indicated that the label-free quantitative proteomic results were highly reliable and reproducible (Supplementary Fig.S1).
4 DISCUSSION
4.1 P. tricornutum resets carbon metabolism under hypoxic condition
The response ofP.tricornutumunder LO condition was diff erent from that of higher plants. In almost all plant species under hypoxic conditions, the rapid activation of LDH, which catalyzes the reduction of pyruvate to lactate, was observed (Banti et al., 2013),whereas a down-regulation of the glycolytic enzyme fructose-bisphosphate aldolase (B7GE67) and the glycolytic by-products lactate was observed inP.tricornutumunder LO condition (Fig.5). We proposed that this diff erence might be due to their diff erent structures. The structures of higher plants are more complex, which divide into photosynthetic and non-photosynthetic tissues. The response of nonphotosynthetic tissues to hypoxia is diff erent to that of photosynthetic tissues, with non-photosynthetic tissues are more susceptible to hypoxia. For gray poplar, glycolytic f lux and ethanolic fermentation were stimulated in roots but not in leaves (Kreuzwieser et al., 2009). It is probable that in photosynthetic tissues, O2generation from photosynthesis is enough for the demand of cells. AsP.tricornutumis single cell photosynthetic organism, its response to LO condition might be similar to photosynthetic tissues of higher plants. That is probably why lactate was not increase but decrease inP.tricornutum. Besides,lactate play an important role in regulation of glycolytic enzymes. A preliminary experiment on lactoylation proteomics shows that the lysine lactoylation of the whole proteome decreases inP.tricornutumcultured under hypoxic conditions(unpublished data), which was coincides with the decreased lactate levels. The lysine lactoylation is enriched in glycolytic enzymes and results in decreased glycolytic output (Gaff ney et al., 2020).This is a feedback mechanism similar to that through 3-phosphoglyceryl-Lys, leading to a decrease in the levels of glycolytic metabolites (Moellering and Cravatt, 2013). The down-regulation of glycolytic enzyme and by-products might weaken the feedback mechanism inP.tricornutum.
An up-regulation of aconitate hydratase 2/2-methylisocitrate dehydratase (B7FUR4), which catalyzes citric acid to isocitrate in the TCA cycle,was observed under LO condition. Yet this did not lead to the increase of downstream products, isocitrate and 2-oxoglutarate. Meanwhile, the intermediate products of the TCA cycle, including succinate,2-oxoglutarate, and fumarate, showed a downward trend (Table 1; Fig.5). Amino acids derived from glycolysis and TCA cycle also showed a downward trend (Table 1; Fig.5), including leucine derived from pyruvate, phenylalanine and tyrosine from phosphoenolpyruvate and erythrose-4-phosphate, and so on. We speculated that this might be a result of the down-regulation of glycolysis products as well as TCA cycle products in LO condition. Besides, a rapid consumption of TCA metabolic intermediates might also result in their depletion. For example, the glyoxylate cycle, which requires plenty of isocitrate,is particularly important in plant seeds (Eastmond et al., 2000). The stored lipids in the seeds generate sugars through acetyl-CoA, which timely supply the energy and carbon skeleton and promote germination and growth. The glyoxylate cycle improves the ability of the organism to use acetyl-CoA. With only a small amount of oxaloacetic acid as a primer, the glyoxylate cycle can continually to produce succinate to replenish four carbon units for the TCA cycle. In this study, we observed the up-regulated propionyl-CoA carboxylase(B7GCL6) and methylmalonyl-CoA mutase(B7FX73). This might promote the conversion of propionyl-CoA to succinyl-CoA and enhanced the use of lipids and replenishment of the TCA cycle. Thus,P.tricornutumcould reset its carbon mechanism to cope with hypoxic stress.
4.2 P. tricornutum enhances biosynthesis of photosynthetic pigment under hypoxic condition
Algae accumulate carotenoids under stress conditions. In this study, we observed an increase in fucoxanthin content inP.tricornutumunder LO condition. Accordingly, the key enzyme of photosynthesis, fucoxanthin chlorophylla/cprotein(B7G4U8), was upregulated. Meanwhile, an amine oxidase (B7GBV4) and a zeaxanthin epoxidase(B7FUR7), which participate in carotenoid biosynthesis, were up-regulated. Amine oxidase catalyzes the conversion of phytoene to phytof luene,and phytof luene to ζ-carotene. ζ-carotene can be converted to zeaxanthin in a six-step reaction.Zeaxanthin epoxidase catalyzes the conversion of zeaxanthin to antheraxanthin and antheraxanthin to violaxanthin. Violaxanthin can be converted to neoxanthin and subsequently fucoxanthin (Dambek et al., 2012). Fucoxanthin in fucoxanthin-chlorophyllacomplexes, resembles the light-harvesting complexes, and functions as an antenna and transfers excitation energy to chlorophyll (Owens and Wold,1986; Papagiannakis et al., 2005). Pigment analysis showed that fucoxanthin content inP.tricornutumincreased in LO culture, consistent with proteomic results. Thus, this response may provide continued energy supply forP.tricornutumunder hypoxic condition. This might be another response ofP.tricornutumto LO condition.
4.3 P. tricornutum initiates other stress resistance mechanisms under hypoxic condition
Up-regulated heat shock proteins might be also one of LO responses ofP.tricornutum. Whether oxidative stress exists under hypoxic condition remains controversial. It seems counterintuitive since oxygen is needed for the production of reactive oxygen species (ROS). However, activation of ROS-regulated genes or ROS-associated is reported in diff erent plants under hypoxic conditions (Banti et al., 2013).Heat Shock Proteins (HSPs) are the most common among the ROS-related proteins, which are also induced by anoxia (Mustroph et al., 2010; Pucciariello et al., 2012). Some ROS-related transcription factors(TFs) such as HsfA2 and ZAT12, associated with ROS production are also regulated by hypoxia(Pucciariello et al., 2012). Our results show an upregulation in levels of heat shock protein HSP20(B7G195), and a down-regulation of a heat shock transcription factor (B7FWU3), which corresponded to the f indings in plants. The synthesis of iron starvation-induced proteins (B7FYL2, B7G4H8) was also up-regulated under LO condition. These proteins usually related with silicon and iron starvation (Zhao et al., 2014, 2018).
The high expression of ornithine and arginine under hypoxic condition may also be one of the stress resistance mechanisms ofP.tricornutum. In plants, the existence of the urea cycle has been demonstrated in some species by the isotope tracer method (Slocum, 2005), but the evidence is limited.It is speculated that ornithine in chloroplasts is the initial substrate for the urea cycle in plants and is the precursor of polyamines; ornithine in mitochondria also participates in the synthesis of proline and other amino acids (Winter et al., 2015).Arginine is a crucial amino acid having several functions in plants. It is not only a component of the proteins, a medium for nitrogen storage and transport but also an osmotic regulator. Arginine not only participates in the synthesis of proline and nitric oxide (NO) through the urea cycle and the citrulline-NO cycle but also in the biosynthesis of polyamines (Winter et al., 2015). Thus, metabolic intermediates of the urea cycle play an important role in stress signaling transduction, regulation of ionic balance, generation of antioxidants and osmotic protective substances, the precursors of proline, polyamines, and NO biosynthesis.Additionally, it is directly involved in plant resistance through the synthesis of osmotic protective substances and antioxidants (Liu et al.,2015).
5 CONCLUSION
Under hypoxic condition, the growth rate ofP.tricornutumwas reduced. Carbon metabolism inP.tricornutumwas reset under an adverse environmental condition, such as the enhancement of lipid utilization. Other stress resistance mechanisms were activated to cope with hypoxia-induced adversity. Due to the diff erence in habitat as compared to the higher plants,P.tricornutumshowed metabolic alterations, which needs further investigation in the future.
6 DATA AVAILABILITY STATEMENT
The data that support the f indings of this study are available from the corresponding author upon reasonable request.
 Journal of Oceanology and Limnology2022年5期
Journal of Oceanology and Limnology2022年5期
- Journal of Oceanology and Limnology的其它文章
- Comparison of three f locculants for heavy cyanobacterial bloom mitigation and subsequent environmental impact*
- Eff ect of light intensity on bound EPS characteristics of two Microcystis morphospecies: the role of bEPS in the proliferation of Microcystis*
- Community structure of aerobic anoxygenic phototrophic bacteria in algae- and macrophyte-dominated areas in Taihu Lake, China*
- Tidal water exchanges can shape the phytoplankton community structure and reduce the risk of harmful cyanobacterial blooms in a semi-closed lake*
- Eff ect of random phase error and baseline roll angle error on eddy identif ication by interferometric imaging altimeter*
- Estimating the evolution of sea state non-Gaussianity based on a phase-resolving model*