
松材线虫侵染不同抗性松树后寄生适应性与致病力的变化
2022-08-27 00:49:36应晨希胡龙娇刘红斌吴小芹
西南林业大学学报 2022年4期
应晨希 胡龙娇 刘红斌 吴小芹
(1. 南京林业大学南方现代林业协同中心,林学院,江苏 南京 210037;2. 江苏省有害生物入侵预防与控制重点实验室,江苏 南京 210037)
松材线虫(Bursaphelenchus xylophilus)是松树萎蔫病(亦称松材线虫病)的病原[1]。自1982年我国南京首次报道松材线虫病发生以来[2],该病在38年的时间里,已先后在我国18省666个县级行政区发生为害,造成严重的经济损失和生态破坏,松材线虫已成为我国最具危险性的林业有害生物之一。近年来,松材线虫病在我国的传播危害呈现一些新特点:发生区突破了传统理论的北部适生界线;传播媒介虫种增加;有研究表明,目前除松墨天牛外林间的其他天牛科(Cerambycidae)昆虫以及象甲科(Curculionidae)、吉丁科(Buprestidae)、叩甲科 (Elateridae)和小蠹科 (Scolytidae)等昆虫也可携带松材线虫[3−7];感病树种由过去的黑松(Pinus thunbergii)、赤松(P. densiflora)和马尾松(P. massoniana)等,扩展到黄山松 (P. taiwanensis)、云南松 (P. yunnanensis)、油 松(P. tabuliformis)、华山松(P. armandii)、红松(P. koraiensis)和落叶松(Larixspp.)等[8−12]。松材线虫病在我国远距离传播的主要方式是人为传播含松材线虫和媒介昆虫的病木及其制品。
研究表明,在自然界中病原物和昆虫均存在寄主转换现象,它们的寄生适应性将直接影响其种群的定殖、扩张继而影响病虫害的发生[13]。有学者对B型烟粉虱(Bemisia tabaci)在寄主转换时的适应性研究发现,烟粉虱的寄主适应度变化符合一般的寄主转移规律,即嗜性相对较强的过渡寄主可以刺激烟粉虱提高寄主适应性[14−16]。而棉蚜(Aphis gossypii)从棉花种群被转到甜瓜(Cucumis melo)上时,其寿命比转到原寄主棉花的成蚜寿命明显降低[17]。目前松材线虫的寄主范围不断扩大,其由媒介昆虫携带从病死木传播到健康木、从病害发生区传播到新地区的过程中都有可能在不同抗性松树之间发生转换,但目前松材线虫在传播过程中寄主发生变化时的寄生适应性和致病力变化状况尚不清楚。
马尾松作为我国本土重要的松属树种,多年来其抗性种源和抗病单株选择在江苏和安徽等省陆续被进行研究,目前已筛选出了一批高抗马尾松,在安徽初步建立了抗松材线虫病候选个体保存园[18]。目前对于松材线虫病的寄主范围和寄主抗性等问题已有相关研究[19−21],而松材线虫在寄主体内的种群增殖力是关乎松树是否发病的关键因子,但关于松材线虫在不同抗性松树体内的种群繁殖参数等与寄生适应性相关因素的研究还少见报道。同时,在松材线虫相关致病机理的研究中,对于纤维素酶在松材线虫病发生中的作用研究报道较多[22−23],但对于在不同抗性寄主体内松材线虫分泌的纤维素酶活性变化状况不甚明确。为此,本研究选取不同致病力的松材线虫通过多次接种不同抗性的寄主松树,对松材线虫侵染松树后的种群繁殖力、致病力变化及其分泌纤维素酶的能力进行研究,以期阐明松材线虫种群的寄生适应性及其在寄主变换时种群繁殖力和致病力的变化规律,为进一步揭示松材线虫种群的入侵定殖及流行为害机制提供参考依据。
1 材料与方法
1.1 供试材料及其培养
供试松材线虫为经前期致病力测定表现为强毒的虫株ZJ和弱毒的虫株YW4,分别分离自浙江富阳马尾松和云南德宏思茅松(P.kesiyavar.langbianensis)[24],保存于江苏省有害生物入侵预防与控制重点实验室。松材线虫生长适温为 25 ℃[19],通过马铃薯葡萄糖琼脂(PDA)平板在 25 ℃ 无光照条件下培养灰葡萄孢(Botrytis cinerea)4~5 d,菌丝长满 PDA 后接入供试松材线虫,相同的条件下培养 5~6 d,待灰葡萄孢菌丝被吃光后采用贝尔曼漏斗法收集松材线虫[25]。将收集的线虫悬浮液 4000 r/min,离心 3 min,弃去上清;再用双蒸水(ddH2O)清洗 3 次,4000 r/min 离心,每次 3 min,备用。
供试植物为3~4年生生长状况基本一致的健康黑松幼树、火炬松(P. taeda)幼树和高抗马尾松幼树(由江苏省林科院抗性马尾松种源筛选获得,种源编号为GD5),培养于南京林业大学温室,培养温度控制在28~32 ℃,相对湿度控制在65%~75%。
1.2 松材线虫接种松树及致病力测定
将2种不同毒力的松材线虫制备成虫悬液(3000 条/mL),以皮接法对3种松幼树进行人工接种,用无菌刀片在近地约10 cm的茎干处割开树皮深达木质部,用镊子取无菌棉球塞入,每株接1 mL的虫悬液,以接种1 mL ddH2O的松树作为对照,封口膜做成漏斗状将切口及棉球包裹住,保证虫液不渗漏。每个处理设置6个重复。根据针叶颜色变化程度,将松树的感病程度划分为以下5个等级:0级:所有针叶均健康呈绿色;Ⅰ级:小部分针叶开始变黄;Ⅱ级:一半左右的针叶变黄或萎蔫;Ⅲ级:大部分针叶褐变萎蔫;Ⅳ级:所有针叶失绿枯萎。在接种后的第15天、30 天和45 天统计松苗的发病率和感病指数,发病率和感病指数按如下公式进行计算[26]:

当任一处理中松幼树全部死亡或感病指数不变时,将所有供试松幼树按不同处理分别剪成细条收集,通过贝尔曼漏斗法分离线虫并洗净,接种至灰葡萄孢培养基进行扩繁再收集,方法同1.1所述。将收集到的线虫按照上述方法再次接种于健康的松幼树并测定相关指标。重复该过程即为第3次接种。
1.3 松材线虫寄生适应性与雌雄性比的检测
采用Braach[27]提出的相关寄生适应性(RHS)系数分析不同毒力松材线虫在3种松树中的定殖状况。
2种不同毒力松材线虫(强毒虫株ZJ和弱毒虫株YW4)与3种不同抗性松幼树(抗性马尾松、火炬松、黑松)共组成6组接种处理,接种后获得的线虫按上述顺序分别命名为ZJ−Ma、ZJ−Huo、ZJ−Hei和YW4−Ma、YW4−Huo、YW4−Hei。在上述的3次接种试验中,当任一处理中松幼树全部死亡或感病指数不变时,通过贝尔曼漏斗法分离松苗中的线虫,并在蔡司显微镜Axio Lab A1下统计每次接种试验中的松材线虫数量(区分死亡松幼树中的线虫数量)和雌雄虫性比(记录接种前线虫性比)。根据RHS系数公式计算3次接种试验中各处理的RHS系数:

1.4 松材线虫分泌的纤维素酶活性测定
1.4.1 松材线虫胞外纤维素酶的提取
提取从不同抗性松幼树上分离得到的线虫分泌的纤维素酶,参考Kojima等[22]的方法稍作修改,将线虫收集后调整浓度为5×104/mL,于20 ℃下不时震荡摇动, 48 h后收集上清液,置于4 ℃冰箱保存备用。
1.4.2 葡萄糖标准曲线的绘制
取DNS 10.0 g,加400 mL去离子水使之悬浮,边混合边慢慢加入150 mL氢氧化钠溶液(每300 mL去离子水中含32.0 g氢氧化钠)。将该悬浮液置于50 ℃水浴中温热至澄清,不断混合下缓缓加入300 g酒石酸甲钠至溶液中,用去离子水稀释至1000 mL,贮存于棕色瓶中。
取0、0.2、0.4、0.6、0.8、1.0 mL的葡萄糖溶液分别加入0、1、2、3、4、5号管中,每管用去离子水补足体积为2.0 mL,然后加DNS溶液,沸水浴10 min,补足体积为14 mL,以0号管为空白对照,采用紫外分光光度计(CARY100,Agilent)测定490 nm吸光度,在Excel中求得标准曲线方程及R2值。
1.4.3 纤维素酶活性的测定
参考王琳等[28]的DNS法和公式进行测定。反应体系为1 mL 含1%羧甲基纤维素钠(CMC,国药集团,上海)的醋酸钠缓冲液和1 mL酶液,经50 ℃反应30 min,加2 mL DNS溶液,置沸水浴15 min,冷却后加入去离子水使得总体积为14 mL,使用紫外分光光度计测定490 nm吸光度。以每分钟每毫升酶液产生1微克葡萄糖的量作为1个活性单位,将测的葡萄糖含量值(C)代入公式E=C×1000/30×1,求得纤维素酶活性值(E)。
1.5 接种后不同松材线虫种群致病力的比较测定
选取2年生健康且生长状况基本一致的黑松幼树为致病力测定的接种对象,对经过3次循环接种不同抗性松幼树上分离的松材线虫进行致病力统一测定比较。接种量为3000条/株,各处理分别包含8个重复,接种后每5 d观察并记录松树发病情况,直至供试松幼树全部死亡或感病指数不再变化。以持续在灰葡萄孢上继代培养的强毒和弱毒虫株(即出发种群,分别命名为ZJ−CK和YW4−CK)接种黑松幼树作为对照。接种方法和指标测定同1.2所述。
1.6 数据分析
使用Excel计算重复的平均值和标准差作为统计结果,并使用SPSS Statistics 17.0软件(IBM中国公司,北京),采用Duncan多重比较法检验各处理的差异显著性,显著性检验水平为P<0.05。
2 结果与分析
2.1 松材线虫接种不同抗性松幼树后致病力的变化
将松材线虫强毒虫株ZJ和弱毒虫株YW4分别接种不同抗性松幼树(高抗马尾松、火炬松及黑松),通过接种—分离培养—再接种的方式,持续进行3轮接种试验,比较3次试验中不同松树的发病率和感病指数的变化。3次循坏接种强弱毒松材线虫的抗性马尾松幼树的发病率和感病指数变化见表1。在第1轮及第2轮接种中,松材线虫弱毒虫株YW4对抗性马尾松幼树未表现出致病性,即在持续观察的45 d中未出现发病症状;接种强毒虫株ZJ后抗性马尾松幼树的发病率也仅有16.7%,感病指数仅为8.3。在第3轮接种中,各松材线虫虫株对抗性马尾松幼树的致病力均有较大程度的提高,其中接种弱毒虫株YW4的抗性马尾松的发病率为50%,感病指数为16.7,而接种强毒虫株ZJ的抗性马尾松幼树发病率为50%,感病指数达到37.5。抗性马尾松幼树对松材线虫的抗性随着松材线虫接种次数的增加呈现明显下降趋势。这些结果表明无论是强毒还是弱毒松材线虫,通过反复侵染抗性松树幼树其致病力均得到了明显增强。

表1 抗性马尾松幼树循环接种松材线虫强毒和弱毒虫株的发病情况Table 1 The incidence of resistant P. massoniana seedlings after cyclic inoculation with high virulent and low virulent strains of B. xylophilus
松材线虫循环接种火炬松幼树后致病力的变化趋势见表2。在3次接种中,强毒虫株ZJ和弱毒虫株YW4对火炬松幼树的致病力随着接种次数的增加而增强。接种强毒虫株ZJ的火炬松幼树的发病率从第1次接种的66.7%增长至第2次接种的83.3%,最后增长至第3次接种的100%,感病指数由37.5增长至66.7,最后增长至100;接种弱毒虫株YW4的火炬松幼树的发病率从16.7%增长至33.3%,最后增长至66.7%,感病指数由8.30增长至20.1,最后增长至52.08。该结果表明,随着接种次数的增加火炬松幼树也对松材线虫的抗性呈明显下降趋势。

表2 火炬松幼树循环接种松材线虫强毒和弱毒虫株的发病情况Table 2 The incidence of P. taeda seedlings after cyclic inoculation with high virulent and low virulent strains of B. xylophilus
松材线虫循环接种黑松幼树后致病力的变化趋势见表3。对易感寄主黑松,松材线虫强毒虫株ZJ在第1轮测定中已表现出较高致病力,黑松幼树发病率为75%,感病指数为66.7,在第3轮测定中感病指数上升为100。接种弱毒虫株YW4的黑松幼树的发病率由第1次测定的33.3%上升为第3次测定的83.3%,感病指数由20.1上升为81.25,YW4致病力提高尤为显著。
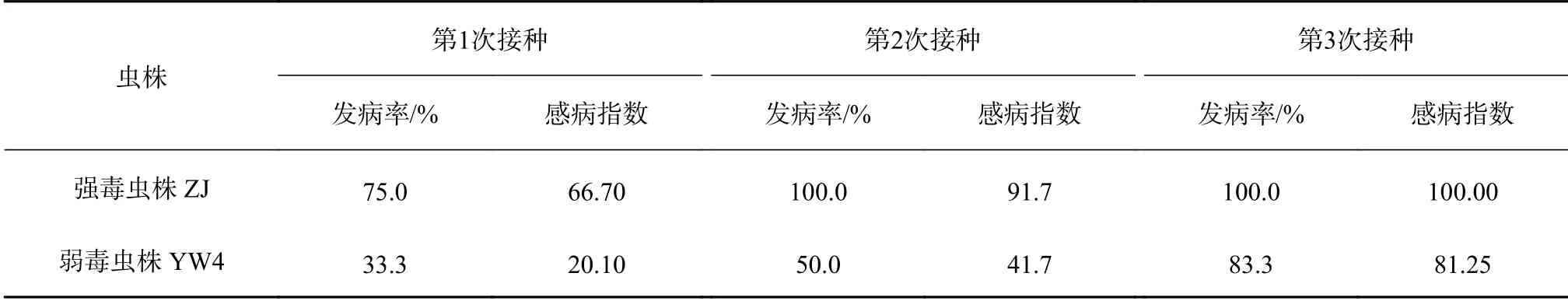
表3 黑松幼树循环接种松材线虫强毒和弱毒虫株的发病情况Table 3 The incidence of P. thunbergii seedlings after cyclic inoculation with high virulent and low virulent strains of B. xylophilus
综合比较不同毒力松材线虫循环接种3种不同抗性松幼树的结果可知,寄主松树无论接种的是强毒虫株还是弱毒虫株,其感病程度高低的顺序依次为黑松种群>火炬松种群>抗性马尾松种群,且经过循环接种3次后测定的发病率和感病指数均显示出较大程度的提升。表明无论强毒或弱毒的松材线虫经多次侵染寄主松树后,其致病力会明显增强。
2.2 松材线虫接种不同抗性幼树后种群寄生适应性的变化
对松材线虫侵染不同松幼树1~3轮后各个种群的寄生适应性RHS系数测定结果见表4。在第1轮的接种试验中,从抗性马尾松幼树中仅分离得到极少量强毒虫株ZJ和弱毒虫株YW4,且接种YW4的抗性马尾松幼树死亡率为0,因此来自抗性马尾松幼树的ZJ种群(ZJ−Ma)RHS系数为0.72,而YW4种群(YW4−Ma)的RHS系数为0。RHS系数最高的为来自易感寄主黑松幼树的ZJ种群(ZJ−Hei),RHS值为9.6;弱毒虫株YW4侵染不同松幼树后各种群的RHS系数均较低,其中最高值为4.5,是来自黑松幼树的YW4种群(YW4−Hei)。

表4 循环侵染不同抗性松幼树过程中松材线虫种群RHS系数Table 4 The RHS coefficient of B. xylophilus populations during cyclic infection with different resistant pine seedlings
在第3轮测定中,强毒虫株ZJ和弱毒虫株YW4在抗性马尾松幼树中的RHS系数均有上升,其中ZJ−Ma种群的RHS系数达到10.5,已高于第1次测定中接种黑松幼树的ZJ−Hei种群的最高值;同时,弱毒虫株YW4−Ma种群的RHS系数达到3.8,已明显高于前2次测定中的YW4−Ma(0)和YW4−Huo的种群(0.4),说明YW4−Ma种群经多次接种高抗性寄主后寄生适应性明显增强。在第3次测定中,松材线虫RHS系数最高仍为接种黑松幼树的ZJ−Hei种群,达到21.6,约达到第1次测定值的2倍,而来自黑松幼树的弱毒YW4−Hei种群的RHS系数为14.1,约达到第1次试验中测定值的3倍。
2.3 松材线虫接种不同抗性幼树后种群性比的变化
松材线虫强毒虫株ZJ和弱毒虫株YW4在每次接种前的雌雄性比均为2.2~2.4,雌虫数量较多,有利于繁殖。将线虫循环接种于不同抗性松树1~3次后,统计分离自寄主体内的松材线虫种群的雌雄性比,结果如图1。在第1轮测定结果中,雌雄性比小于1的种群有3个,分别为ZJ−Ma种群、YW4−Ma种群和YW4−Huo种群;性比为1的种群有1个,为ZJ−Huo种群;雌雄性比大于1的种群有2个,是ZJ−Hei种群和YW4−Hei种群。说明松材线虫在第1次侵染抗性马尾松幼树中雄成虫数量大于雌成虫,而在易感寄主黑松幼树中强毒和弱毒虫株的雌成虫明显增多,这暗示,松材线虫在易感松树中适应性和繁殖力较强从而导致松树发病快,发病程度重。
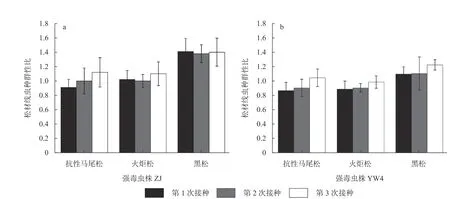
图1 循环侵染不同抗性松幼树过程中松材线虫种群性比的变化Fig. 1 Variation of population sex ratio of B. xylophilus during cyclic infection with different resistant pine seedlings
在第3轮侵染寄主测定中,松材线虫种群的雌雄性比有一定程度的上升,除了YW4−Huo种群为0.98小于1之外,其余5个松材线虫种群的雌雄性比均高于1,即循环侵染寄主后松材线虫多数种群中雌虫所占比例增加,进一步表明循环侵染可以提高松材线虫种群的寄生适应性。ZJ−Hei种群的雌雄性比在3次测定中结果基本保持一致,可见易感黑松幼树是其适宜的寄主,在该寄主环境下松材线虫种群扩繁顺利。这也说明,当松材线虫种群在寄主体内适应性较好时,种群性比趋于稳定。
2.4 松材线虫接种不同抗性幼树后纤维素酶活性的比较
为测定不同抗性松幼树上松材线虫纤维素酶活性,首先进行葡萄糖标准曲线绘制,经计算所得标准曲线方程计算公式为:y=1.5213x+0.0319。相关系数R2=0.9984。之后对来自不同抗性松幼树上松材线虫分泌的纤维素酶活性测定的结果见图2。
比较强弱毒松材线虫多次接种不同抗性松幼树后其分泌的纤维素酶的活力,结果发现强毒虫株群体的纤维素酶活性依次为ZJ−Ma >ZJ−Hei >ZJ−Huo,均高于弱毒虫株;而弱毒虫株YW4的纤维素酶活性依次为YW4−Hei> YW4−Ma>YW4−Huo,但第3次接种后变化为YW4−Ma> YW4−Hei> YW4−Huo。循环侵染不同抗性寄主不同程度地增加了松材线虫种群的纤维素酶活力,其中松材线虫ZJ−Ma和YW4−Ma种群的纤维素酶活力均出现显著上升,ZJ−Huo、YW4−Huo、ZJ−Hei、YW4−Hei种群的纤维素酶活力上升差异不显著(图2)。由此可见,循环侵染高抗性寄主对松材线虫种群纤维素酶活力的增强效应较大。
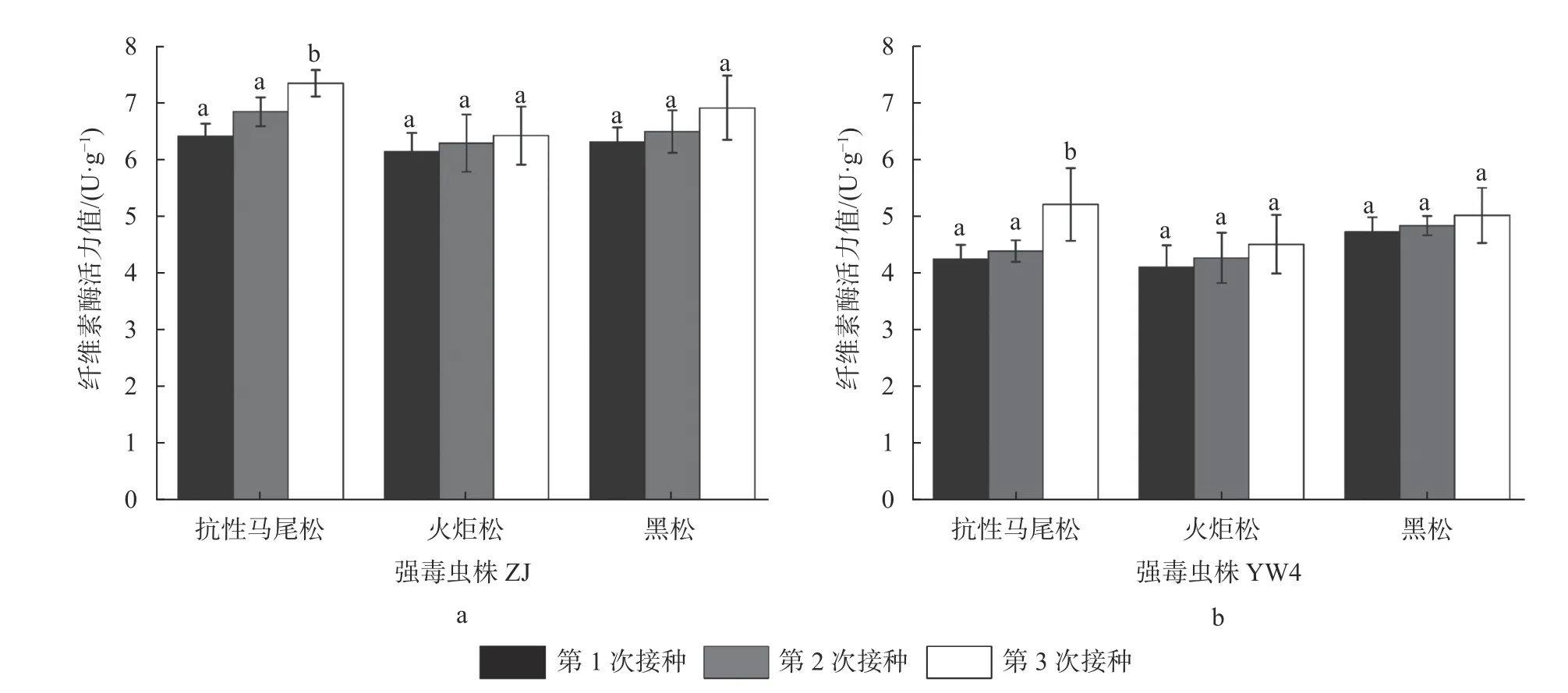
图2 循环侵染不同抗性松幼树过程中松材线虫纤维素酶活性的比较Fig. 2 Comparison of the cellulose activity of B. xylophilus during cyclic infection with different resistant pine seedlings
2.5 循环侵染不同抗性松幼树后松材线虫种群致病力的比较
将经不同抗性松幼树体内增殖后的松材线虫6个种群与对照(出发种群ZJ−CK和YW4−CK)在2年生黑松幼树上进行致病力的比较测定(表5)。结果表明,作为对照的松材线虫强、弱毒虫株出现第1株病死植株的时间分别为第21天和第33天,而在经循环多次接种不同抗性松幼树后的不同种群中,出现第1株病死植株的时间均不同程度的缩短,其中发病最快的为来自经抗性松幼树接种的ZJ−Ma种群和ZJ−Huo种群,接种松幼树后第15天即出现病死植株;发病最慢的为YW4−Huo种群和YW4−Hei种群,接种黑松幼树后第21天出现病死植株,但也比原始出发虫株提前12 d发病。同时,接种ZJ−Ma种群的松幼树全部死亡的历期最短,为21 d,其发病率为100%,感病指数为100,可见该种群致病力最强。接种YW4−Hei种群的发病松幼树死亡历期较长为36 d,但这一数值也低于接种原虫株YW4−CK(45 d,松幼树未全部死亡,发病率和感病指数不再变化)。在发病率和感病指数方面,弱毒虫株YW4接种各个不同松幼树种群的变化数值较为明显,其中经接种抗性松幼树的YW4−Ma和YW4−Huo种群的发病率为87.5%,已达到强毒虫株的水平。接种易感松幼树的YW4−Hei种群的发病率为75%,感病指数为77.5,也成为中等致病力的虫株。这些结果表明松材线虫强、弱毒虫株均可以通过多次侵染松幼树的方式提高其致病力,且循环侵染高抗松幼树后,线虫致病力提高更为显著。
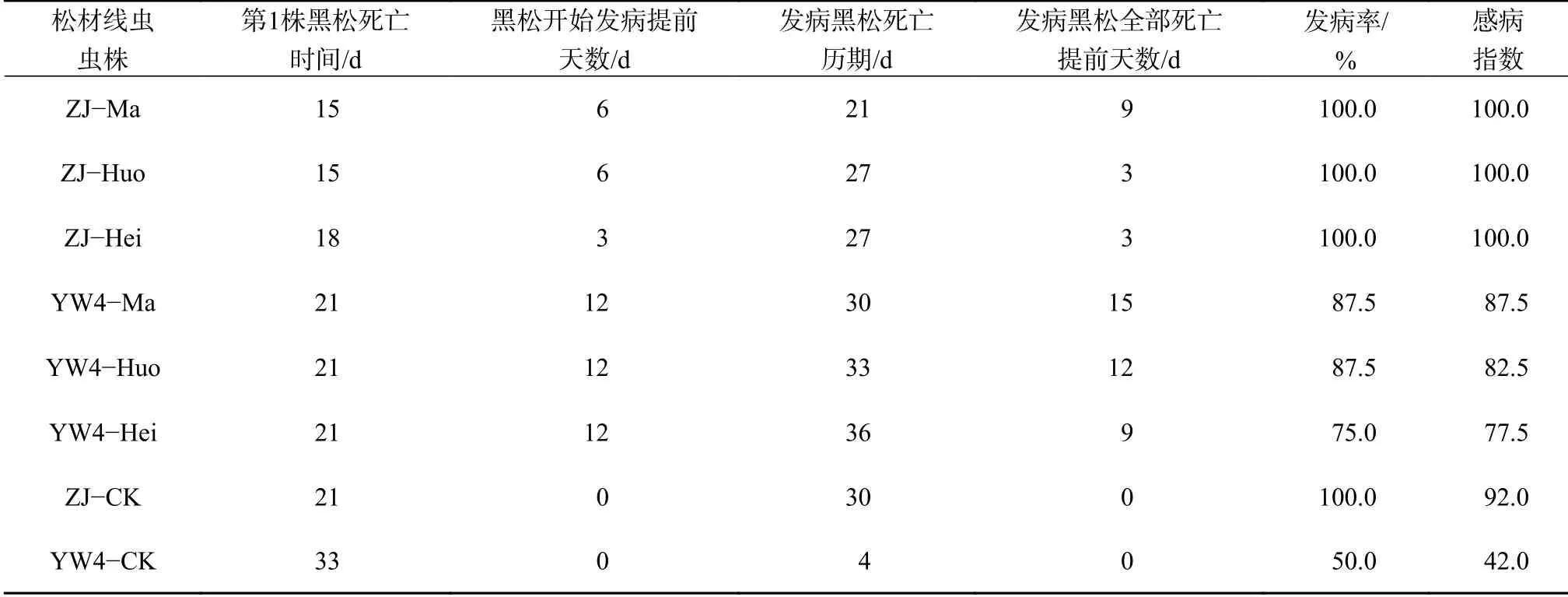
表5 循环接种后不同松材线虫种群的致病力比较Table 5 The comparison of pathogenicity of different B. xylophilus populations after cyclic inoculation
3 结论与讨论
有研究表明,不同寄主和地理来源的松材线虫其致病力存在差异[29−30]。而这种差异与松材线虫侵染不同抗性松树后而产生的获得致病力是否有关并不清楚。在以往有关寄主适应性的研究中,有人认为病原从嗜好性较强的寄主转移到嗜好性相对较弱的寄主后,其寄主适应性会迅速下降[31−32]。本研究中将松材线虫强毒虫株和弱毒虫株接种到不同抗性寄主松幼树中,通过接种—分离—再接种的不断侵染寄主的循环方式,发现松材线虫强毒和弱毒虫株对不同松幼树的致病力均得到了不同程度的提高,这种致病力的提升在弱毒虫株侵染抗性松树上的表现更为明显。同时,强毒松材线虫在侵染抗性寄主马尾松幼树后致病力的增幅显著大于其侵染易感松幼树黑松。另外,无论是松材线虫强毒或弱毒虫株,其在接种抗性马尾松幼树后引起的发病率和感病指数均低于其在火炬松幼树中引起的发病率和感病指数,说明本试验采用的抗性马尾松幼树的抗性较高,对于抗松材线虫病具有较明显的效果。
采用种群相关寄生适应性(RHS)系数来描述不同种群对不同寄主的致病力差异能较好地反映线虫种群和寄主之间的关系。张治宇等在测定松材线虫和拟松材线虫致病性时,表明线虫群体的致病力差异明显,RHS系数变化较大[33−34]。本研究通过对多次接种不同抗性松幼树中的线虫RHS系数的测定,较好的反映了松材线虫种群在寄主体内适应性的变化。在第1次接种测定中,各松材线虫种群的RHS系数在0~10之间,其中,在抗性马尾松幼树体内的弱毒虫株种群不能正常繁殖,线虫数量很少;而随着反复的侵染,线虫的寄生适应性得到提高,在第3次接种测定中,线虫所有种群的RHS系数均处于上升趋势,而在抗性马尾松幼树体内的强毒虫株ZJ−Ma种群的RHS系数甚至超过了第1次测定中在感病寄主黑松幼树体内虫株的水平。该结果表明,无论是强毒还是弱毒松材线虫,通过反复侵染寄主,其种群适应性均可得到较大的提高,最终都可在高抗寄主中生存并增殖。可以推测,如果继续侵染不同抗性寄主,松材线虫弱毒虫株的致病力会得到增强,同时原本高抗的松树种群也可能因线虫种群适应性的提高而抗性下降。
有研究发现,营养条件可以调节生物种群性比[35]。如马尾松松毛虫幼虫在饥饿条件下,性比下降为0. 67,对照性比为0. 95,表明在饥饿条件下存在明显的雄性化倾向,而试验也发现雌虫食量大,资源需求阈值较高,饥饿导致雌虫死亡比例明显偏高[36]。本研究对接种不同抗性松幼树松材线虫种群雌雄性比的变化进行比较发现,在易感黑松幼树体内,松材线虫种群适应性较好,种群雌雄性比也保持在一个相对稳定的水平。而在高抗寄主幼树体内松材线虫种群适应性较低(第1次接种)时,雌雄性比值小于1,种群中雄成虫数量相对较多;随着线虫种群适应性的提高(第3次接种),雌雄性比值也随之升高,比值大于1,说明通过不断侵染寄主其雌虫数量逐渐占据优势,种群繁殖量也随之增长。因此,当线虫种群适应性提高时,其种群繁殖力增强从而使高抗松幼树抗病力下降。在对松材线虫相关基因的研究中发现,当沉默与松材线虫生存相关基因如半胱氨酸蛋白酶基因(Bx-cpls)及自噬相关基因(Bx-ATG1及BxATG8)后,松树体内的线虫数量相应减少,且松树的发病推迟。由此说明线虫的繁殖力减弱相应地会降低其致病力[37−38]。
松材线虫种群分泌的纤维素酶一直被认为是松材线虫可能的致病因素之一,相关研究也反映了纤维素酶在发病过程中所起的一定作用,即松材线虫侵染松树过程中,可通过分泌纤维素酶软化松树细胞壁,从而有利于其侵染[39−43]。本研究比较了来自不同抗性松幼树上松材线虫纤维素酶活性的变化。结果表明,强毒虫株的纤维素酶活性整体高于弱毒虫株,接种高抗寄主幼树的松材线虫种群的纤维素酶活力明显高于接种低抗寄主幼树的松材线虫种群;循环接种后,接种不同抗性寄主幼树的松材线虫种群的纤维素酶活力随着接种次数的增加均有一定程度的提高。这暗示在多次接种过程中,松材线虫致病力的增强与松材线虫分泌的纤维素酶活提高有一定关系。
上述研究表明松材线虫通过不断侵染松幼树,其在寄主松幼树体内的种群性比会不断上升并趋于稳定,线虫种群持续增殖,分泌的纤维素酶活力增强,从而表现出对寄主适生性和致病力的提高。同时,在自然环境中,松材线虫的寄主发生转换的现象是存在的。因此推测,当线虫从易感寄主由媒介昆虫转换至高抗寄主时,其种群适应性最初还不高,短期内可能并不会对寄主产生严重影响,但如果这种情况持续发生,存活的线虫种群的寄生适应性就会不断提高,导致线虫致病力增强而最终成功侵染为害抗病的寄主种群。本研究结果有助于解释在松材线虫病传入我国初期,大多数马尾松是较抗病的而如今马尾松普遍发病的现象。至于目前松材线虫在我国的寄主范围不断扩大是否与此有一定关联尚待进一步研究。
松材线虫无论是强毒还是弱毒虫株,均可通过不断侵染松幼树的方式提高其种群适应性、雌雄性比和分泌的纤维素酶活性,进而增强其致病力,且循环侵染高抗松幼树的增强效应更为显著。本研究结果为进一步揭示松材线虫病在我国的发生流行规律提供了重要的科学依据。
猜你喜欢
美食(2022年12期)2022-12-09 02:08:48
浙江林业科技(2021年3期)2021-07-13 09:03:56
农村百事通(2019年23期)2019-12-30 09:47:25
美食(2019年7期)2019-09-10 07:22:44
NBA特刊(2018年21期)2018-11-24 02:48:04
故事作文·低年级(2018年9期)2018-09-17 18:35:32
长江大学学报(自科版)(2018年14期)2018-07-17 08:37:02
科学大众·小诺贝尔(2018年12期)2018-02-22 12:05:38
长江大学学报(自科版)(2013年23期)2013-10-27 08:45:37
中国瓜菜(2012年1期)2012-04-29 00:44:03
