
The eff ect of cylindrospermopsin on the bacterioplankton community: a microcosm experiment on water from Dishui Lake, Shanghai, China*
2022-10-08 01:16SichengYINRuohanHUANGErikJEPPESENLijingCHENLiqingWANGXinFANGWeiZHANG
Sicheng YIN , , Ruohan HUANG , , Erik JEPPESEN , Lijing CHEN , Liqing WANG ,Xin FANG , Wei ZHANG ,**
1 Engineering Research Center of Environmental DNA and Ecological Water Health Assessment, Shanghai Ocean University,Shanghai 201306, China
2 Department of Ecoscience, Aarhus University, Silkeborg 8600, Denmark
3 Sino-Danish Centre for Education and Research (SDC), University of Chinese Academy of Sciences, Beijing 100049, China
4 Limnology Laboratory and EKOSAM, Department of Biological Sciences, Middle East Technical University, Ankara 06800, Turkey
5 Institute of Marine Sciences, Middle East Technical University, Mersin 33731, Turkey
6 School of Business, Macau University of Science and Technology, Taipa, Macao 999078, China
Abstract Eutrophication and climate warming have intensif ied the global expansion of invasive cyanobacteria such as Cylindrospermopsis spp. and Chrysosporum spp. Cylindrospermopsin (CYN)produced by species of the latter two genera may harm phytoplankton, zooplankton, and f ishes. However,eff ects of CYN on the bacterioplankton community have not been studied. Based on high-throughput sequencing, we explored the eff ect of CYN on the structure and function of the bacterioplankton community by adding pure CYN to in- situ water collected from a brackish coastal shallow lake: Dishui Lake, China. We found that most bacterioplankton taxa had a certain tolerance potential to CYN, but that high concentrations of CYN (40 μg/L) caused a signif icant decrease in microbial abundance and functional groups. Of the dominant phyla, Actinobacteria had the strongest tolerance to CYN. Network analysis indicated that CYN caused shifts in the community structure of the bacterioplankton, reducing community stability and structural complexity. High CYN concentrations also reduced the correlation between the diff erent bacterioplankton groups, and the abundances of some bacterial taxa associated with the denitrif ication function and the process of carbon transfer in the microbial food web were inhibited. Our study provides new insight into the response of the bacterioplankton communities to harmful algal toxins produced by cyanobacteria.
Keyword: cylindrospermopsin; bacterioplankton; denitrif ication function; carbon cycles; Dishui Lake
1 INTRODUCTION
With global warming and the intensif ication of eutrophication of water bodies, the problem of cyanobacteria blooms has become increasingly serious(Yan et al., 2017). With these blooms, cyanobacteria metabolites enter the water column, aff ecting severely the aquatic ecosystem. Among the metabolites,cyanotoxins have become a central issue due to their strong toxicity and the environmental hazards that they pose (Briland et al., 2020), potentially inhibiting the growth of dominant aquatic organisms, such asMicrocystissp.,Lemnaminor,Daphniamagna, andWolffi aarrhizal(Jámbrik et al., 2010; Bednarska et al., 2014; Rzymski et al., 2014), and shifting the food web in lakes and oceans (Pohnert et al.,2007). The cyanotoxins accumulate in consumers up through the food chain, thereby aff ecting survival and reproduction (Wilson et al., 2006).Microcystisis the most common blooming cyanobacteria, and impacts of microcystins (MC), produced by these algae, on the aquatic ecosystem have been widely reported. Studies have shown that MC can aff ect the underwater organs of macrophytes and their growth(Chen et al., 2020), cause abnormal development of the freshwater snailLymnaeastagnalis(Lance et al., 2007, 2011) and inhibit the photosynthesis of microalgae (e.g.,Chlorellasp.), which ultimately inhibits the proliferation of algae (Singh et al., 2001;Campos et al., 2013).
In recent years, other cyanobacteria with a strong dispersal ability and invasive potential have been observed to develop blooms, e.g. in Chinese rivers,lakes, and reservoirs (Jiang et al., 2014; Lei et al., 2014).Among these, CYN-producingCylindrospermopsisandChrysosporiumhave attracted special attention(Fadel et al., 2014; Burford et al., 2016; Zhang et al.,2017) as they have shown strong competitiveness and may replaceMicrocystisunder certain conditions (Lei et al., 2014; Zhang et al., 2017), in part as result of CYN production (Zhang et al., 2017). Several studies have focused on the impact of CYN on various aquatic organisms. Beyer et al. (2009) found that CYN inhibited the growth of reed roots and caused necrosis of root epidermal cells. Zhu and Chen (2012)showed that CYN induced oxidative damage to grass carp lymphocytes, leading to cell apoptosis. Another investigation found that CYN stimulated sympatric phytoplankton to produce extracellular alkaline phosphatase (APA), thereby increasing the available phosphate content in the water column (Bar-Yosef et al., 2010). These results are helpful to understand the potential damage of CYN to aquatic ecosystems and how the CYN-producing species may dominate water bodies; however, knowledge about the response of microbial assemblages to CYN in aquatic ecosystems is limited.
As decomposers and consumers, bacteria play an important role in the material circulation and energy f low of the surface water ecosystem (Zhou et al.,2009). Moreover, when the lake water is exposed to contaminants, specif ic bacterial groups will be activated and degrade the toxic substances, thereby reducing or eliminating their harm (Hyenstrand et al.,2003). Because microorganisms are more sensitive to environmental changes and have a shorter growth cycle, they typically respond to environmental pollutants before other species (Bloem and Breure,2003). Studies have shown that cyanobacteria blooms and cyanotoxins aff ect the community structure and function of bacterioplankton in diff erent ways(Wilhelm et al., 2011). Non-toxic soluble organic carbon (DOC) released by cyanobacteria can be used as a carbon source to stimulate the growth of bacteria (Wang and Priscu, 1994), but cyanotoxins may also have a negative impact on bacterioplankton,for instance by reducing bacteria diversity or slowing down the process of the nitrogen cycle by inhibiting the growth of denitrifying bacteria (Yang et al., 2008;Li et al., 2016).
Dishui Lake, a coastal man-made lake with a total area of 5.56 km2, is located in the southeast of Shanghai, China (Lin et al., 2014). In recent years,the CYN-producing cyanobacteriaChrysosporumovalisporumhas been observed in the lake, with eff ects on the phytoplankton and zooplankton communities(Zhang et al., 2018). In this study, we aimed to explore the eff ect of CYN on the structure and function of the bacterioplankton community in Dishui Lake in a microcosmic experiment involving addition of pure CYN and high-throughput sequencing analysis. We hypothesised that 1) CYN can inhibit the growth of bacterioplankton and thus change their community structure and 2) the high concentrations of CYN can shift the functional taxa in the aquatic ecosystem.Our study provides new insight into the relationship between heterotrophic bacteria and cyanobacterial toxins and, thus, the carbon and nitrogen cycles in shallow lakes.
2 MATERIAL AND METHOD
2.1 Exposure experiment of planktonic microorganisms to CYN
In September 2019, surface water samples were collected at 0.5-m depth in a nearshore zone of Dishui Lake, an area without cyanobacteria blooms.In-situ water temperature (WT), pH, dissolved oxygen (DO), and salinity (SAL) were measured using an YSI Pro-plus multisensor sonde. Other water quality parameters, total phosphorus (TP), and chlorophylla(chla) were measured simultaneously,and the results of these measurements can be found in our previous study (Zhang et al., 2018). Average values of physical and chemical parameters of Dishui Lake are given in Table 1.
The water samples were kept cold at 4 ℃ and transported to the laboratory where they were mixed evenly and transferred into 1-L beakers. The sampleswere tested for CYN before our experiments, and no CYN was detected. Three concentration levels of pure CYN (Enzo life science): low group (6 μg/L),medium group (20 μg/L), and high group (40 μg/L)were set up in the exposure experiment. The initial lake water without CYN was used as control. The f inal culture volume was 500 mL. All the beakers were kept at 25±1 ℃ under a light intensity of 2 500 lx(12-h dark∶12-h light cycle). Each treatment group had three replicates, and the beakers were manually shaken three times a day during the experimental cycle.

Table 1 Water quality and physicochemical properties of Dishui Lake in September 2019 shown as average± standard deviation
After 48 hours of incubation, 300 mL of the mixed water sample was removed and f iltered through 5-μm and 0.2-μm f ilters. The f ilter membrane was placed in a sterile centrifuge tube, quickly frozen with liquid nitrogen and then stored in a refrigerator at -20 °C.The samples were then forwarded to Metabio Science& Technology Co., Ltd. (Wuxi, China) for Miseq(Illumina) high-throughput sequencing.
2.2 Sample DNA extraction
Metagenomic DNA was extracted by PowerSoil®DNA Isolation Kit from MOBIO Laboratories,Inc., according to the manufacturer’s instructions.UV spectrophotometry was used to detect the concentration and purity. The specif ic method was applied following the instructions of the kit.
2.3 PCR amplif ication and sequencing analysis
In this study, the variable region of bacterial 16S rDNA V3-V4 was selected as the target amplif ication fragment for the subsequent highthroughput sequencing, and 16S rDNA V3-V4 universal primer 341F/805R was chosen as extract. The primer sequences were 341F: 5′-CCCTACACGACGCTCTTCCGATCTG-3′, 805R:5′-GACTGGAGTTGGCACCCGAGAATTCCA-3′,and Barcode discrimination sequence samples were added at the end of 3′ (Caporaso et al., 2011). PCR reaction conditions were: pre-denaturation at 98 °C for 30 s, denaturation at 98 °C for 15 s, annealing at 50 °C for 30 s, extension at 72 °C for 30 s, 25-27 cycles, incubation at 72 °C for 5 min; and storage at 4 °C (Biorad C1000 Touch PCR, Shanghai).
The products were detected by electrophoresis 2%agarose gel, and the Axygen gel recovery kit recovered the target fragments. Quant-i T PicoGreen dsDNA Assay Kit was used to quantify the PCR products on the Microplate reader (BioTek, FLx800), and then the samples were mixed according to the amount of data required for each sample.
The original image data f ile obtained from Illumina Miseq sequencing was transformed into raw reads by base calling analysis. Each sample was distinguished with a barcode, and the sequence quality was controlled and f iltered. The sequence was spliced according to the overlap relationship, and the spliced sequence was subjected to quality control and was f iltered again. Finally, the optimized sequence(total: 2 309 479, average: 192 457 per sample, max:300 694, min: 43 461) was obtained for downstream analysis. The optimized sequences were analyzed by Operational Taxonomic Units (OTUs) cluster analysis and species taxonomy. All sequences were divided into OTUs relative to diff erent similarity levels.In this study, the biological information of OTU at 97% similarity level was statistically analyzed, and OTUs with an abundance below 2 were removed.The sequences and related date were deposited into the National Center for Biotechnology Information(NCBI) database with the Sequence Read Archive(SRA) accession number of SRR18810468.
2.4 Statistical analysis
The results were handled in Past V3.22 and R
4.0.2 software. We calculated the α diversity index in the Past V3.22 program. The results of Bray-Curtis distances were visualized by Principal Coordinate Analysis (PCoA) plots in the R 4.0.2 environment using the “ape” package, and signif icant diff erences in bacterioplankton community structure among diff erent treatment groups were tested using one-way analysis of similarities (ANOSIM). The species with an absolute abundance greater than 10 in each group were selected for analysis of the co-occurrence network(Carey et al., 2017). For the observed and simulated networks, we calculated structural properties that assessed network complexity. These characteristics included the number of nodes and edges (i.e., taxa richness), connectance (the proportion of potential connections in a network that are actually connected),average degree (average number of connected edges per node), average path length (the mean shortest path length between all nodes), betweenness centralization(the ratio of the number of paths passing through this node among all the shortest paths to the total number of shortest paths), degree centralization (measuring node centrality), clustering coeffi cient (a metric of how connected nodes are in the network), and diameter (maximum value among shortest distances)(Newman, 2003; Kara et al., 2013; Carey et al., 2017;Zhu et al., 2021). We used the “Psych” package in R 4.0.2 to calculate inter-species correlations.Networks were visualized with the software programs Cytoscape v.2.8.3 and Gephi 0.9.2. Venn charts and heat maps were made using the software Origin 2016.Bar charts were mapped using Excel.
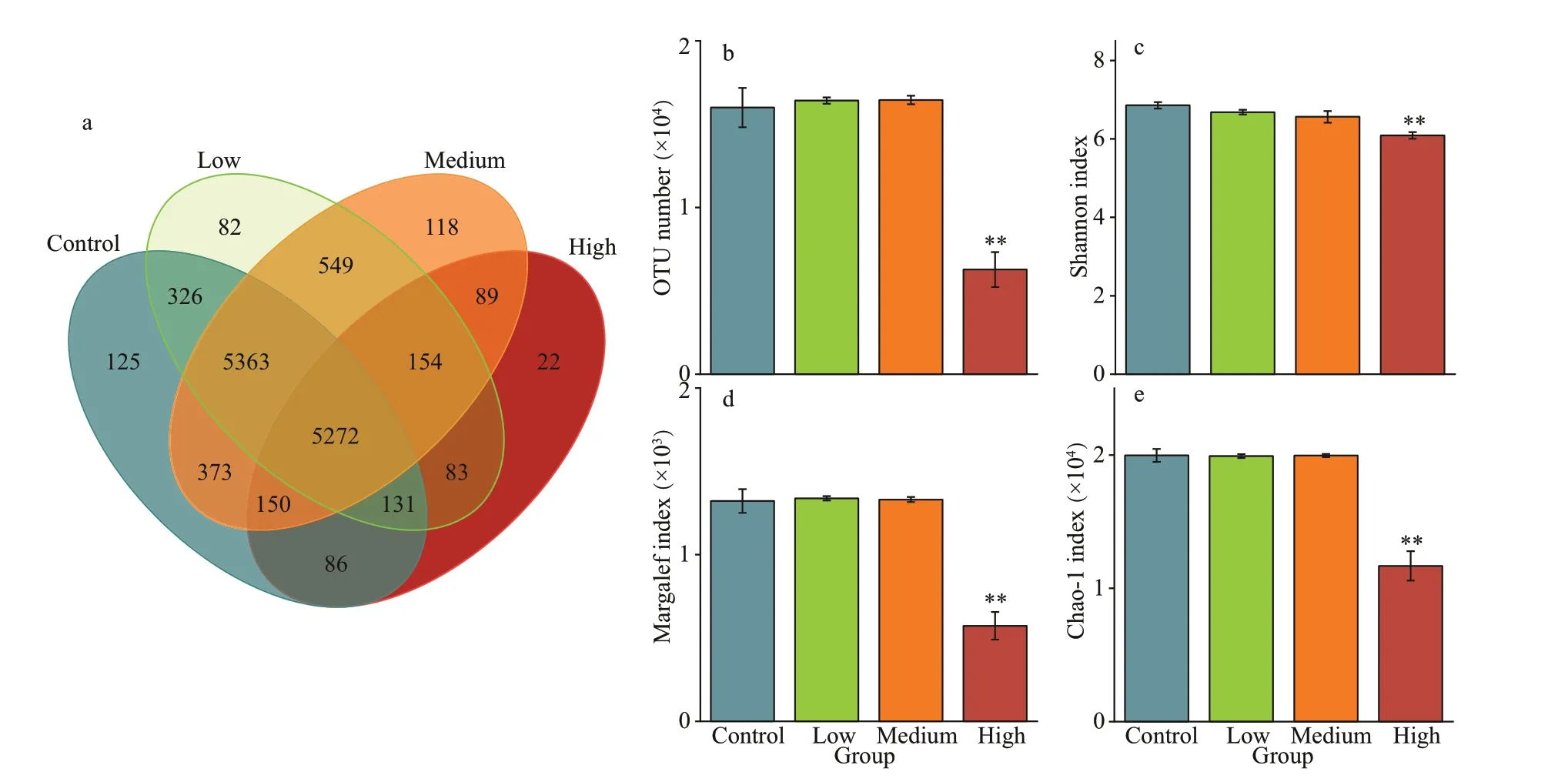
Fig.1 OTUs overlapping Venn charts and α diversity analysis
3 RESULT
3.1 Bacterial taxa diversity
The OTUs of the four groups were 20 748, 20 834,20 956, and 11 386, respectively, and 5 272 taxa were common for all four groups. The control group had the highest (n=125) taxa richness of particular taxa(occurring only in one group), but shared 11 092,11 158, and 5 639 OTUs with the low, medium,and high CYN groups, constituting 53%, 53%, and 50% of the total taxon number, respectively (Fig.1a).By contrast, the three treatment groups had higher numbers of shared taxa, the highest (n=11 338) being shared between the low and medium CYN groups(Fig.1a). With increasing CYN concentrations, the number of unique OTUs in each treatment group gradually decreased.
Compared with the control group, the number of OTUs in the low and medium CYN groups tended to be higher, though not signif icantly so (P>0.05), while the number of OTUs in the high CYN group was signif icantly lower (P<0.01). The Shannon index,the Margalef index, and the Chao-1 index of the high CYN groups were also signif icantly lower than those of the control group (P<0.01) (Fig.1b-e).
3.2 Abundance and composition of bacterioplankton
The total bacteria abundances in the low and medium CYN groups were 1.17 and 1.29 times that of the control group, but the diff erence was not signif icant (P>0.05). Contrarily, the bacteria abundance in the high CYN group was signif icantly lower (68.3%) than in the control group (P<0.05).A diff erent response pattern to CYN was found for the clades, phyla. The relative abundances of Proteobacteria and Comamonadaceae in the high CYN group were signif icantly lower than those of the other three groups (P<0.05 andP<0.01). Unlike Proteobacteria, the diff erence in absolute abundance of actinobacteria between the four groups was not signif icant (P>0.05) (Fig.2). Compared with the other groups, the relative abundance of Methylophilaceae was signif icantly higher in the medium CYN group(P<0.01) (Fig.3).
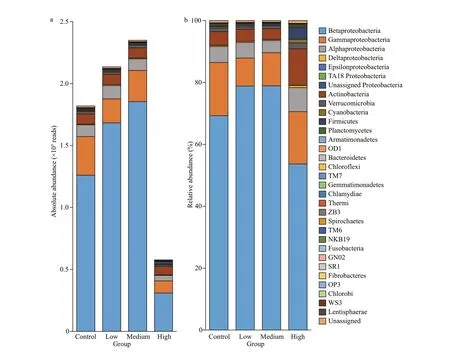
Fig.2 The abundance and relative abundance of the bacterioplankton community at phylum or class level
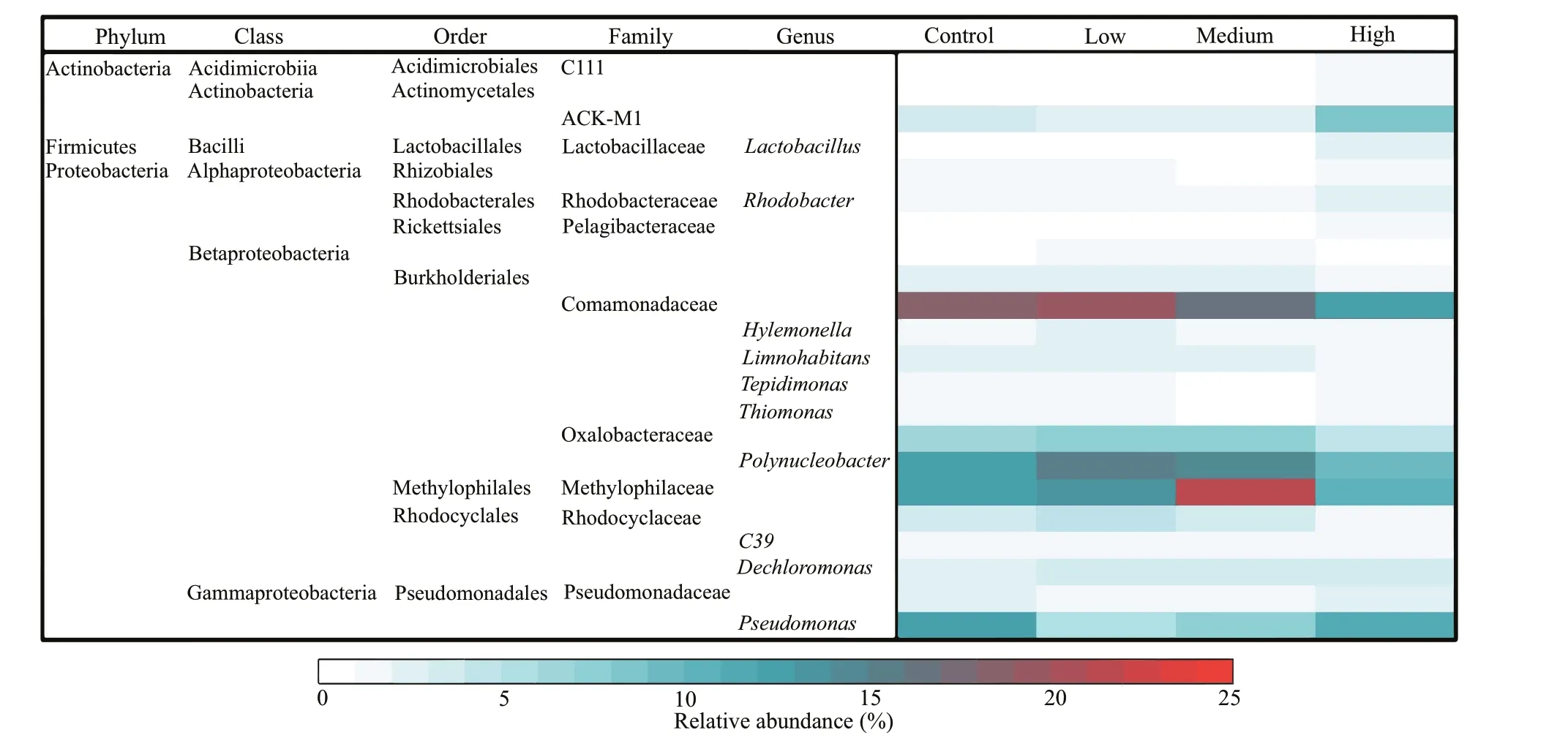
Fig.3 The relative abundance of the dominant genera
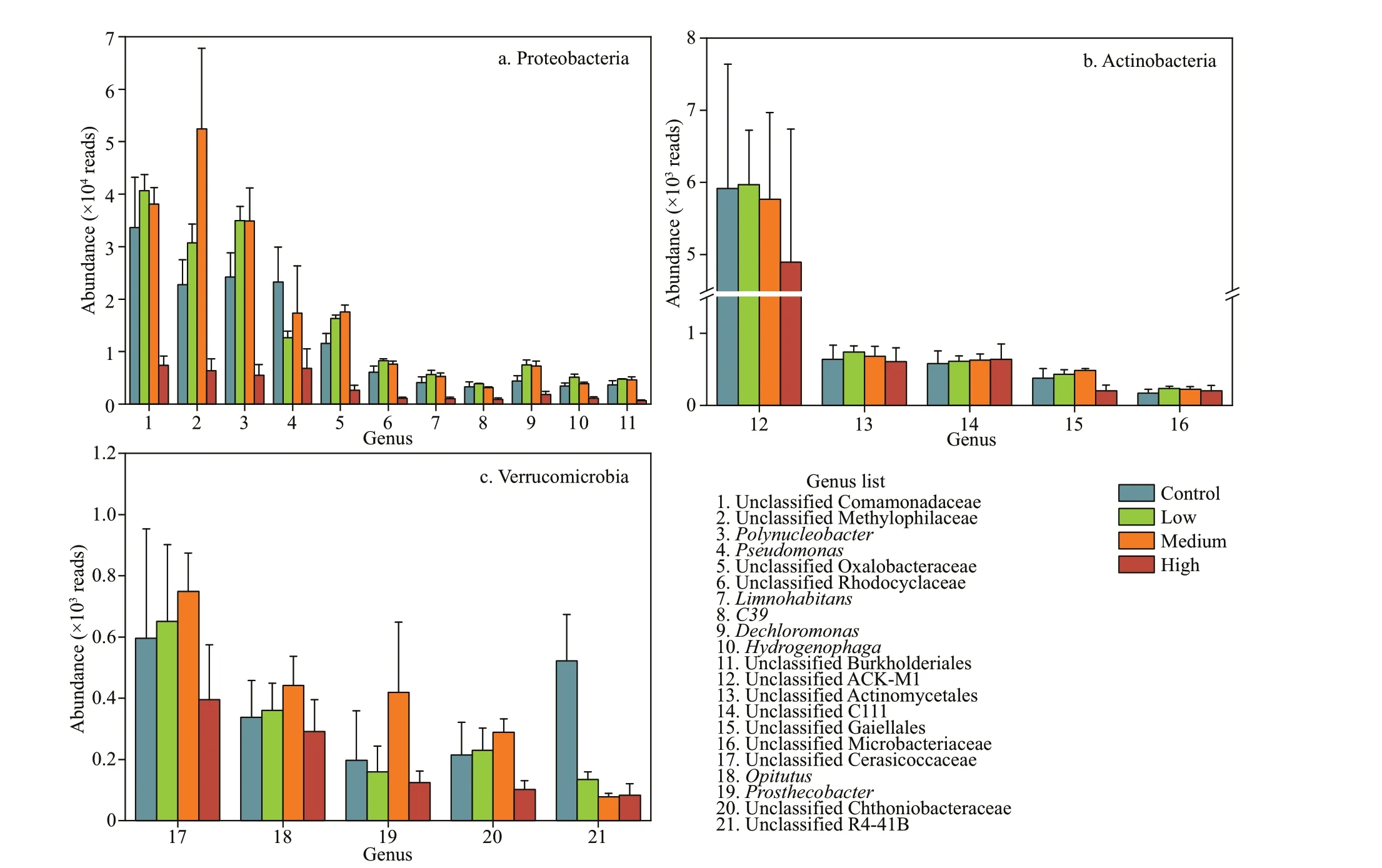
Fig.4 The abundances of the dominant genera of Proteobacteria, Actinobacteria, and Verrucomicrobia
Compared with the control group, the abundances of the most dominant genera increased in the low and medium concentration groups and decreased in the high concentration group. The abundances ofPseudomonas, unclassif ied Comamonadaceae, and unclassif ied R4-41B were signif icantly lower in the high CYN group than in the control group (P<0.05)(Fig.4).
3.3 Changes in planktonic bacteria community structure
The PCoA results showed that CYN exposure,especially at the high concentration level, changed the community of the main bacteria clades. Bacteria,Proteobacteria, and Planctomycetes in the high CYN group were signif icantly diff erent from the other groups (P<0.05). Actimobacteria, Verrucomicrobia,Firmicutes, and Cyanobacteria in the three treatment groups were not signif icantly diff erent from those of the control group (P>0.05). Armatimonadetes were signif icantly diff erent from the control group and here also diff erences between the high CYN group and the other treatment groups were found (P<0.05)(Fig.5).
3.4 Network analysis
The taxa from Proteobacteria and Actinobacteria were closely related to other phyla, while OD1 and Armatimonadetes were less related to other phyla(Fig.6). The bacteria communities in the control group exhibited signif icantly higher network complexity than the CYN treatment groups as indicated by more nodes (i.e., greater richness) and edges, higher network connectance and average degrees (Fig.7a-d).The transitivity among taxa was poorer in the high CYN group than in the other groups as suggested by lower clustering coeffi cients and larger diameters(Fig.7h-i). However, community stability tended to increase with rising CYN concentrations, reaching relatively stable conditions in the high CYN group as indicated by connectance, betweenness centralization,and degree centralization (Fig.7c, f, & g).
4 DISCUSSION
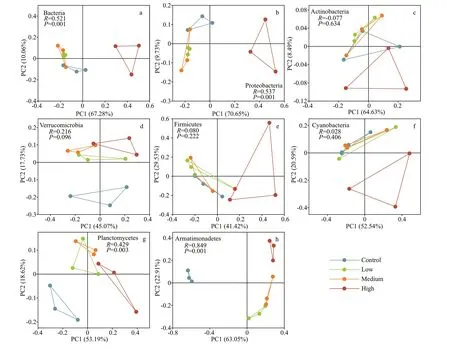
Fig.5 PCoA analysis diagram of diff erent dominant phyla based on Bray-Curtis distance
In aquatic ecosystems, cyanotoxins can inhibit the growth of other aquatic organisms and promote their own growth (Valdor and Aboal, 2007). Our results showed that most bacterioplankton taxa in Dishui Lake exhibit a certain tolerance to CYN, but a high CYN concentration (40 μg/L) led to signif icant decrease in the abundance of the dominant microbes,such as Comamonadaceae (Fig.4). CYN can inhibit protein and glutathione synthesis and cause gene damage (Campos et al., 2013). Glutathione is the most important non-protein sulfhydryl compound in bacteria cells and plays an anti-oxidant stress role(Copley and Dhillon, 2002). Strong oxidative stress produced by high concentrations of CYN may be one of the reasons for the signif icant decrease in bacteria abundance. We found that Actinomycetes were most resistant to CYN (Figs.4-5). A variety of actinobacteria (e.g., taxa belonging to Streptomyces)have the ability to dissolveMicrocystiscells and degrade microcystins (Choi et al., 2005). The high tolerance of actinobacteria to CYN indicates that they also have a potential for degrading CYN. Although a review showed that CYN concentrations may exceed 100 μg/L (Rzymski et al., 2014), most of the f ield survey results from lakes or reservoirs have shown CYN concentrations underCylindrospermopsisblooming events <10 μg/L (Fadel et al., 2014; Lei et al., 2014; Lu et al., 2021). We found that CYN increased the abundance of bacterioplankton at 6- and 20-μg/L CYN, thus showing “hormesis”, as also found in toxicological experiments elucidating the impact of antibiotics, microcystins and other substances on microorganisms (Chen and Jiang, 2011). Another investigation showed that CYN had allelopathic eff ects and stimulated other cyanobacteria to produce alkaline phosphatase (APA) (Rzymski et al., 2014).We speculate that the low and medium dosages of CYN in our experiment may have promoted the release of APA by bacterioplankton and that the increase of soluble inorganic phosphorus in the water column promoted the growth of these organisms.
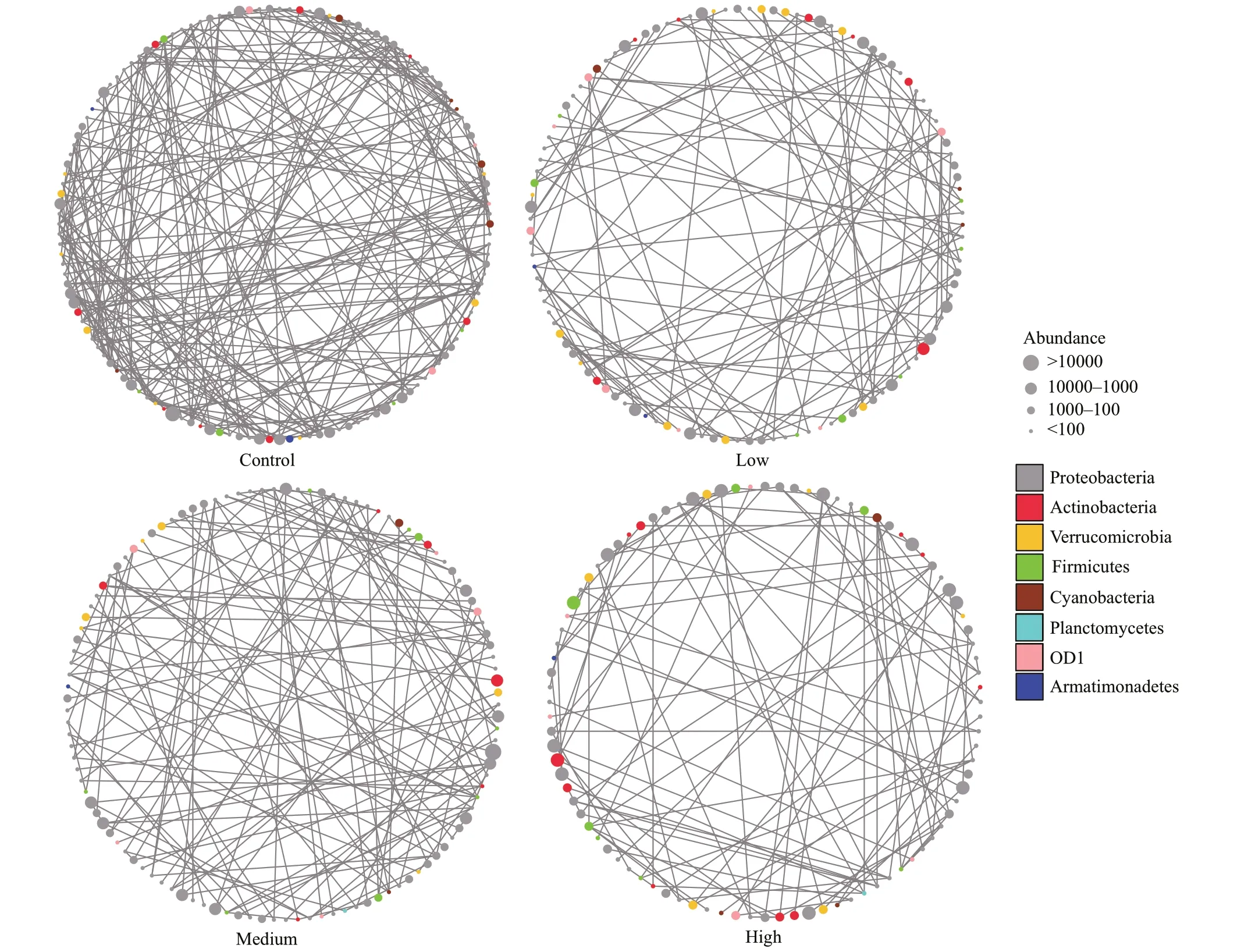
Fig.6 Observed networks of bacterioplankton co-occurrences in the four CYN concentration treatments (0, 6, 20, and 40 μg/L)
The diversity and community structure of bacterioplankton are closely related to their function.Our network analysis indicated that CYN caused shifts in the community structure of bacterioplankton,reducing their structural complexity, which subsequently aff ected their functions. We found that both the absolute and relative abundance of Comamonadaceae and Rhodocyclaceae decreased signif icantly in the high-CYN group (Fig.4), and many of their taxa (e.g.,Dechloromonas) play an important role in the denitrif ication process (Takahashi et al., 2011; Tian et al., 2015; Dong et al., 2019). The denitrif ication process is the key approach to transform nitrogen from reactive nitrogen to inert nitrogen with implications for the nitrogen cycle and global nitrogen balance (Murray and Knowles, 2003; Huang et al., 2017). A previous study by Yang et al. (2008)revealed that high concentrations of MC can inhibit denitrifying bacteria. However, in their study, the MC concentration had to reach mg/L level to produce the inhibitory eff ect, rarely occurring in natural waters. In our experiment, CYN had a signif icant inhibitory eff ect on denitrifying microorganisms(e.g., Methylophilaceae) at the μg/L level, suggesting a more potent eff ect on denitrif ication. In addition,among the several genera signif icantly inhibited by CYN, Pedosphaerales, mainly found in the sediments,plays an important role in the carbon cycle (Shi et al.,2018). Studies have also found that CYN can aff ect the predation of microorganisms by protozoa, thereby impacting the carbon transfer in the microbial food web (Bagatini et al., 2014).
We found that the high concentration of CYN could reduce the transmissibility of the bacterioplankton community (Fig.7), indicating a lessening of the inter-species interaction among bacterioplankton (Feng et al., 2017), and thus a weakening of the “small world”nature of the bacterioplankton community (Carey et al., 2017). Generally, the stability of a weaker small world network is less aff ected by the hub taxa (Carey et al., 2017). This may explain the recovery of the stability of the bacterioplankton community with increasing CYN concentrations (Fig.7).
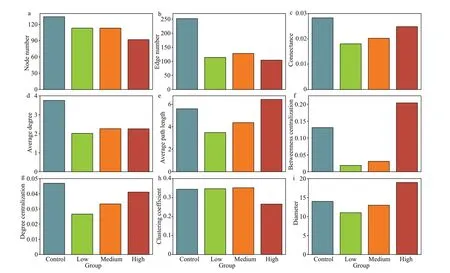
Fig.7 Parameters of the networks of bacterioplankton co-occurrences in Fig.6
5 CONCLUSION
Our results indicate that the high concentrations of CYN (40 μg/L) aff ected the abundance and the complexity and stability of the community structure of bacterioplankton in the water column, and that Proteobacteria and Verrucomicrobia were more sensitive to CYN exposure than Actinobacteria. Our results also reveal the great potential of CYN in shifting the microbial loop function in aquatic ecosystems under blooms of CYN-producing cyanobacteria.
6 DATA AVAILABILITY STATEMENT
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT
We would like to express our deep thanks to Anne Mette Poulsen from Aarhus University for English assistance.
 Journal of Oceanology and Limnology2022年5期
Journal of Oceanology and Limnology2022年5期
- Journal of Oceanology and Limnology的其它文章
- Comparison of three f locculants for heavy cyanobacterial bloom mitigation and subsequent environmental impact*
- Eff ect of light intensity on bound EPS characteristics of two Microcystis morphospecies: the role of bEPS in the proliferation of Microcystis*
- Community structure of aerobic anoxygenic phototrophic bacteria in algae- and macrophyte-dominated areas in Taihu Lake, China*
- Tidal water exchanges can shape the phytoplankton community structure and reduce the risk of harmful cyanobacterial blooms in a semi-closed lake*
- Eff ect of random phase error and baseline roll angle error on eddy identif ication by interferometric imaging altimeter*
- Estimating the evolution of sea state non-Gaussianity based on a phase-resolving model*