
扬子鳄幼鳄求救鸣声声谱特征及个体签名
2018-12-11 03:32:00赵琳,章平,张方
安徽师范大学学报(自然科学版) 2018年5期
赵 琳, 章 平, 张 方
(安徽师范大学 生命科学学院,安徽 芜湖 241000)
与其他爬行动物相比,鳄类可以发出较为复杂的声音曲目[1,2]。对幼鳄而言,根据所受刺激的不同,可将其声音曲目分为孵化鸣声(hatching call)、联系鸣声(contact call)、求救鸣声(distress call)及威胁和干扰鸣声(threat and disturbance call)[1,3-7],这些鸣声是幼鳄与母鳄及其家族其他成员间建立通讯联系的重要媒介[1]。
在被天敌捕食时,幼鳄会发出求救鸣声[3,5,8],其目的可能有两方面:一是吸引母鳄的注意,以获得特定的关爱和保护[8];二是向同伴发出示警信号,以警示同伴及时采取措施抵御潜在的危险[6,9]。在听觉上,幼鳄不同的声音曲目差异不大[8],但有研究表明,眼镜凯门鳄(Caimancrocodilus)、密河鳄(Alligatormississipiensis)和尼罗鳄(Crocodylusniloticus)的求救鸣声与其发出的其他声音曲目不同[5]。
个体识别是一个动物根据个体与众不同的特征识别另一个动物[10],在动物的交流中具有重要作用[11,12]。多种动物的幼体在独立之前,父母会为其提供资源和保护[13],在此过程中,父母与其幼体使用特定的线索相互识别,并建立起较为精确的识别系统,以避免给无关的幼体照顾[14]。在哺乳类、鸟类和两栖类动物中,声通讯是动物间相互交流和识别的重要方式之一,且具有独特的个体签名音,可能更有利于进行亲缘识别[6,15,16]。
扬子鳄(Alligatorsinensis)母鳄具有保护卵及其幼体的习性[17]。在幼鳄孵化后的几天内,母鳄对幼鳄的鸣声较为敏感,且对幼鳄的求救鸣声具有强烈的行为反应[16,17]。当扬子鳄幼鳄受到天敌袭击时,其发出的求救鸣声具有怎样的声谱特征?幼鳄的求救鸣声是否具有个体差异性?本研究通过模拟“天敌袭击”诱导1日龄的扬子鳄幼鳄发出求救鸣声[9],分析其声谱特征,通过与幼鳄出壳后鸣声[7]对比,初步探明扬子鳄幼鳄求救鸣声的声谱特征及其是否存在个体签名音。
1 材料与方法
1.1 试验地点和试验对象
2016年8月至9月,在安徽省芜湖市籍山扬子鳄繁殖中心录制声音。从7个孵化箱随机选取1日龄扬子鳄幼鳄(出壳后24小时之内的幼鳄),刺激幼鳄并进行录音。
1.2 声音录制和声音分析
用Shure SM11话筒(频率范围50Hz~15kHz)和便携式专业录音机(Sound Devices,Model702,频率范围10Hz~96kHz)录制声音。通过模拟“天敌袭击”,即抓住尾巴并摇晃[9],刺激1日龄幼鳄发出求救鸣声。录音时,话筒距离1日龄幼鳄吻端10~20mm,每只幼鳄录音14~30声,每只1日龄幼鳄录音总时长1分钟左右。录音期间,环境温度约31℃,相对湿度约90%,噪音约42dB SPL。
用SELENA和Batsound Pro vs.3.31(Electronik,Uppsala,Sweden)对求救鸣声的频谱和时长参数进行分析,采样频率设为48kHz。根据SELENA给出的语谱图(spectrograms)和示波图(oscillograms),分析声音的基频和时长参数:起始频率F0init(Hz)、最高频率F0max(Hz)和结束频率F0ter(Hz),及向上调频时长Dup(ms)、向下调频时长Ddown(ms)和总时长Dtot(ms)(图1);根据基频和时长参数计算向上调频坡度Slopeup(Hz/ms)和向下调频坡度Slopedown(Hz/ms)(图1)。两个调频坡度按照以下公式计算:Slopeup=(F0max-F0init)/Dup,Slopedown=(F0ter-F0max)/Ddown。根据Batsound Pro vs.3.31给出的能谱图(power spectrum),分析幼鳄求救鸣声的主频Fdom(Hz)(图2)。
1.3 数据分析
独立样本t-检验,将求救鸣声与出壳后鸣声的相应声音参数进行比较[7],以探明求救鸣声的声谱特征及其与出壳后鸣声的差异;通过Kruskal-Wallis(K-W)检验,判断求救鸣声的各声音参数在个体间是否具有显著差异。去除声音数目不足16声的个体,选择其余每只1日龄幼鳄的前16个求救鸣声,并去除具有显著相关性的部分参数,对其余幼鳄求救鸣声的声音参数做判别分析,判定求救鸣声是否具有个体签名音。
2 结 果
2.1 求救鸣声频谱特征
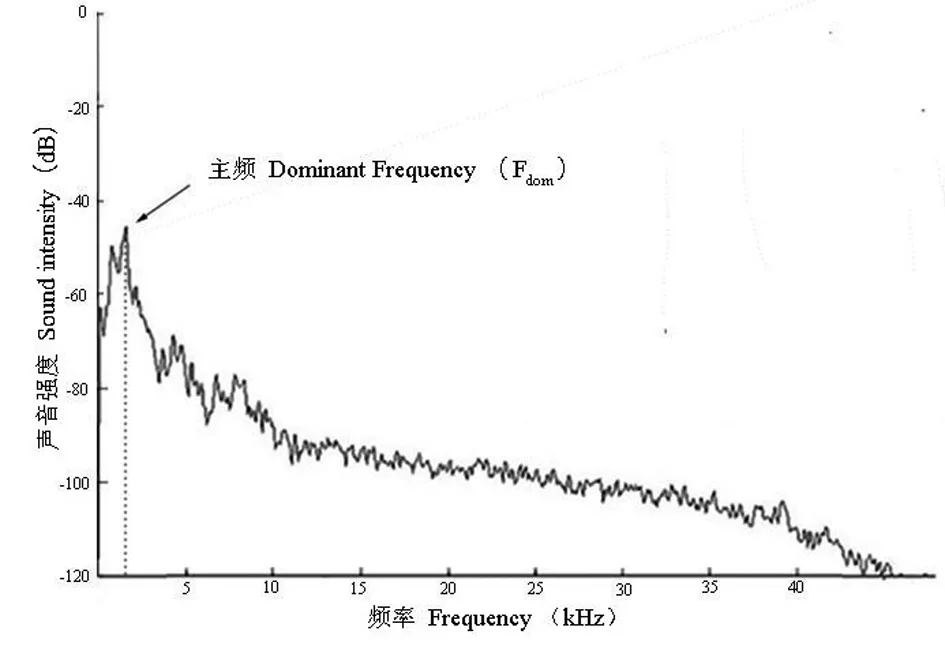
图2 1日龄扬子鳄幼鳄求救鸣声的能谱图Fig.2 Power spectrum of distress calls of one-day old Chinese alligator hatchlings
本研究中,求救鸣声均是1日龄幼鳄在闭口时发出。从声音的语谱图可以看出(图1),幼鳄求救鸣声由多组持续叠加的谐波构成,并伴随强烈的向上调频和向下调频,且绝大多数声音在第3谐波显示了最高能量。
求救鸣声的基频<1000Hz,其中起始频率(467.9±62.8)Hz,最高频率(567.8±69.6)Hz,结束频率(220.2±30.3)Hz;主频>1000Hz(表1)。向上调频时长和向下调频时长占总时长的百分比(Dup/Dtot,Ddown/Dtot)分别为14.3%±3.2%和85.3%±3.5%,向上调频坡度和向下调频坡度分别为(21.5±9.3)Hz/ms和(-11.4±1.9)Hz/ms(表1)。
与出壳后鸣声相比,求救鸣声的起始频率、最高频率、主频及向上调频时长和向下调频时长、向上调频坡度和向下调频坡度均显著偏高(表1,P<0.05),两种声音的结束频率和总时长无显著差异(表1,P>0.05)。
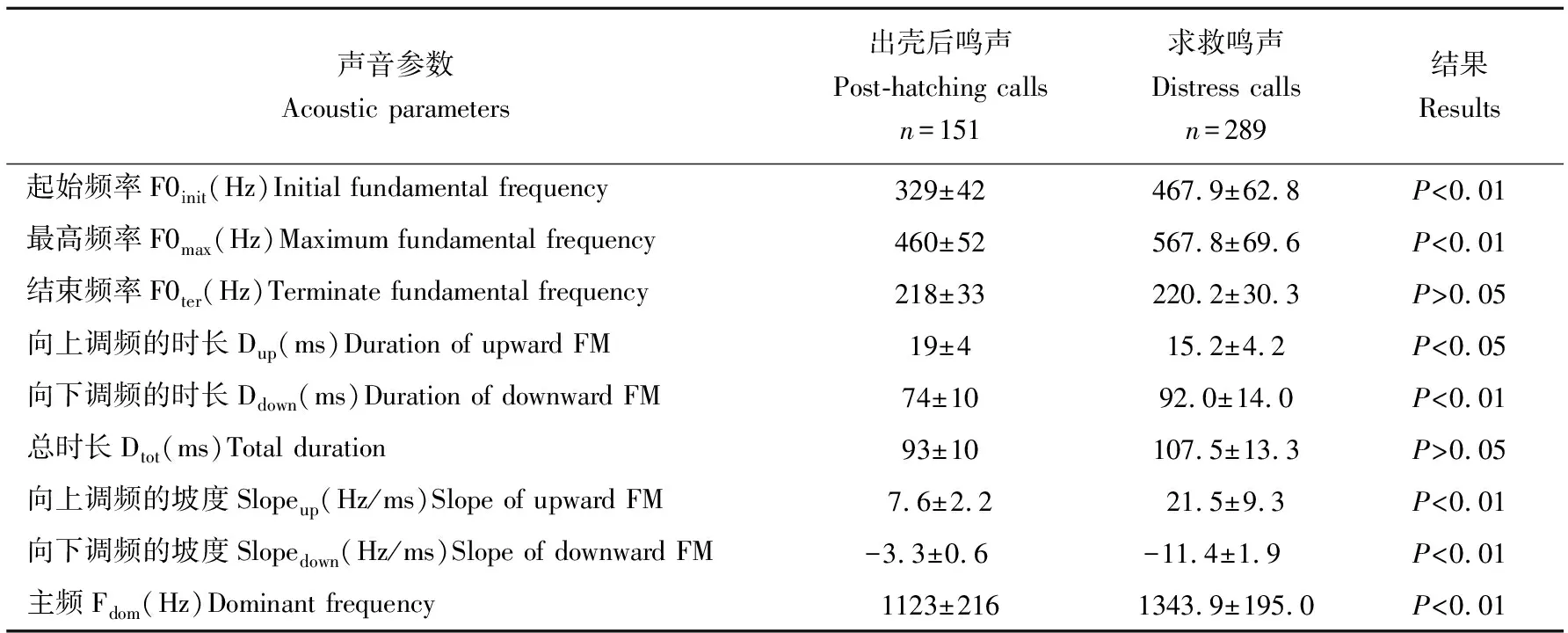
表1 1日龄扬子鳄幼鳄求救鸣声和幼鳄出壳后鸣声的声谱特征的差异
n:鸣声个数;出壳后鸣声:出壳后鸣声的数据引自陈媛媛等[7]
n:calls number;Post-hatching calls:data of post-hatchling calls came from Chen Yuan-yuan et al[7]
2.2 求救鸣声的个体签名
K-W检验结果显示(表2),求救鸣声的起始频率、最高频率、结束频率、主频、向上调频时长、向下调频时长、向上调频坡度及向下调频坡度在个体间均存在显著差异(P<0.01)。

表2 1日龄扬子鳄幼鳄求救鸣声的个体间差异
判别分析结果显示:原始数集中,平均52.6%的求救鸣声被正确分配到相应的幼鳄个体,随机数集中;平均7.9%~11.3%的鸣声被正确分配到相应的幼鳄个体(表3)。配对样本t-检验显示,随机数集平均分配到相应个体的概率与原始数集的相比显著偏低(表3)。
3 讨 论
初孵幼鳄鸣声的基频(包括起始频率、最高频率和结束频率)、主频和时长会随着幼鳄年龄的变化有所不同[6],且幼鳄的年龄越小,声音的频率越高[18],因此本研究选取1日龄的幼鳄为实验对象,有效防止了年龄差异对实验结果产生的影响。
本研究中,K-W检验及判别分析结果表明,1日龄扬子鳄幼鳄求救鸣声具有个体签名音,据此推测在扬子鳄幼鳄与母鳄的交流过程中,母鳄可能会根据求救鸣声识别其幼鳄。求救鸣声具有较短的向上调频时长和较长的向下调频时长,且向上调频较为急剧和向下调频相对缓慢。虽求救鸣声与出壳后鸣声结构相似,但二者声音类别不同。1日龄扬子鳄幼鳄可能通过改变鸣声的起始频率、最高频率和主频以及向上调频时长和向下调频时长,来改变鸣声的类别。在传播过程中,声音的频率越高,其衰减速度越快,传播距离越短[19]。求救鸣声具有较高的起始频率、最高频率和主频,可缩短其传播距离,可能更有利于在巢穴附近的母鳄迅速采取措施给幼鳄以照顾和保护[20]。在野外,近距离声通讯可防止天敌听到,降低幼鳄被捕食的几率,对保证幼鳄的存活率有一定的意义[7]。
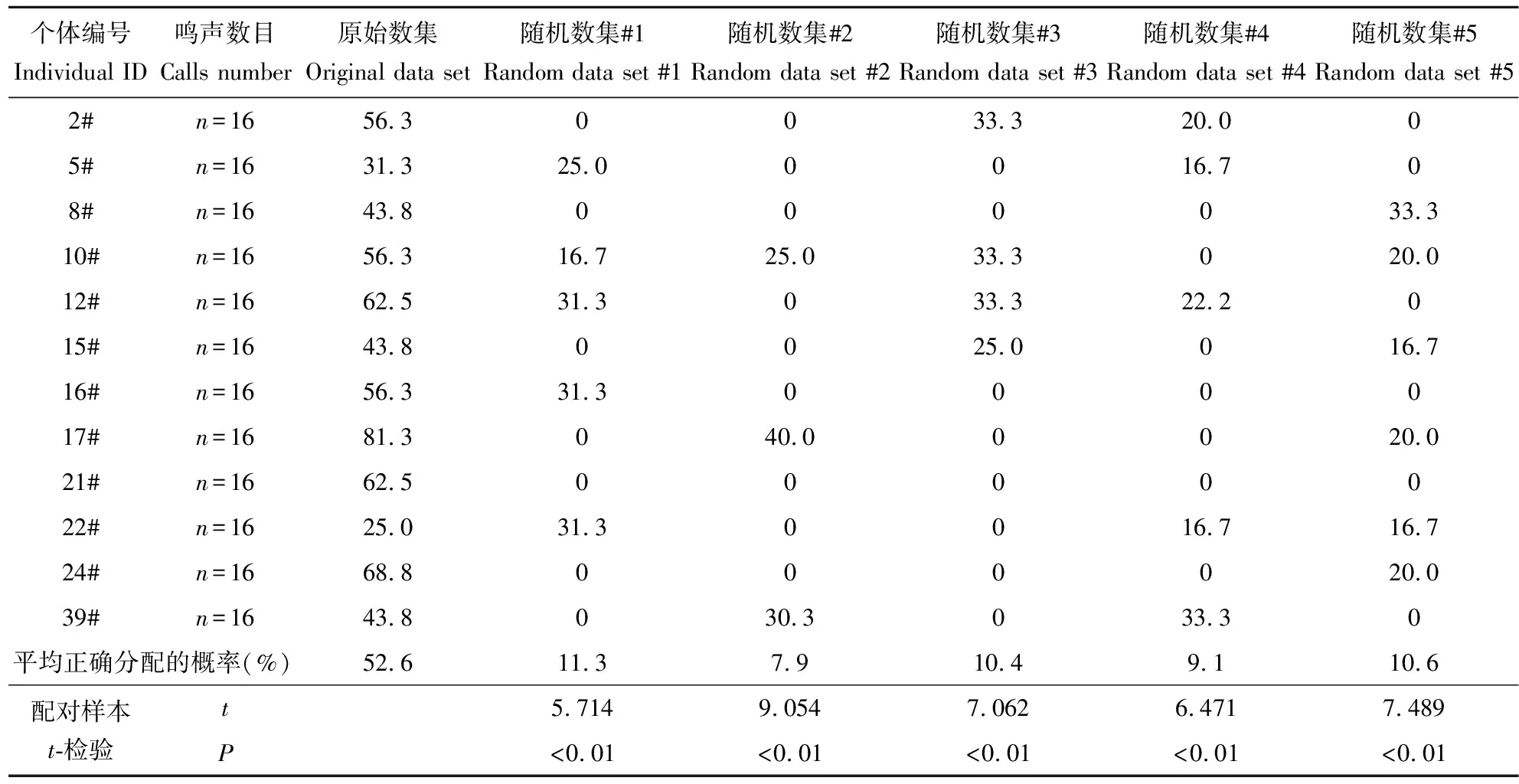
表3 1日龄扬子鳄幼鳄求救鸣声的判别分析结果
随机数集#1~#5:从原始数集中随机选取
Random data set #1~#5:selected from the Original data set randomly
鳄类鸣声具有种间差异[1,18],如尼罗鳄的多数鸣声由多组向下调频的谐波组成[6],眼镜凯门鳄的鸣声均由多组向下调频的谐波组成[2],1日龄扬子鳄幼鳄的求救鸣声均由多组具有向上调频和向下调频的谐波组成。鳄类求救鸣声的声音强度及频率与腭瓣的开合程度有关[4,5],根据鸣叫时腭瓣的开合状态,鳄类求救鸣声可分为呻吟声(moan,腭瓣闭合,频率较低)和尖叫声(screech,腭瓣张开,频率较高)[4]。本研究中,求救鸣声均在腭瓣闭合时发出(即呻吟声),说明扬子鳄幼鳄和眼镜凯门鳄的求救鸣声存在种间差异。另外,由于本试验选取的扬子鳄幼鳄与眼镜凯门鳄年龄不同,也可能会产生这种差异。
猜你喜欢
作文周刊·小学四年级版(2024年4期)2024-03-02 12:40:19
东坡赤壁诗词(2023年2期)2023-05-30 15:02:14
能源工程(2021年5期)2021-11-20 05:50:42
诗歌月刊(2021年8期)2021-08-25 18:54:10
小天使·二年级语数英综合(2021年6期)2021-08-09 10:25:54
小哥白尼(野生动物)(2021年10期)2021-02-12 03:10:14
浙江林业(2018年7期)2018-05-14 10:26:50
西部广播电视(2015年9期)2016-01-18 03:46:07
西部广播电视(2015年9期)2016-01-18 03:46:04
海军航空大学学报(2015年4期)2015-02-27 13:45:51
