
武夷山米槠林群落凋落物动态及与气象因子的关系
2022-08-26 10:59:22黄石德
西南林业大学学报 2022年4期
黄石德
(1. 福建省林业科学研究院,福建 福州 350012;2. 福建武夷山森林生态系统国家定位观测研究站,福建 武夷山 354315)
森林凋落物是指由植物地上部分生产并归还回地表的所有有机物质的总称,通常由落叶、落枝、花果繁殖器官及碎屑等部分组成,大量的有机质和矿质元素通过凋落物从植物的冠层传输到土壤表面[1−3]。叶片的凋落被认为是植物应对不利生长条件的一个重要生存策略,如天气变冷或土壤变干等[4−5]。在一定程度上,凋落物量在森林生态系统动态、碳和养分循环及森林生产力等方面起着关键性的作用[6−8]。因此,森林凋落物是森林生态系统整体功能的良好指标[9]。森林生态系统中观察到凋落物的季节格局,主要呈单峰模式、双峰模式或不规则模式[10]。森林凋落物的时间变异是产生森林碳循环不确定性的重要来源。研究表明森林类型、起源、林龄、树种丰富度、物候节律及遗传特性等生物因子是影响凋落物季节格局的重要因素[11−13]。如袁方等[14]报道凋落物量总体表现为雨林 > 常绿阔叶林 > 针阔混交林 > 落叶阔叶林 > 针叶林;常绿阔叶林凋落物量的季节格局比亚热带松林相对稳定,这可能与其演替阶段的不同有关[15]。此外,气温、降雨、风速、太阳辐射等气象因素也影响凋落物的季节格局[10,16−19]。如房焕英等[20]在沙县官庄林场杉木(Cunninghamia lanceolata)人工林开展连续7 a凋落物监测,结果发现其凋落物量对降水量和风速最为敏感,其次为气温与日照时间。在东北长白山红松(Pinus koraiensis)混交林发现月均蒸散量和温度是月凋落物量最佳的预测因子[9]。然而,有关凋落物量与气象因子关系的相关研究仍较为有限,特别在长期定位监测方面。此外,近年来气候变化对凋落物的影响成为凋落物研究领域的热点。然而,森林凋落物量及其季节格局受气候变化影响程度仍不明确。因此,探讨关于气象因子对森林凋落量的影响是很有必要的,有助于更好地理解森林生态系统对气候变化的响应过程。
武夷山国家公园保存有我国东南大陆面积最大、最完整的中亚热带常绿阔叶林森林生态系统。武夷山地区山高坡陡,降雨丰富,极易受到气候变化的影响[21]。而有关的气候变化情景模型预测未来我国亚热带地区将面临着气温升高、降雨减少的场景[22]。武夷山森林凋落物量的季节和年变化格局已有报道[23−25]。然而,有关气象因素对凋落物量和季节性的关系却知之甚少。鉴此,本研究选择武夷山米槠林群落作为研究对象开展了其凋落物及其组成连续4 a(2016—2019年)的定位监测,并结合同期研究区域内的气象数据,通过分析米槠(Castanopsis carlesii)林群落凋落物及其组分的年、季节动态变化,及其与气象因子的相互关系,揭示影响凋落物量的气象因素,为深入认识森林凋落物量在气候变化背景下的变化规律及其对森林生态系统的影响提供科学依据。
1 研究区概况
武夷山国家公园处于福建省北部(27°31′20″~27°55′49″N,117°24′12″~117°59′19″E),其周边分别与福建省武夷山市、建阳区、邵武市、光泽县及江西省铅山县毗邻,总面积1001.4 km2。该地属中亚热带季风气候区,2016—2019年年均气温为15.9 ℃,最冷月(1月)均温为6.9 ℃,最热月(7月)均温为24.0 ℃;2016—2019年年均降水量为2765 mm,降雨量丰富,降雨量多集中在春夏两季(3—8月)约占年降雨量的77.7%,秋冬两季则较为干旱,仅占全年降雨量的22.3%(图1)。武夷山国家公园保存有全球同纬度最完整、最典型、面积最大的中亚热带森林生态系统,地带性植被为亚热带常绿阔叶林,主要分布在海拔400~1100 m处,壳斗科(Fagaceae)是构成其森林群落的主要建群种、优势种[26]。

图1 2016—2019年武夷山站月平均气温和降雨量Fig. 1 Monthly average air temperature and rainfall of Wuyi Mountain station from 2016 to 2019
实验区样地位于福建武夷山森林生态系统国家定位观测研究站(以下简称为武夷山定位站)以西的米槠林群落内,林龄超过70 a,林内郁闭度为0.9,平均林分密度为2980株/hm2,米槠平均胸径为19.97 cm,平均树高为17.6 m,群落层次分明,米槠为样地所在群落优势种。伴生树种主要有树参(Dendropanax dentiger)、浙江润楠(Machilus chekiangensis)、罗浮柿(Diospyros morrisiana)和丝栗栲(Castanopsis fargesii)等。灌木层主要有杜茎山(Maesa japonica)、黄毛润楠(Machilus chrysotricha)及柃木(Eurya japonica)等;草本层植被主要有狗脊蕨(Woodwardia japonica)和草珊瑚(Sarcandraglabra)等。
2 研究方法
2.1 样地布设及凋落物收集
在研究区内,选择典型有代表性的区域布设1个40 m × 60 m的固定样地(27°42′32″N,117°44′55″E),海拔631 m,坡度30°,进一步分成24个10 m × 10 m小样方。从中随机选择10个小样方,并在这些小样方内的中心点位置附近布设1个1 m × 1 m的凋落物收集器,共计布设10个。凋落物收集器由孔径为0.5 mm大小的尼龙网制作而成,其距地面40~50 cm水平放置,对凋落物开展长期监测。2016年1月至2019年12月期间,于每月月末收集凋落物,并将收集的凋落物分成落叶、落枝(< 2.0 cm的小枝和树皮)、繁殖器官(花果)及碎屑等4个组分,置于烘箱内,于65 ℃烘干至恒质量恒定后称取其质量,计算凋落物质量、不同组分凋落物量及其占比、月凋落物量与年凋落物量,最后推算出单位面积凋落物量[25]。
2.2 气象数据的收集
气象数据来源于福建武夷山森林生态系统国家定位观测研究站收集的各项气象指标,包括平均气温、最高气温、最低气温、降雨量、最大降雨量、平均相对湿度、太阳辐射、平均风速及极大风速等。
2.3 数据处理
用变异系数表征凋落物量的年度变化情况,通过不同年度凋落物量的标准误除以其均值获得。凋落物季节变化幅度通过观测期间最大凋落物量与最小凋落物量之差除以月均凋落物量获得。用Origin 8.5绘制图表,运用SPSS 19.0软件对结果进行分析,采用单因素方差分析方法比较不同年份、不同组分间凋落物量的差异性;采用Pearson相关系数分析不同组分凋落物量间的相关性;采用相关分析凋落物与气象因素的相关性,显著性水平均为0.05。
3 结果与分析
3.1 凋落物量及其组分年动态变化
如表1所示,2016—2019年米槠林群落凋落物量变化范围为468.30~520.41 g/(m2·a),最小值出现在2018年,最大值则出现在2017年,4a凋落物量均值为(487.24 ± 11.63) g/(m2·a),变异系数为7.85%。
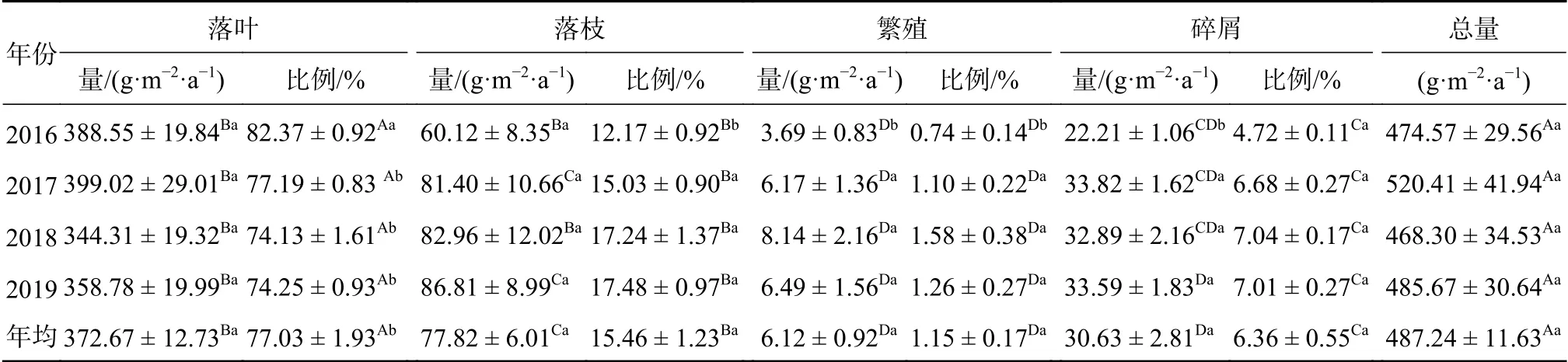
表1 米槠林群落年凋落物量及其组成Table 1 Annual total litter productions and components of C. carlesii forest
2016—2019年间米槠群落凋落物各组分含量表 现 为 落 叶(344.31~399.02 g/(m2·a)) > 落 枝(60.12~86.81 g/(m2·a)) > 碎屑(22.21~33.82 g/(m2·a)) > 繁殖器官(3.69~8.14 g/(m2·a))。其中,落叶占凋落物总量的70%以上,是米槠群落凋落物中绝对优势组分,其年动态与总凋落物量的变化趋势基本一致。落枝的凋落量占比表现为逐年增加的趋势,从2016年的12.17%增加到2019年的17.48%。碎屑的凋落量占比在4.72%~7.04%,其中2016年显著低于其他年份(P<0.05)。繁殖器官的凋落量所占比例最小,其占比仅为0.74%~1.58%。
3.2 凋落物量及其组分年内动态变化
如图2所示,米楮林凋落物总量在年内均表现为双峰曲线,其最高峰出现在4月,占全年凋落物总量的16.4%,尔后逐渐下降,在8月出现谷值,仅占全年凋落物总量的4.1%。而后在11月出现第2个峰值,占全年凋落物总量的9.5%。米槠林凋落物总量的季节变化幅度为229.3%。2016年、2017年、2018年和2019年米槠林凋落物量的季节变化幅度分别为233.3%、104.1%、143.7%和134.8%,其中2016年有最大的季节变化幅度,表明2016年凋落物量的季节变化较为剧烈,而2017年有最小的季节变化幅度,表明该年份凋落物的季节变化较为平缓。

图2 2016—2019年米槠林凋落物总量的季节格局Fig. 2 Seasonal pattern of total litter productions of C. carlesii forest from 2016 to 2019
由图3可知,各组分凋落物量季节变化存在差异。米槠林凋落叶年内变化趋势与凋落物总量的变化基本一致,也表现为双峰曲线,其最高峰也出现在4月(64.46 g/(m2·a)),第2次峰值出现在11月(34.87 g/(m2·a));凋落叶的谷值出现在1月,为14.86 g/(m2·a),仅为峰值的23.1%。观测期间,凋落叶的季节变化幅度为261.6%。与凋落叶年内变化趋势不同,凋落枝出现3次峰值,分别出现在2月、4月和12月,其凋落量分别为9.14、10.58、7.89 g/(m2·a);凋落枝的谷值则出现在8月,仅为峰值的19.1%。观测期间,凋落枝的季节变化幅度为237.0%。凋落繁殖器官则主要集中在4—7月和10—12月,分别占凋落繁殖器官总量的43.4%和30.5%。观测期间,凋落繁殖器官的季节变化幅度为202.1%。凋落碎屑的年内变化趋势与凋落枝相似,其年内也出现3次峰值,分别出现在2、5和11月,其凋落量分别为3.10、3.66、2.84 g/(m2·a);凋落碎屑的谷值出现在8月,仅为峰值的38.5%。观测期间,凋落碎屑的季节变化幅度为156.7%。
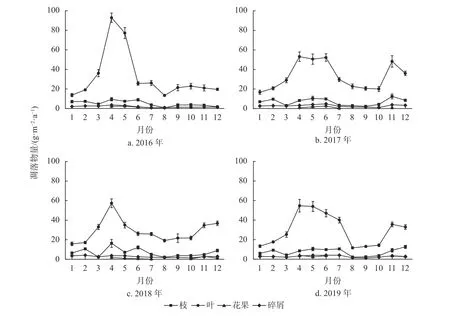
图3 2016—2019年米槠林不同组分凋落物量的季节格局Fig. 3 Seasonal pattern of litter productions for different components of C. carlesii forest from 2016 to 2019
3.3 不同组分凋落物量的相关分析
由表2可知,凋落物量与落叶、落枝和碎屑均呈极显著相关(P<0.01),相关系数分别为0.983、0.671和0.713,与繁殖器官未达到显著水平。落叶与落枝和碎屑均呈极显著相关(P<0.01),相关系数分别为0.529和0.613,与繁殖器官未达到显著水平。落枝与碎屑呈极显著相关(P<0.01),与繁殖器官未达到显著水平。繁殖器官与碎屑呈显著水平(P<0.05),相关系数为0.435。
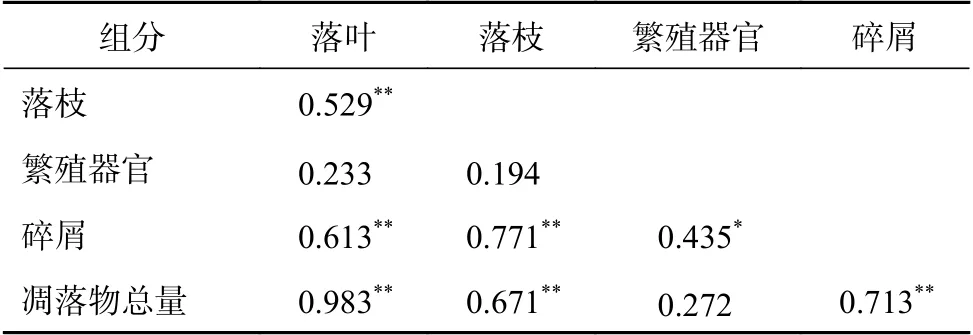
表2 凋落物各组分量相关分析Table 2 Correlation analysis of litter productions for different components
3.4 凋落物及其组分与气象因子的相关性分析
如表3所示,除繁殖器官与当月平均气温、最高气温显著负相关外(P<0.05),其他凋落物组分与当月平均气温、最高气温和最低气温均未达到显著水平。落叶、碎屑与凋落物总量与当月降水量和最大降水量显著相关(P<0.05),落枝和繁殖器官与当月降水量和最大降水量未达到显著相关。凋落物总量与各组分则与当月平均风速、极大风速和相对湿度均未达到显著水平。落枝、繁殖器官和碎屑凋落量则与当月太阳辐射呈极显著负相关(P<0.01)。

表3 凋落物不同组分与气象因子的相关性系数Table 3 Correlation coefficients between litter productions of different components and various meteorological factors
由表4可知,凋落物总量与各气象因子相关分析显示,凋落物总量明显受最高气温、最低气温和平均气温滞后的影响。凋落物总量与当月前2~4个月的最高气温、最低气温和平均气温显著正相关(P<0.05),与当月的前8~9个月的最高气温、最低气温和平均气温显著负相关(P<0.05)。凋落物总量与当月及前1~2个月的日最大降雨量和降雨量显著正相关(P<0.05)。凋落物总量与当月前3~4个月太阳辐射显著正相关(P<0.05)。凋落物总量与平均风速和极大风速均未见显著相关。此外,凋落物总量与当月前5月相对湿度显著负相关(P<0.05)。
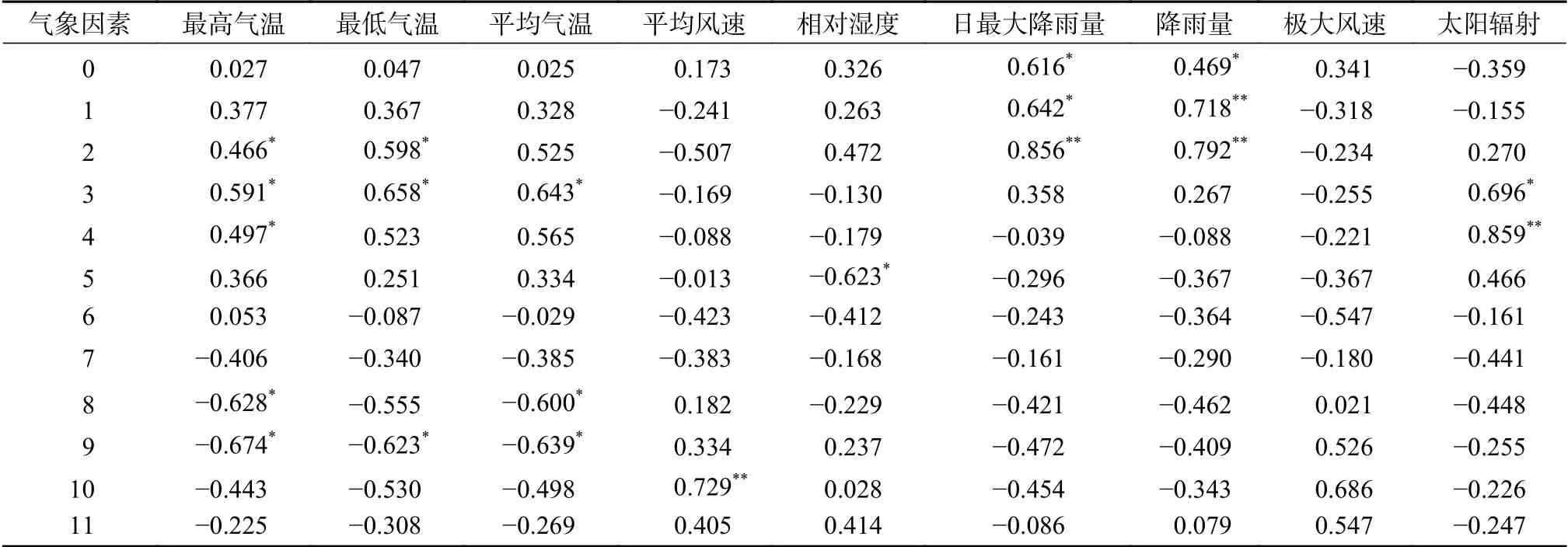
表4 不同月份气象因子与月凋落物总量的相关系数Table 4 Correlation coefficients between meteorological factors from different months and monthly total litter productions
4 结论与讨论
森林凋落物量反映森林生态系统初级生产力水平,也是整体森林生态系统功能的良好指标[9,24]。本研究米槠林群落年凋落物量为487.24 g/(m2·a),略高于同一区域甜槠(C. eyrei)林群落年凋落物量(471.25 g/(m2·a))[25],远低于鼎湖山南亚热带常绿阔叶林(845 g/(m2·a))[27]和尖峰岭的热带山地雨林(768.6 g/(m2·a))[28],明显高于华北落叶松(Larix principis-rupprechtii)(474.0 g/(m2·a))[29]和沂蒙山区(321.2~336.8 g/(m2·a))[30],表明不同气候区间凋落物量差异显著,这主要是由不同气候区间水热差异引起的[28]。
森林凋落物主要由落叶、落枝和其他组分(包括繁殖器官及碎屑等)组成,其中落叶占绝对优势[25]。本研究中米槠林群落凋落叶占凋落物总量的 (77.03 ± 1.93) %,落叶在凋落物总量中占绝对优势,这与前人的研究结果相吻合[9,27,31]。袁方等[14]基于1970—2017年发表的373组中国天然林凋落物数据的分析,认为常绿阔叶林凋落叶占凋落物总量的 (66.43 ± 9.75) %,略低于本研究中米槠林群落凋落叶占比,这主要与凋落繁殖体占比的差异有关,他们报道常绿阔叶林繁殖体占总凋落物量的9.10%,而本研究花果繁殖体仅占凋落物总量的1.15%。此外,本研究米槠林群落凋落物各组分贡献大小顺序为落叶 > 落枝 > 碎屑 > 繁殖器官,这与福建闽北地区米槠天然林叶 > 枝 > 杂物 > 花果的顺序相吻合[32],落叶和落枝构成了米槠林群落凋落物中最主要的组分,两者的动态特征直接决定了总凋落物的季节变化格局。
本研究中,米槠林群落凋落物总量和凋落叶季节动态呈双峰型,分别出现在雨季的4月和旱季的11月,与热带亚热带地区森林凋落物多呈现双峰型的结果是一致的[17,33]。一般认为在中亚热带常绿阔叶林中雨季出现凋落高峰是由于春季开始升温,日照时间变长及土壤水分可用性增加使多数常绿阔叶林新叶大量萌发,并促进衰老叶片的脱落[25];在旱季,植物为了应对水分胁迫发生生理性凋落;而在初冬由于低温以及某些落叶树种的物候规律,进而增加了凋落物量。本研究中,米槠林群落凋落物低谷值则主要出现休眠期的1—2月和生长基本结束的8—9月,这与同一区域甜槠林群落[25]和神农架4种典型森林[34]的观察结果一致。
气象因子一直被认为是影响森林凋落物量的重要因素[1,10]。然而,有关森林凋落物对不同气象因子的敏感程度的研究结果并不完全一致。部分研究报道认为仅气温是影响森林凋落物的气象因素,如在浙江天童山常绿阔叶林[35]和东北主要森林类型[36]的研究认为凋落物量主要受气温的影响,而未受到降水量和风速的影响。这与本研究的结果并不吻合。可能与研究区域降水量的差异有关,本研究观测期间年降水量超过了2700 mm,远远高于浙江天童山1374.7 mm和东北地区450~1340 mm的降水量[35−36],显然丰富的降水改变了凋落物脱落对气温的响应。本研究发现当月降水量、日最大降水量及其太阳辐射是影响凋落物及其组分的重要因素,而与当月气温和风速基本未呈现显著相关,这与Zhang等[10]报道在热带地区降水和辐射是调节森林凋落物的限制因子结论相符的。在干旱季节,水分胁迫导致叶片脱落是植物应对干旱的适应机制[37−38],而潮湿多雨促进植物养分供应或强降水对叶片的物理机制[9,37],进而增加凋落物量。此外,潮湿的热带和亚热带地区森林往往在辐射充足时随着新叶子的出现而使成熟叶片脱落,而在秋季太阳辐射的减少往往又成为秋季叶片脱落的信号[39]。
而本研究发现温度对凋落物量季节动态影响滞后明显,这与鼎湖山常绿落叶阔叶混交林的研究结果一致,这可能是由于本研究对象为天然林,其涵盖了不同生活型的树种,他们有着不同的叶片性状、凋落格局及物候对策[18,40],此时凋落物的季节格局则主要表现为受到物候节律的控制。本研究发现凋落物总量与当月前2~4个月的最高气温、最低气温和平均气温呈显著正相关,这可能是由于温度的升高延迟了叶片的衰老[40],使叶片获得更长的叶寿命,从而延迟了叶子脱落时间,从而增加凋落物后期产量[18]。此外,本研究中繁殖器官与平均气温和最高气温呈显著的负相关,这主要是由于花果繁殖器官的脱落受其自身物候节律的控制,主要出现在温度较低的春季和秋冬之交,此时有较低的气温。
可见,武夷山米槠林群落凋落以叶、枝为主,凋落物总量直接受落叶、落枝凋落量的影响,其中落叶是凋落物的优势组分。凋落物总量和落叶呈双峰曲线,其主峰和次峰分别出现在4月和11月,其谷值出现在8月;而落枝和凋落碎屑年内出现3次峰值,其谷值均出现在8月;凋落繁殖器官主要集中在4—7月和10—12月。当月降水量、日最大降水量及其太阳辐射与凋落物总量显著正相关,而平均气温、最高气温和最低气温对凋落物总量季节动态的影响有滞后性,对于更好地理解在气候变化背景下森林生态系统功能变化具有重要意义。
致谢:感谢福建武夷山森林生态系统国家定位观测研究站的金昌善、何伟及福建省林业科学研究院林捷、高伟、黄雍容在野外监测及数据采集中给予的帮助。该项目还得到福建省森林培育与林产品加工利用重点实验室和武夷山国家公园研究所项目的资助。
猜你喜欢
地下水(2023年5期)2023-10-13 06:03:28
小哥白尼(野生动物)(2022年5期)2022-08-15 08:45:24
今日农业(2021年3期)2021-12-05 01:46:23
今日农业(2021年10期)2021-11-27 09:45:24
公民与法治(2020年17期)2020-10-27 02:28:08
中国化肥信息(2019年1期)2019-01-17 21:31:12
红土地(2018年12期)2018-04-29 09:16:50
作文评点报·低幼版(2018年7期)2018-02-09 16:28:12
植物研究(2017年5期)2017-11-10 01:47:20
五邑大学学报(社会科学版)(2016年2期)2016-03-13 09:24:47
