
木荷种子园自由授粉家系生长遗传变异及初选
2022-09-26 01:05欧阳天林朱柯帆邱建勋刘武阳邱全生肖德卿王云鹏周志春
中南林业科技大学学报 2022年9期
欧阳天林,朱柯帆,邱建勋,刘武阳,邱全生,肖德卿,王云鹏,周志春
(1.江西省林业科技实验中心,江西 信丰 341600;2.江西省信丰县林木良种繁育中心,江西 信丰 341600;3.中国林业科学研究院亚热带林业研究所 浙江省林木育种技术研究重点实验室,浙江 杭州 311400)
种子园在林木良种生产中发挥着极其重要的作用,然其仍不可避免地存在部分表现不优的基因型,通过剔除表现不良的基因型可优化种子园结构,提升种子园良种的遗传品质。构建优良遗传品质的育种群体是进行多目标性状遗传改良的基础,是保障高世代育种持续推进的必要条件。林木在环境压力选择下形成的适应性特征可通过生长性状表达出来[1]。子代测定是遗传改良的核心内容,也是高世代育种的重要环节。通过子代测定,回溯确定建园优良亲本和前向选择下一世代育种材料,可显著提高遗传增益。马尾松在1代选择群体的基础上通过配合选择的方法选择出2代育种材料,材积的遗传增益达到了21.95%[2]。杉木在初代种子园基础上,经过遗传改良的无性系,材积遗传增益达到了52.80%[3]。综上研究表明,通过对初代种子园的遗传改良,均可达到一定的改良效果。因此,通过初代种子园的子代测定,对种子园经营管理、实现种子园良种的高产稳产具有重要意义[4-6]。
木荷Schima superba为山茶科Theaceae木荷属常绿阔叶大乔木,是我国亚热带常绿阔叶林的主要建群种,是我国南方主要的珍贵阔叶用材、高效生物防火和生态修复造林树种[7-9]。木荷作为珍贵优质阔叶用材树种,育种起步较晚。前期研究发现,木荷天然林优树自由授粉家系的生长性状存在丰富的遗传变异[10]。然而,目前对木荷1代种子园建园亲本的优劣和遗传改良效果仍缺乏研究,这滞缓了木荷遗传改良的推进。鉴于此,本研究选用木荷1代无性系种子园自由授粉家系的7年生测定材料,分析其树高、胸径和材积指数的遗传变异规律及其所受遗传控制,并估算木荷家系的遗传增益。在此基础上,评价种子园建园亲本优劣,同时筛选出优良家系,以期为木荷的遗传改良提供科学指导及为高世代育种提供优良材料。
1 材料和方法
1.1 试验材料和设计
7年生木荷种子园自由授粉家系子代测定林营建于江西省信丰县林木良种繁育中心(114°55′E、25°23′N),试验地属中亚热带季风气候区,年均气温19.9℃,年均降水量为1 568 mm,无霜期为298 d,土壤为山地黄红壤,肥力中等。参试材料为木荷1代无性系种子园自由授粉家系49个和未经遗传改良的当地商品种(对照)1个,共计50个(表1)。

表1 参试家系的产地及数量Table 1 The source and number of the tested S.superba families
2014年春,选用50个木荷种子园自由授粉家系的1年生轻基质容器苗(容器规格4.5 cm×10 cm,苗高25 cm),采用完全随机区组设计营建木荷家系试验林,5次重复,8株单列小区,即每个家系40株,株行距2.0 m×2.0 m。造林前每穴施复合肥0.1 kg,按一般生产经营进行造林和幼林抚育。
1.2 数据调查和处理
2020年12月初,对木荷1代无性系种子园自由授粉家系进行全林测定,调查性状包括树高(H)和胸径(D),以D2×H计算材积指数。采用SAS(v 9.2)软件的GLM和VARCOMP等过程进行方差分析和方差分量估计,进而估算遗传参数[11-12]。
性状分析使用混合线性模型,为:

式中:Yijk为第i个区组第j个家系第k个单株观测值;u为群体平均效应;Bi为第i个区组效应;Fj为第j家系的效应;BFij为第i家系和第j区组的互作效应;Eijk为机误。此外,u和Bi为固定效应,其余为随机效应。
家系遗传力:

式中:n为小区株数调和值;b为区组数;为家系方差分量;为机误;为家系与区组互作的方差分量。
表型变异系数:CVp=σp/X×100%。
式中:Pσ为性状的表型标准差;为遗传方差分量;为性状的群体均值。
式中:G为现实增益;ΔG为遗传增益;Xi为入选家系均值;为商品对照测定值;h2为家系遗传力。
2 结果与分析
2.1 木荷家系生长性状变异
7年生木荷的树高、胸径和材积指数在家系间均存在极显著的遗传差异(P<0.01)(表2)。树高、胸径和材积指数的最大家系分别为最小家系的1.43、1.62和3.83倍,可见材积指数在家系间的差异较大,胸径次之,这为优良家系的选择提供了基础。

表2 木荷家系生长性状方差分析†Table 2 Variance analysis of the growth traits of S.superba family
木荷家系的树高、胸径和材积指数的方差分量占比结果(表3)显示,生长性状的家系效应平均占比为14.05%,其中树高受家系效应影响最大(15.57%),材积指数和胸径受家系影响较小,分别为13.14%和13.45%,说明7年生木荷的树高受遗传效应的影响较大。此外,木荷家系的生长性状除受遗传效应的影响之外,还受遗传与环境互作效应的影响,平均占比为57.94%。
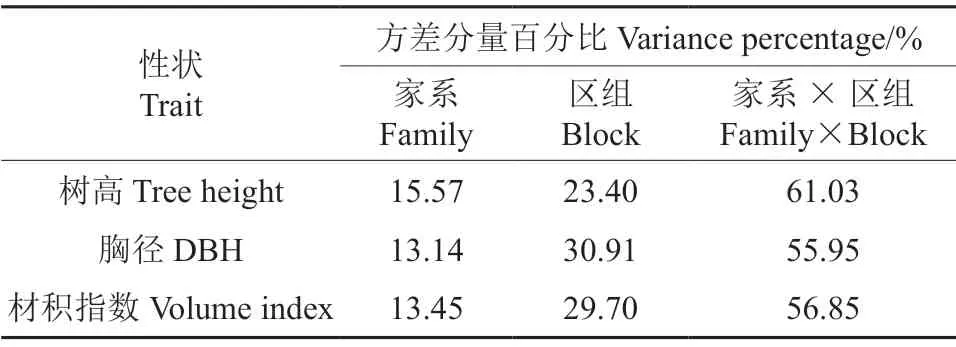
表3 木荷生长性状方差分量百分比Table 3 The variance percentage of the growth traits of S.superba
7年生木荷家系树高、胸径和材积指数的表型变异系数(7.89%、9.64%和27.06%)均高于遗传变异系数(6.28%、9.02%和24.60%)(图1),这有利于木荷速生优良家系的选择。

图1 木荷树高、胸径和材积指数的变异系数Fig.1 Variation coefficients of height, DBH and volume index of S.superba
2.2 木荷家系生长性状的遗传力
7年生木荷树高、胸径和材积指数的家系遗传力分别为0.47、0.42和0.44(图2),均受较强的遗传控制;单株遗传力分别为0.27、0.19和0.23,均受中度的遗传控制。木荷家系遗传控制要强于单株遗传控制,这为木荷优良家系的选择提供了可能。

图2 木荷树高、胸径和材积指数的遗传力Fig.2 Heritability of height, DBH and volume index of S.superba
2.3 木荷优良家系选择及增益估算
为验证木荷1代无性系种子园的遗传改良效果,将木荷种子园自由授粉家系与商品对照进行比较。结果(表4)显示,所有参试家系的生长表现均显著优于对照,树高、胸径和材积指数的群体均值分别高出对照的23.29%、27.11%和110.88%,说明木荷1代种子园遗传改良效果显著。鉴于木荷1代种子园具有较为显著的改良成果,同时本研究中树高和胸径均受较高的遗传控制,且变异系数处于较高水平。因此,采用独立挑选法,以木荷家系树高和胸径均高于对照家系均值的25%为标准筛选出16个优良家系,如YP35、YP7和GT2,入选率为32.65%。入选的16个优良家系树高、胸径和材积指数的平均现实增益分别为34.31%、38.57%和170.02%,平均遗传增益分别为16.12%、16.20%和74.81%。

表4 入选的16个7年生木荷优良家系生长表现及增益Table 4 Growth performance and gain of 16 7-year-old S.superba families
3 讨论与结论
3.1 讨 论
林木的遗传和变异研究是开展遗传改良的前提,掌握遗传变异规律是制定育种策略的基础[13-14]。木荷作为珍贵阔叶用材树种,育种起步较晚,了解其遗传变异规律,选择优良亲本对建园亲本的选择和种子园去劣疏伐等有重要意义。本研究中7年生木荷树高、胸径和材积指数各家系间均存在极显著的差异,木荷1代种子园家系的生长表现显著优于当地商品种,这表明进行家系选择可以取得较大的遗传增益。木荷是一个速生期较长的阔叶树种,幼龄期便对光照、养分和生长空间等具有较强的竞争能力,早期生长较快、表现较好的家系在后期生长中不会发生早衰现象,早期生长处于劣势的家系后期很难超越优势家系[9],表明对木荷家系进行早期选择或淘汰是可行的。研究中还发现木荷家系的树高、胸径和材积指数存在极显著的区组×家系间互作差异,说明生长性状不仅受遗传的影响,还受立地环境与遗传互作的影响。因此,在生产上应针对立地环境选用适生的品种,以达到木荷人工林的高效培育。
精确估算遗传参数对于预期增益和科学制定育种策略具有指导意义[15-16]。王云鹏等[9]发现3年生和10年生的木荷胸径所受遗传控制要强于树高,5年生时则是树高较强。潘艳艳等[17]发现8年生和11年生日本落叶松的树高所受遗传控制强于胸径。本研究发现7年生木荷树高所受遗传控制也强于胸径。林木性状遗传力估算值不仅因性状而有差异,而且与参试材料的数量及试验材料对环境的适应程度有关,林龄、所处生境和试验设计等条件的不同也会导致遗传力的差异。性状受较强的遗传控制有利于获得最大化的遗传增益效果[18-19],木荷树高、胸径和材积指数受较强的家系遗传控制,树高所受到的遗传控制均略高于胸径和材积指数。这既表明木荷通过家系选择可获得较高的遗传增益,也表明在一定的选择强度下,选用树高作为选择性状更有望获得较高的遗传增益。本研究还发现木荷各性状的单株遗传力估算值相对较低,可能原因是7年生木荷还处于幼龄期,单株间生长性状的基因型差异还未完全表达。这也表明通过种子园去劣疏伐,去除表现较差的无性系,有利于提高种子园的整体改良效果。
变异系数大小可反映群体的变异程度,是衡量性状遗传变异潜力的有效指标[20-21]。本研究中,木荷家系树高、胸径和材积指数的遗传变异系数低于表型变异系数,意味着因环境的影响,导致木荷家系性状的表型变异大于其相应的遗传变异。胸径的表型和遗传变异均高于树高,这说明胸径较树高具有更大的改良空间。胸径的变异系数均要高于树高和材积指数,这可能是因为纯林中木荷家系为竞争更多的光照和生长空间优先选择了分配树高生长的策略,从而使得个体间胸径产生较大差异,树高差异较小[22],这也反映出木荷个体对同质资源的一种普遍竞争规律,即竞争木树高可促进对象木树高的生长。据此规律,在生产上可尝试采用目标树与竞争木相结合的方式来促进林木生长,从而实现最大化的林分生长力。
遗传增益是反映林木遗传改良效果的直接指标[23]。通过对木荷1代无性系种子园自由授粉家系进行增益估算,其结果表明木荷种子园改良具有显著效果。木荷种子园自由授粉家系生长表现较好,具有较高的遗传品质,这进一步证明利用木荷种子园良种造林可大大提高林分生产力。这也意味着应根据子代测定结果,伐除木荷种子园中生长表现不良、遗传品质差和低产型无性系,以优化种子园结构。同时,也表明通过对测定家系进行前向选择,可为后续世代提供优良亲本,从而达到较高增益的目标。选择家系时若以单独的生长性状作为标准,其结果随林龄的增长往往会出现变化,而兼顾不同的性状优选出的高生产力家系更加稳定[24-25]。对于用材树种木荷而言,树高和胸径是其主要的经济性状,可反映其生长的优劣,且本研究中木荷树高和胸径均受较强的遗传控制,变异系数处于较高水平。鉴于此,本研究以高于商品对照均值25%为选择标准,初选出16个优良家系。初选的木荷家系树高、胸径和材积指数均值均明显高于商品对照,材积指数的现实增益达170.02%,遗传增益达74.81%,处于较高水平,从16个优良家系中选择出的优良单株可作为下世代的育种亲本。
现阶段仅利用7年生木荷家系测定材料来研究树高、胸径和材积指数的遗传变异规律及所受遗传控制和验证木荷1代种子园遗传改良效果,初选出一批生长优良家系。然而,由于木荷生长周期长,对立地较敏感,今后还需结合多点联合试验进行多年观测,进一步探究木荷家系生长性状的遗传结构及遗传稳定性,并筛选高配合力的亲本组合,以便更加科学地指导木荷育种和营林生产实践。
3.2 结 论
7年生木荷1代无性系种子园自由授粉家系间存在丰富的遗传差异,有较高的选择潜力,树高、胸径和材积指数受中等至较强的遗传控制,均受较强的家系遗传控制,其生长均显著优于商品对照,遗传改良效果显著,初选出的YP35、YP7和GT2等共16个木荷优良家系,材积指数的现实增益达170.02%,遗传增益达74.81%,处于较高水平,可从中筛选优良个体作为下一代育种亲本。
猜你喜欢
林业科学(2022年5期)2022-08-20
散文诗世界(2022年5期)2022-05-23
辽宁林业科技(2021年3期)2021-07-08
南方林业科学(2021年1期)2021-03-16
山西水土保持科技(2020年2期)2020-12-22
农民致富之友(2017年4期)2017-04-10
现代农业科技(2017年4期)2017-04-10
绿色科技(2017年1期)2017-03-01
浙江林业(2015年4期)2015-12-21
湖北农业科学(2014年3期)2014-07-21
