
栗斑腹鹀和三道眉草鹀物种分化研究
2022-09-28 09:32:14张立世尚伟平严蓉飞姜云垒
东北师大学报(自然科学版) 2022年3期
张 雪,张立世,尚伟平,严蓉飞,姜云垒,李 时
(1.吉林农业大学生命科学学院,吉林 长春 130118;2.吉林农业大学动物科学技术学院,吉林 长春 130118)
隔离是物种分化的条件之一.一般而言,新物种的形成,先是不同种群间由于某种原因发生了地理隔离,而后各种群内的基因型和表型受到选择压力的影响而朝着不同的方向发生适应性进化,最终出现生殖隔离形成新的物种.但对于分布区域距离较近的物种而言,性别隔离则可能是促进其发生分化的主要原因.
物种形成是一个复杂而又漫长的过程[1].传统的理论认为,地理隔离是物种形成的前提,但越来越多的研究表明,地理隔离并不是影响物种形成的唯一因素,尤其是对于扩散能力较强的鸟类而言,即使是在有基因流存在的情况下也能发生分化[2].近年来,研究者们已通过分子钟推测出古气候的变化也是驱动物种分化的一个因素,例如:由于更新世冰期-间冰期的气候波动以及冰期避难所的存在,导致不同避难所内的动物种群发生了独立的进化,进而形成了新的物种[3].有研究[4]表明,分布于北方地区的鸟类更易受到第四纪冰期的影响而被隔离在不同的破碎化地理分布区内发生分化.此外,生态因子也是驱动物种形成的一个重要因素,在没有明显的地理屏障存在的条件下,生活在不同环境中的同一物种的不同种群,由于对不同环境条件的适应而产生了遗传分歧,甚至出现生殖隔离形成新的物种[5-6].然而,对于分化时间未超过200万年的年轻物种,在发生二次接触时,可能会发生杂交并产生可育后代[7],也可能将特定性状不对称地从一个物种向另一个物种渗透[8-10].
历史上,栗斑腹鹀(Emberizajankowskii)曾是一种常见鸟类[11-12],主要分布区包括中国、俄罗斯和朝鲜[12-13].在过去50年间,由于栖息地丧失和破碎化等原因导致栗斑腹鹀个体数量剧减,已于2010年被世界自然保护联盟(IUCN)列为濒危物种[14],现存栗斑腹鹀种群主要分布于我国内蒙古的中东部地区.
三道眉草鹀(E.cioides)和栗斑腹鹀同为鹀属鸟类,已有研究[15-16]表明二者为形态相似的近缘物种,且在栗斑腹鹀分布区内为同域分布物种.然而,驱动二者发生物种分化的主要原因,以及第四纪冰期是否对二者物种分化产生了影响仍不清楚.为此,本文通过分子钟追溯了二者的分化时间,进而推测了驱动二者间发生物种分化的历史原因;同时,通过形态特征和鸣声特征研究了性别隔离对其物种分化的影响,研究结果将有助于更深入地认识和了解生物演化的过程,并为第四纪冰期在物种分化中的作用提供更为广泛的科学依据.
1 材料和方法
1.1 野外采样和数据收集
2012—2019年每年的4—7月,在内蒙古自治区的通辽市和赤峰市等地,对栗斑腹鹀和三道眉草鹀的繁殖情况进行调查,并采集形态数据、鸣声数据和血液样本.
在繁殖期内,栗斑腹鹀和三道眉草鹀的雄鸟常站在树梢进行警戒,利用便携式数字录音机(TASCAM HD-P2)配以强指向话筒(Sennheiser MKH416 P48)对其进行录音,收集两种鸟的鸣声数据.为避免对栗斑腹鹀和三道眉草鹀的繁殖产生过大影响,待雏鸟7~8日龄时,利用雾网捕捉成鸟,在翼下肱静脉采集成鸟和雏鸟的血液样本,每巢采集3~5只雏鸟.将采集到的血液样本(10~20 μL/只)与1 mL无水乙醇混合于1.5 mL已灭菌的离心管中,于-20℃冰箱或液氮中保存,带回实验室后保存于-80℃冰箱中.繁殖期结束后,利用雾网捕捉成鸟,并用电子秤和数显游标卡尺对捕获成鸟的体重、体长、头喙长、喙长、喙宽、喙高、头长、头高、头宽、尾长、翅长、跗跖长、爪长和中趾长,共14个主要形态参数进行测量,收集形态数据.
收集完形态数据和血液样本后,将成鸟原地放飞,并将雏鸟重新放回到所在巢中.根据后续的跟踪观察,采集血样和形态测量均未造成雏鸟或成鸟的死亡.
2012—2019年,共采集到234份栗斑腹鹀血液样本和60份三道眉草鹀血液样本.2017—2019年,一共录制到131只栗斑腹鹀的1 254个鸣声和77只三道眉草鹀的759个鸣声.共对79只栗斑腹鹀和20只三道眉草鹀成鸟的主要身体指标进行了测量.
1.2 鸣声和形态数据分析
首先,利用SPSS中非参数检验的1-样本K-S(Kolmogorov-Smirnov)检验,检测所用形态数据和鸣声数据是否符合正态分布.
对所有个体的形态数据进行双尾T检验(2-TailedtTest),判别其性别二态性.因为栗斑腹鹀和三道眉草鹀在繁殖期内主要是雄鸟站在枝头鸣唱、鸣叫和报警,因此录制的鸣声均为雄鸟鸣声.由于本文所使用的均为雄鸟的鸣声数据,因此,未对鸣声数据进行性别二态性检测.
如研究数据参数符合正态分布,则利用独立样本T检验(Independent Samples Test)判定不同物种间各形态和声信号参数是否存在显著差异,根据给出的Levene方差齐性检验结果判断方差是否相等.如方差相等(P>0.05),则依据假设方差相等时的显著值判断;如果方差不相等(P<0.05),则依据假设方差不相等时的显著值判断.
如研究数据不符合正态分布,则利用非参数检验中的两独立样本检验进行检测.
选取61只录音效果好(栗斑腹鹀30只,三道眉草鹀31只),且每只个体均具有10条以上可用声波数据的样本进行统计分析.鸣声数据的非参数检验结果显示,所有的鸣声数据均符合正态分布(P>0.05).
选择2018—2019年记录的14个形态参数的个体用于后续研究,包括栗斑腹鹀51只(雌性26只、雄性25只),三道眉草鹀19只(雌性12只、雄性7只).各形态参数的1-样本K-S正态分布检验结果表明,栗斑腹鹀各形态参数中,除头喙长和中趾长外,其余各形态参数均符合正态分布(P>0.05);三道眉草鹀各形态参数均符合正态分布(P>0.05).
利用SPSS 19.0软件对栗斑腹鹀和三道眉草鹀的主要形态和鸣声特征参数进行聚类分析,并构建聚类图.
1.3 DNA提取和线粒体ND2基因扩增
利用AxyPrep血液基因组DNA试剂盒和苯酚-氯仿法,对234份栗斑腹鹀和60份三道眉草鹀血液的全基因组进行提取,并参考文献[17]给出的引物和方法进行PCR扩增.
所有的PCR扩增产物均用1%的琼脂糖凝胶,在装有1×TAE缓冲液的电泳槽中进行电泳检测,所用标志物(Marker)为DL 2000.扩增所得样本送生物公司进行测序.
1.4 遗传距离和系统发育分析
测序所得序列用BioEdit version 7.2.5和Editseq version 5.0软件进行编辑.用Clustal X ver.1.8软件对测序所得的线粒体ND2基因进行比对,并对比对后的序列辅以人工校正,截取起始密码子(AUG)和终止密码子(UAA,UGA,UAG)之间的序列用于后续的数据分析.
分别利用PHYML软件中的最大似然法(maximum likelihood,ML),PAUP* ver.4.0b10软件中的邻接法(neighbor joining,NJ)和最大简约法(maximum parsimony,MP),以及MrBayes ver.3.1.2软件中的贝叶斯法(Bayesian inference,BI)重建基于线粒体ND2单倍型序列的ML,MP,NJ和BI系统发育树.利用PopArt1.7软件构建栗斑腹鹀和三道眉草鹀的单倍型网格图.
1.5 物种分化时间
基于线粒体ND2基因序列,用BEAST version 1.6.2软件估算两个物种最近的共同祖先时间.ND2的突变率参考文献[17]关于黄鹀和白头鹀的研究,采用每百万年2%的突变率.应用jModeltest 2.1.10软件计算所得的栗斑腹鹀和三道眉草鹀单倍型的最优碱基替代模型为TIM2+I,而在Bayesian Evolutionary Analysis Utility Version(BEAUti)v1.6.2平台中并没有TIM模型,因此,本文采用与最优模型最相似的碱基替代模型.估算分析共运行3次,每次运行100万次,采样频率为1 000次/代,开始的10%作为“老化(burn-in)”结果舍弃.最终的结果在TRACER version1.5平台中合并后进行分析.
2 结果
2.1 栗斑腹鹀和三道眉草鹀的鸣声分化
基于物种间鸣声数据的持续时间、最高频率、最低频率和时间间隔4个参数的分析发现,栗斑腹鹀和三道眉草鹀鸣声间除持续时间外,均不具有显著性差异,其中三道眉草鹀鸣唱语句持续时间更长(见表1),说明栗斑腹鹀和三道眉草鹀在语句结构上存在差异,需进一步研究揭示二者语句结构上的差异.
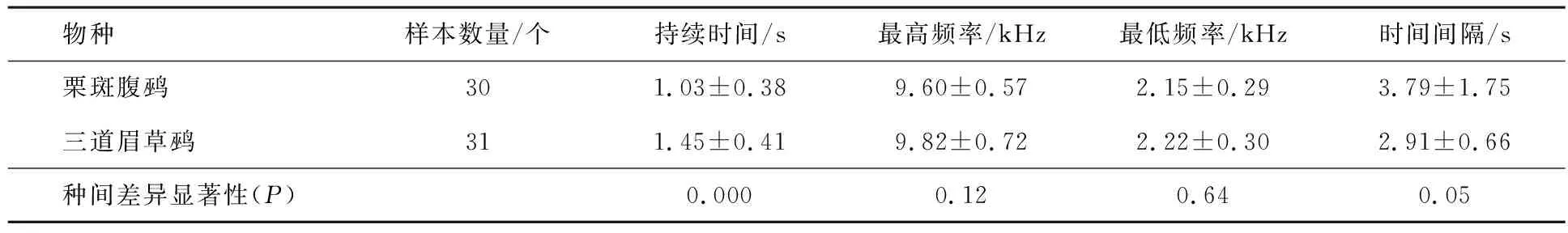
表1 栗斑腹鹀和三道眉草鹀鸣声特征(平均值±标准差)
基于不同区域栗斑腹鹀种群间鸣声数据的持续时间、最高频率、最低频率和时间间隔4个参数的分析发现,查干套力皋和嘎亥图两地分布的栗斑腹鹀个体除时间间隔外,均不具有显著差异(见表2).
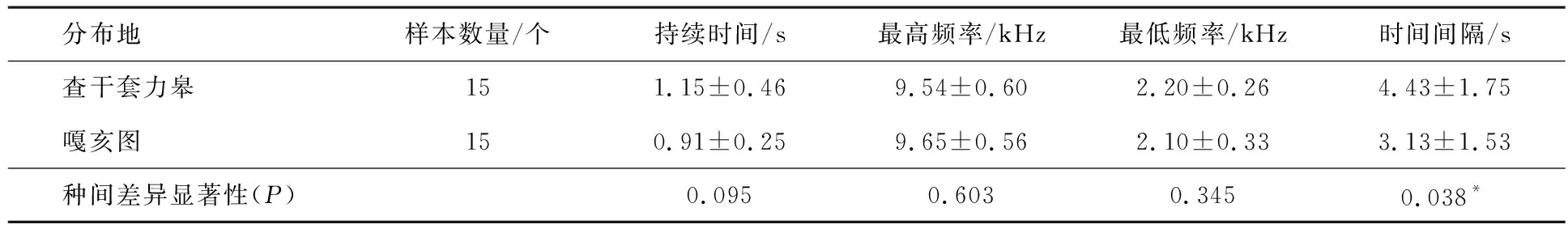
表2 查干套力皋栗斑腹鹀和嘎亥图栗斑腹鹀鸣声特征(平均值±标准差)
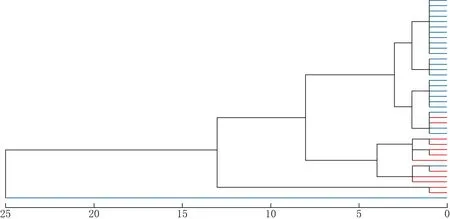
利用栗斑腹鹀和三道眉草鹀的鸣声数据进行聚类分析,结果见图1.由图1可见,61只个体样本聚成了2个主要分枝(A和B),然而聚类分析并没有将栗斑腹鹀和三道眉草鹀完全分开,在分枝A中,栗斑腹鹀和三道眉草鹀聚在了一起.
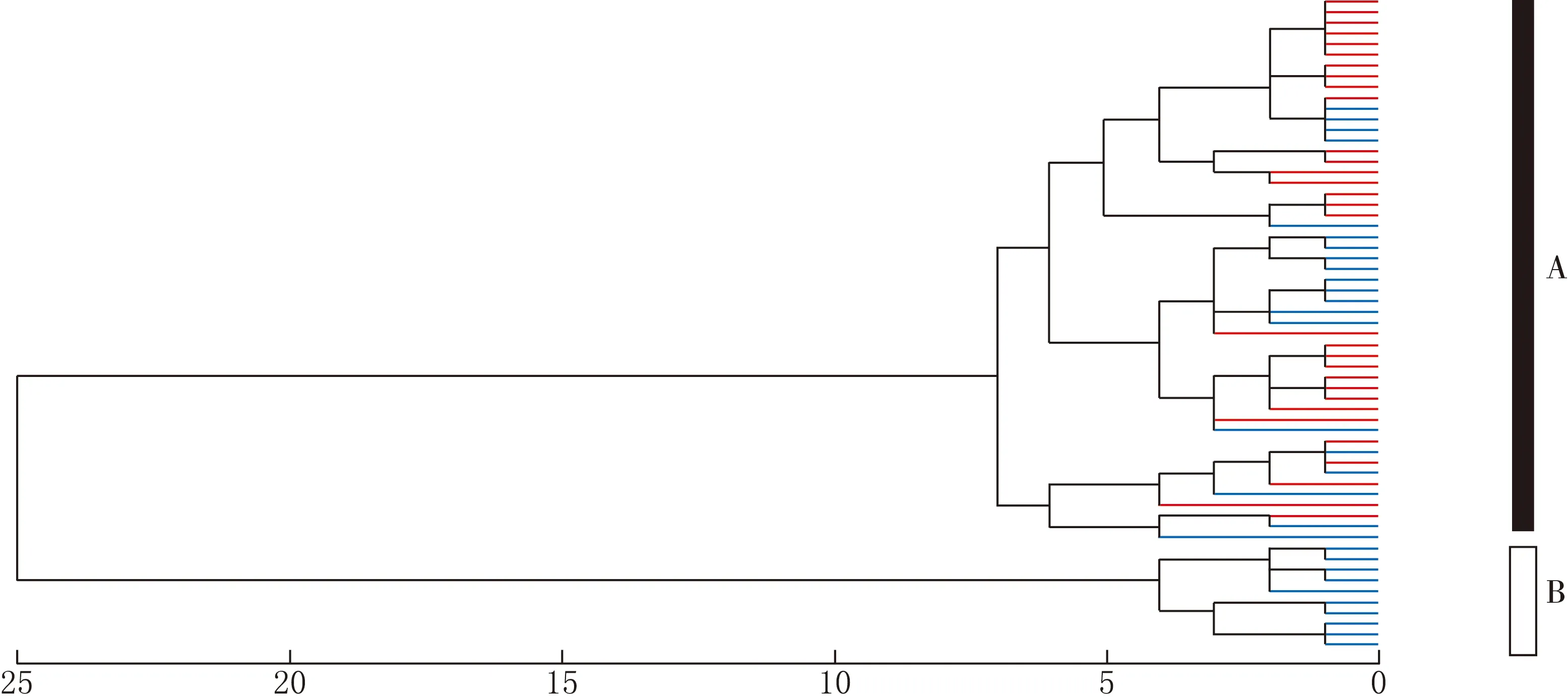
红色为三道眉草鹀,蓝色为栗斑腹鹀
2.2 栗斑腹鹀和三道眉草鹀的形态分化
基于物种间形态数据的分析发现,栗斑腹鹀的翅长、跗趾长、体长、体重、头喙长和尾长在雌、雄间存在显著差异,其他形态参数雌雄间无显著差异(见表3);三道眉草鹀的翅长、体长和尾长在雌、雄间存在显著差异,其他形态参数雌、雄间无显著差异(见表3).二者在主要形态参数上均表现为雄性个体较雌性个体略大.
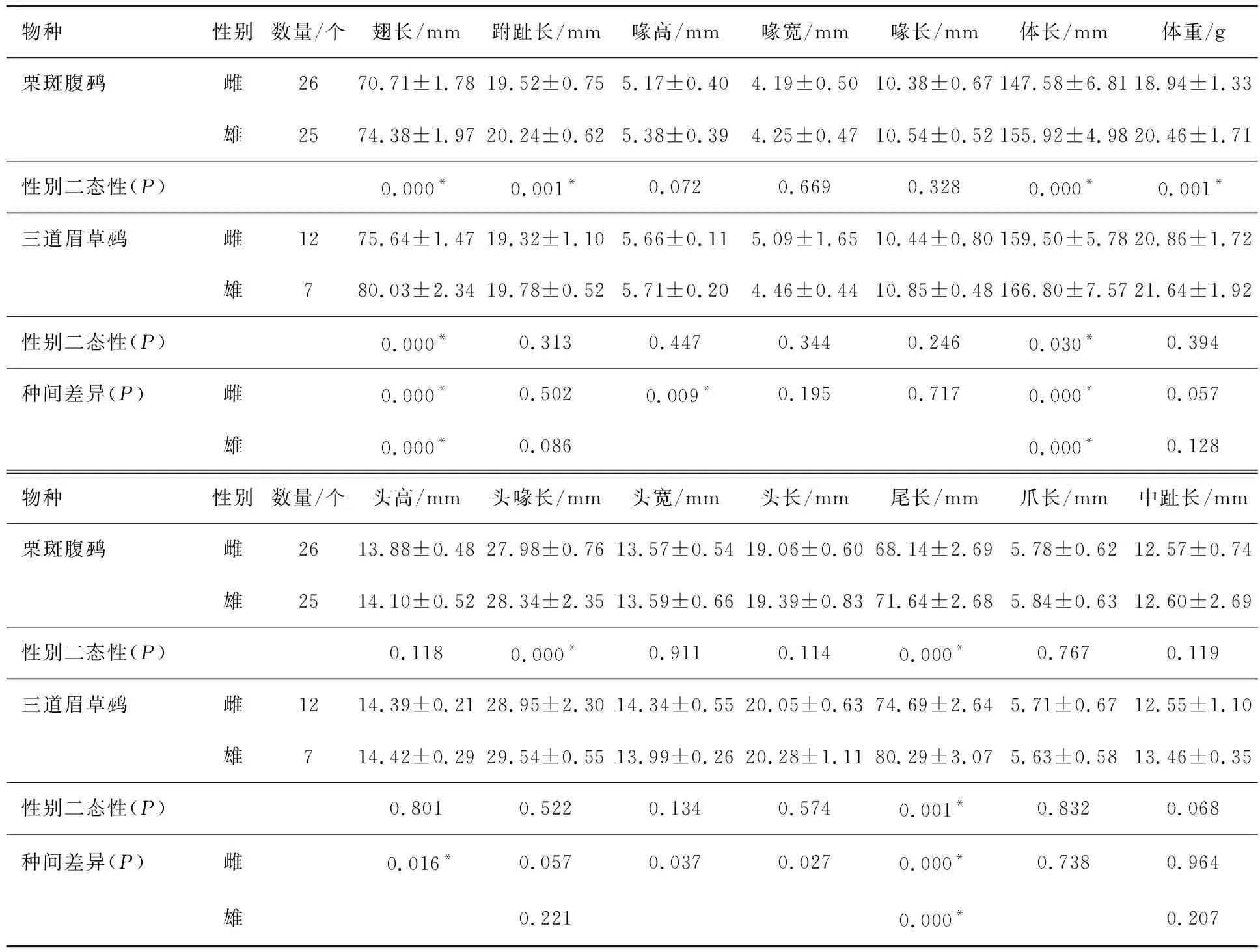
表3 栗斑腹鹀和三道眉草鹀外部形态特征比较(平均值±标准差)
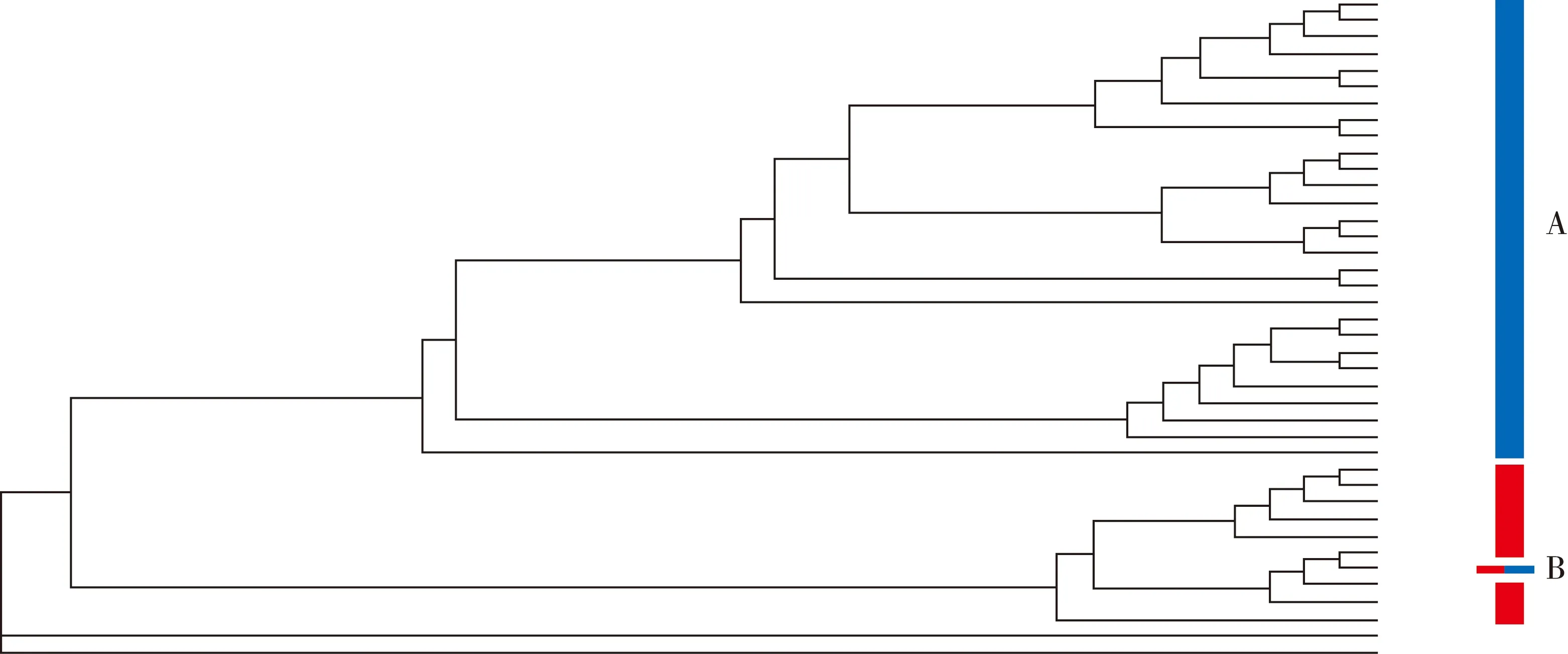
分别对栗斑腹鹀和三道眉草鹀没有性别差异的形态参数、雌性形态参数和雄性形态参数进行聚类分析,三种方法得到了一致的聚类图.其中,依据二者雌性形态参数构建的聚类图(见图2),栗斑腹鹀和三道眉草鹀聚在了一起.
红色为三道眉草鹀个体,蓝色为栗斑腹鹀
2.3 栗斑腹鹀和三道眉草鹀的系统发育树和分化时间
通过4种方法重建的栗斑腹鹀和三道眉草鹀的系统发育树获得了一致的拓扑结构,由其中的 ML进化树(见图3)可见,多数的栗斑腹鹀和三道眉草鹀的单倍型被分为两大进化枝,且具有高的节点支持率(ML:76.2%,MP:100%,NJ:100%,BI:0.9).然而,被聚入到三道眉草鹀进化枝中的单倍型Hap29为两个物种的共享单倍型,包含了4只栗斑腹鹀和21只三道眉草鹀个体.使用PopArt平台制作的栗斑腹鹀和三道眉草鹀单倍型网络图(见图4)显示,两个物种间存在51次碱基改变.
红色为三道眉草鹀个体,蓝色为栗斑腹鹀
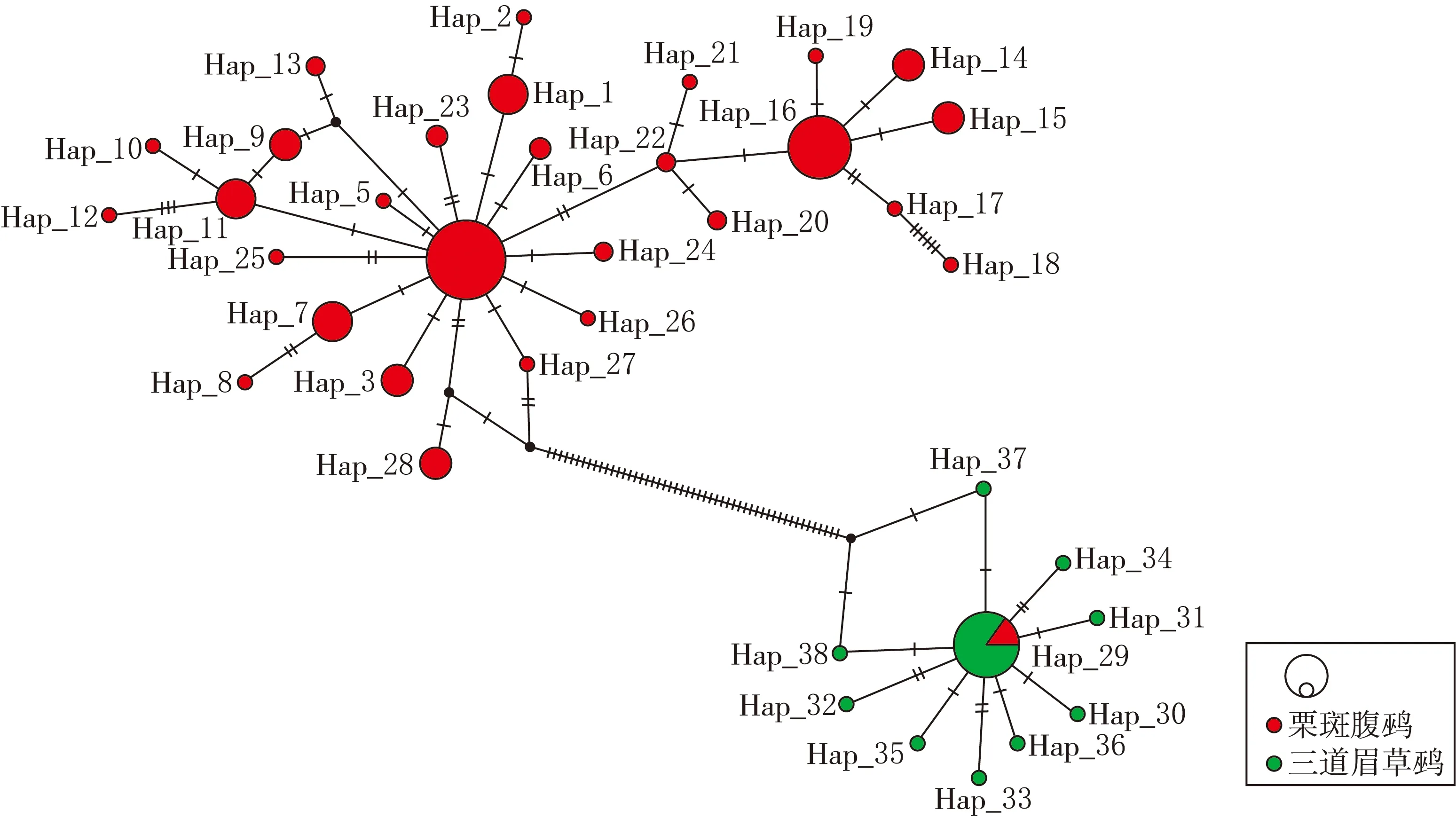
图4 ND2基因序列单倍型网络图
从每巢中选择一只雏鸟,估算栗斑腹鹀和三道眉草鹀的遗传多样性,结果(见表4)表明,栗斑腹鹀的单倍型多样性(h=0.94,标准差=0.01)和核苷酸多样性(π=0.006 2,标准差=0.000 1)均高于三道眉草鹀(h=0.52,标准差=0.11;π=0.000 8,标准差=0.000 2).

表4 栗斑腹鹀和三道眉草鹀线粒体ND2基因遗传多样性
对两个物种的最近共同祖先时间的计算结果表明,栗斑腹鹀和三道眉草鹀的最近共同祖先生活在136万年前(95%置信区间(HPD)=1.05~1.70).实验结果中所有参数的有效样本量(ESS值)都超过了500,其中TMRCA平台的ESS值达到了7 902.从计算结果来看,栗斑腹鹀所有单倍型的最近共同祖先可以追溯到2万年前(95%置信区间=0.15~0.31),三道眉草鹀所有单倍型的最近共同祖先可以追溯到15万年前(95%置信区间=0.08~0.23).
3 讨论
3.1 鸣声分化
鸣声是鸟类重要的通讯手段,包含着丰富的信息,在吸引和稳定配偶关系、保卫领域、育雏等环节发挥着重要作用,具有物种特异性[18].本文对内蒙古扎鲁特旗区域内,栗斑腹鹀和三道眉草鹀两个物种鸣声参数的分析结果表明,二者间除持续时间外,其他声学参数均不具有显著性差异(见表1).已有研究[19]表明,鸣声差异受到很多因素的影响,比如生境类型、地理差异和学习行为等.三道眉草鹀具有学习栗斑腹鹀鸣声的能力[18],通过本文利用鸣声参数构建的聚类图可见,61只个体聚成了2个主要分枝(A和B),但是在A分支中,栗斑腹鹀和三道眉草鹀聚在了一起,聚类分析并未将栗斑腹鹀和三道眉草鹀完全分开.因此,二者间鸣声的相似性可能是由于三道眉草鹀的学习所致.对不同区域内生活的栗斑腹鹀的鸣声特征进行分析后发现,在查干套力皋和嘎亥图地区分布的栗斑腹鹀鸣声参数中,只有时间间隔存在显著差异(见表2),推测可能是由于地理差异导致的不同区域的栗斑腹鹀间出现了鸣声分化.有研究表明,很多鸟类都具有鸣声微地理变异,如:杨晓菁等[20]对武汉地区的白头鹎进行的鸣声研究表明,相隔2.5 km的两个种群就能产生完全不同的鸣唱特征,地理距离越近鸣唱特征越相似.关于栗斑腹鹀不同种群间鸣声特征是否存在微地理变异仍需要进一步研究.
3.2 形态分化
多数鸟类的形态和鸣声一样,具有物种特异性,这在鸟类分类学中具有重要的作用.本文的研究表明,栗斑腹鹀和三道眉草鹀的喙高具有显著差异,三道眉草鹀的喙要比栗斑腹鹀高(见表3),这可能是由于二者食物来源的差异所导致的.Haavie等[21]对西丛鸦(Aphelocomacalifornica)和西丛鸦落基山亚种(A.c.woodhouseii)进行的研究发现,这两个最近分化的物种,由于食物资源的不同,在喙形上表现出明显的不同,西丛鸦有更重的钩状喙,而西丛鸦落基山亚种的喙则较窄、较长,因此认为鸟喙的大小和形状反映了鸟类食物资源的差异[22].此外,栗斑腹鹀在体型上也较三道眉草鹀略小一些.两个物种的雄性个体都较雌性个体略大,这可能是性选择的结果,雌性更偏爱体型较大的雄性[19,23-24].
鸟类的尾,作为飞行特性的一个组成部分,具有辅助飞行的作用,可以很好地提高鸟类在飞行时的稳定性.尾羽这一功能的进化促进了鸟类的多样化,使它们能够更广泛的飞行,从而开发更大范围的生态位[25].本文的研究表明,三道眉草鹀的尾长要长于栗斑腹鹀,同时三道眉草鹀的翅长也要长于栗斑腹鹀(见表3),表明三道眉草鹀可能具有更好的飞行能力和适应性.然而,利用栗斑腹鹀和三道眉草鹀的形态参数构建的聚类图同样未将栗斑腹鹀和三道眉草鹀完全分开(见图2),表明栗斑腹鹀和三道眉草鹀间尚未出现完全的形态分化,二者间仍具有相似的形态特征.
3.3 分子分化
分子进化的中性理论认为,DNA中的大部分突变是中性或者近中性的,并不会改变生物的表型,也不会对生物的适合度产生影响或影响很小.因此,DNA中同义突变的累积会远高于非同义突变,而同义突变一般不会导致表型改变[26].这一结果表明,仅仅依据表型特征对物种分化进行研究是不全面的,可能会忽略掉DNA中存在的大量并未引起表型改变的遗传变异.本研究利用线粒体数据重建了栗斑腹鹀和三道眉草鹀的系统发育树,发现大部分个体都被归入到了正确的分类群中.但Hap29单倍型为栗斑腹鹀和三道眉草鹀共有的单倍型.尽管单倍型网格图显示两个物种在历史上出现了快速分化,二者间存在51次碱基改变[27],但仍不能否定栗斑腹鹀和三道眉草鹀间可能由于较近的分化时间和不完全的表型分化而在同域分布区内发生过杂交渐渗.之前的研究表明,一些物种间由于分化尚不完全,在接触区内常常会发生种间杂交并产生可育后代[7],即便物种分化时间达到1 700万年,也可能诞生可遗传的后代[28].
研究[27]表明,第四纪冰期-间冰期旋回的气候变化对物种形成、分布区演变、种群分化、遗传格局等方面产生了重要影响.许多新物种的形成以及种群分化都发生在这一时期.本文参考Irwin等[17]关于黄鹀和白头鹀的研究,以2%每百万年作为ND2基因的突变率,用BEAST version 1.6.2平台估算了两个物种的最近共同祖先时间,结果表明栗斑腹鹀和三道眉草鹀的最近共同祖先生活在136万年前(95%置信区间=1.05~1.70).这一时期,中国正处于“鄱阳冰期—鄱阳-大姑间冰期—大姑冰期”的转换时期[29],气候的波动和温度的下降可能迫使这两个物种的祖先被隔离在不同的冰期避难所中,并在不同的冰期避难所内发生了独立的进化,进而形成新的物种.
本文的研究表明,栗斑腹鹀和三道眉草鹀的分化时间未超过200万年,为典型的年轻物种[4],这也可以解释二者间在形态特征和鸣声特征上尚未发生完全分化的原因.在内蒙古繁殖分布区内的栗斑腹鹀所有单倍型的最近共同祖先可以追溯到23万年前(95%置信区间=0.15~0.31),三道眉草鹀所有单倍型最近共同祖先可以追溯到15万年前(95%置信区间=0.08~0.23).
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
东坡赤壁诗词(2023年2期)2023-05-30 15:02:14
清华金融评论(2022年4期)2022-04-13 21:33:11
诗歌月刊(2021年8期)2021-08-25 18:54:10
小天使·二年级语数英综合(2021年6期)2021-08-09 10:25:54
国际放射医学核医学杂志(2021年10期)2021-02-28 08:43:46
科学大众(中学)(2019年3期)2019-05-17 10:04:30
安徽师范大学学报(自然科学版)(2018年5期)2018-12-11 03:32:00
汽车观察(2018年10期)2018-11-06 07:05:26
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
