
酶制剂对水稻秸秆青贮发酵品质及体外消化特性的影响
2022-08-24 05:50:40吴永杰丁浩邵涛赵杰董东代童童尹雪敬宗成李君风
草业学报 2022年8期
吴永杰,丁浩,邵涛,赵杰,董东,代童童,尹雪敬,宗成,李君风
(南京农业大学草业学院饲草调制加工与贮藏研究所,江苏南京 210095)
我国水稻(Oryza sativa)秸秆年产量达23402.0 万t,占农作物秸秆总量的22.4%,但水稻秸秆饲料化利用占比仅为可收集量的6.9%,约20.0%的水稻秸秆被焚烧或乱堆乱放,不仅污染了环境,还造成了资源浪费[1]。因此若能将水稻秸秆通过青贮技术转化为反刍动物饲料对保障粮食安全,促进资源有效利用和农业生态环境保护具有重要意义[2]。青贮可延长饲草的保存时间,提高适口性,缓解因时节和地区差异造成粗饲料供给不均衡的压力,为牛羊等反刍家畜全年提供均衡的日粮。乳酸菌的数量、活性及水溶性碳水化合物含量是影响青贮发酵品质的关键因素。水稻秸秆粗纤维含量高、水溶性碳水化合物含量低、表面附着的乳酸菌数量较少,常规青贮效果并不理想[3-4]。
水稻秸秆粗纤维主要包括纤维素(32%~47%)、半纤维素(19%~27%)和木质素(5%~24%),它们相互交联组成了复杂的网络结构,这种复杂紧密的结构限制了反刍动物对水稻秸秆的消化利用[5-6]。水稻秸秆青贮中添加纤维素酶和木聚糖酶可以降解纤维素、半纤维素,释放水溶性碳水化合物,促进乳酸发酵,快速产生乳酸、降低pH,提高青贮发酵品质[7]。Zhao 等[8]研究表明,水稻秸秆青贮中添加纤维素酶,提高了纤维素转化率和水溶性碳水化合物含量。Souza 等[9]发现添加不同水平的纤维素酶,显著提高了玉米(Zea mays)青贮饲料的体外产气量。纤维素酶和木聚糖酶不仅可打破水稻秸秆的细胞壁结构,也能与反刍动物瘤胃微生物酶协同作用,提高瘤胃对粗饲料的消化率[10-11]。
本试验以水稻秸秆为材料,探讨添加不同酶制剂对水稻秸秆青贮饲料结构性、水溶性碳水化合物组分含量、发酵品质及体外消化特性的影响,为水稻秸秆饲料化利用提供理论依据和技术支撑。
1 材料与方法
1.1 试验设计
设对照组(control,CO),0.3%蒸馏水;纤维素酶组(0.3% cellulose,CE),0.3%纤维素酶;木聚糖酶组(0.3% xylanase,XE),0.3%木聚糖酶;组合酶组(combined enzyme group,CX),0.15%纤维素酶+0.15%木聚糖酶。试验采用完全随机区组设计,共48 个实验室青贮窖(4 个处理×4 个青贮天数×3 个重复),分别在青贮3、7、14 和30 d 后开窖取样分析。
1.2 试验材料
将种植于南京农业大学试验田的水稻于2017 年10 月收获,去除籽粒后,用铡刀切成2~3 cm,并混合均匀。新鲜水稻秸秆的化学成分见表1。酶制剂由中国南京Oddfoni 生物科技有限公司提供,其添加量以商家说明书为准。将配好的纤维素酶、木聚糖酶均匀喷洒于1.3 kg 切碎的水稻秸秆中,对照组喷洒等量蒸馏水,充分混匀后装填于2 L 聚乙烯实验室青贮窖中,压实后密封(密度为650 g·kg-1鲜重),置于室温(25±5)℃下保存。以2 只装有永久性瘘管的健康成年荷斯坦奶牛为瘤胃液供体,于晨饲前收集瘤胃内容物,快速混匀后用4 层纱布过滤于预热至39 ℃保温发酵瓶中,并持续通入CO2维持厌氧环境,密封待用;瘤胃液供体奶牛圈养,日粮组成为干草和精料(1∶1)。

表1 水稻秸秆化学成分Table 1 Chemical compositions of rice straw
1.3 样品处理与测定项目
1.3.1常规营养成分分析 分别于青贮3、7、14和30 d 后开窖,取出全部青贮饲料,充分混匀后,以四分法称取30 g 样品于100 mL 三角瓶中,加入90 g去 离 子水,4 °C 浸 提24 h 后 以2 层 纱布 和 定 性滤 纸过滤,将所得浸提液-20 ℃冷冻保存。浸提液用来测定青贮饲料的pH 值、氨态氮(ammonia nitrogen,AN)、乳 酸(lactic acid,LA)和 挥 发 性 脂 肪 酸(volatile fatty acids,VFAs)。pH 用HA NNA pH211 型(意大利哈纳科技有限公司)pH 计测定;氨态氮含量采用苯酚-次氯酸盐法测定[4];采用安捷伦1260 高效液相色谱仪(德国安捷伦科技有限公司,配备示差检测器和Carbomix H-NP5 色谱柱,流动相:2.5 mmol·L-1H2SO4;流速:0.5 mL·min-1;温度:55 ℃;美国赛分科技有限公司)测定乳酸、乙酸(acetic acid,AA)、丙酸(propionic acid,PA)和丁酸(butyric acid,BA)含量[8]。
水稻秸秆和剩余青贮饲料,65 ℃烘箱中烘干60 h 以上至恒重,测定干物质(dry matter,DM)含量[4];采用KjeltecTM 2300 型全自动凯氏定氮仪测定粗蛋白(crude protein,CP)含量[4];采用ANKOM 2000 纤维分析仪(美国ANKOM 科技公司)测定中性洗涤纤维(neutral detergent fiber,NDF)、酸性洗涤纤维(acid detergent fiber,ADF)和酸性洗涤木质素(acid detergent lignin,ADL)含量[8];按差值计算半纤维素(hemicellulose,HC)和纤维素(cellulose,CL)含量(半纤维素含量由NDF 减去ADF 得出;纤维素含量由ADF 减去ADL 得出)[12];采用蒽酮-硫酸比色法测定水溶性碳水化合物(water soluble carbohydrate,WSC)含量[4];采用安捷伦1260 型高效液相色谱仪(德国安捷伦科技有限公司)测定葡萄糖、果糖和蔗糖含量,配备示差检测器和Skim-pack SCR-101C 色谱柱(流动相:0.3%氢氧化铵乙腈溶液;流速:0.4 mL·min-1;温度:35 ℃,日本岛津公司)[13]。
1.3.2体外消化率及产气参数测定分析 称取1 g 青贮饲料样品(65 ℃烘干后粉碎,过1 mm 筛)于滤袋中[14-15],密封后置于血清瓶中,再向血清瓶中加入10 mL 瘤胃液和40 mL McDougal 缓冲液,置于摇床中培养(39 ℃,150 r·min-1),进行空白校正后,使用压力传感器技术记录4、8、12、24、48、72 h 的产气量[16-17]。试验结束后,从血清瓶中取出滤袋,洗涤后在105 ℃烘箱中干燥3 h,测定DM 、NDF 和ADF 残留量,计算体外消化率[18]。累积产气量(gas production,GP)数据拟合为指数方程[19]。
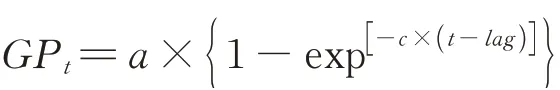
式中:GPt为t时刻累积产气量(mL);a为理论最大产气量(mL);c为产气速率常数(mL·h-1);lag为产气开始时的延迟时间(h);t为读数时间点。
1.4 数据处理
采用统计分析系统(SAS 8.0)GLM 程序对数据进行双因素方差分析(ANOVA),采用Tukey’s HSD 方法对处理间和青贮天数间平均值进行多重比较(P<0.05)。
2 结果与分析
2.1 酶制剂对水稻秸秆青贮发酵品质的影响
酶制剂、青贮天数及交互作用显著影响水稻秸秆青贮过程中LA、AA、PA、BA 含量和LA/AA(P<0.05)。随青贮时间延长,各处理组LA、AA、PA、BA 和总VFAs 含量逐渐提高(表2),青贮30 d 各处理组均显著高于青贮3 和7 d(P<0.05);CO 组LA/AA 随青贮时间延长持续降低(P>0.05),CE、XE 和CX 组先升高后下降,青贮14 d达到最大值,显著高于青贮3、7 和30 d(P<0.05)。青贮30 d,CE、XE 和CX 组LA 含量和LA/AA 显著(P<0.05)高于CO 组,AA、PA、BA 和总VFAs 含量显著降低(P<0.05);与CE 和XE 相比,CX 组有较高的LA 含量、LA/AA 和较低的AA、PA、BA 和总VFAs 含量(P<0.05);青贮30 d,CX 组LA 含量达到最大值(34.13 g·kg-1DM),青贮14 d,CX 组LA/AA 达到最大值(4.26)。
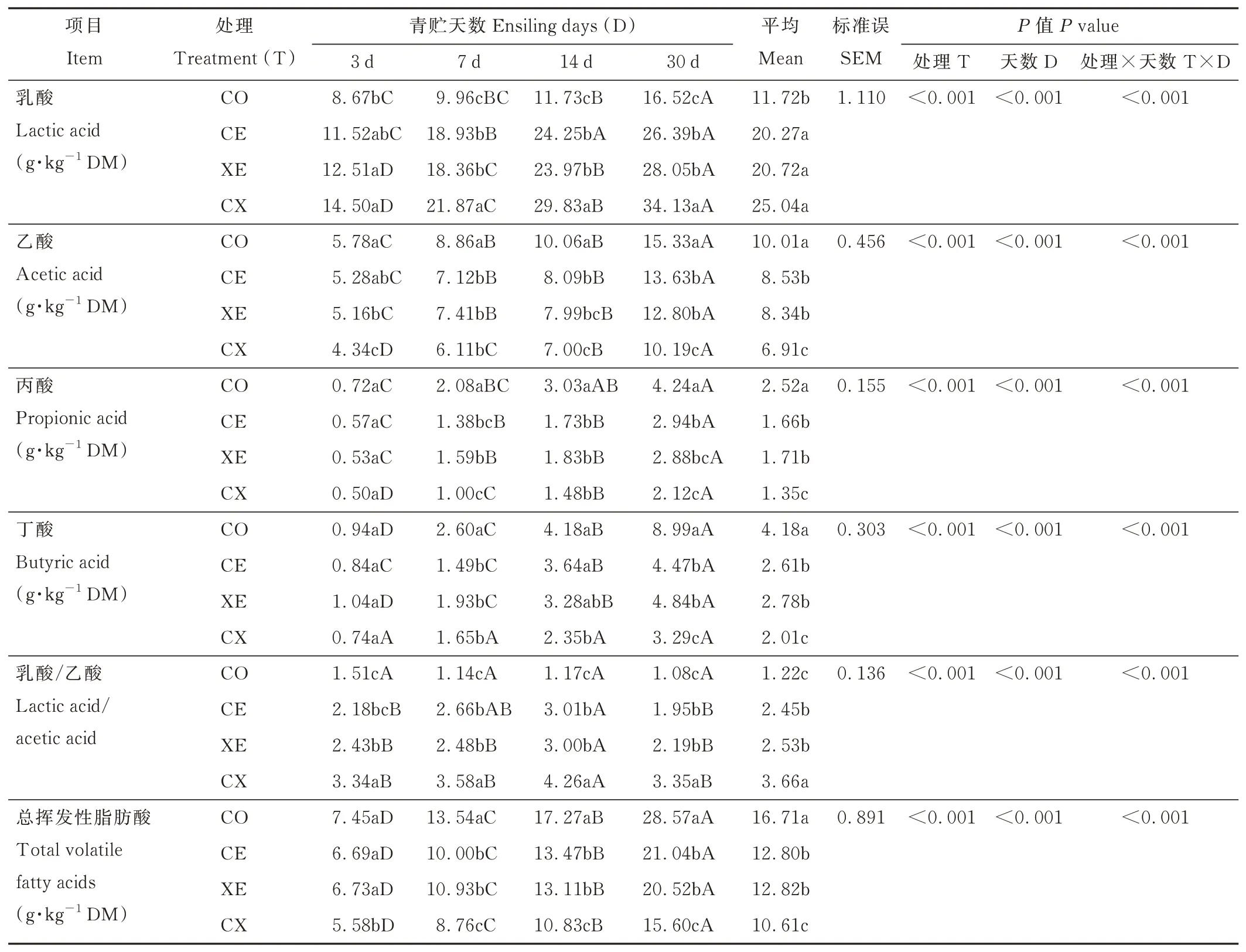
表2 酶制剂对水稻秸秆青贮过程中有机酸含量的影响Table 2 Effects of enzymes on organic acid contents of rice straw during ensiling
酶制剂和青贮天数显著影响水稻秸秆青贮过程中pH、DM 损失、DM 和AN 含量(P<0.05)(表3)。随着青贮时间的延长,各处理组pH、DM 含量逐渐降低,DM 损失和AN 含量逐渐提高,青贮7 d 后各处理组(除CE 和XE组)pH 均显著低于青贮3 d(P<0.05),DM 损失(除XE 组)均显著高于青贮3 d(P<0.05);青贮14 d 各处理组AN含量均显著高于青贮3 d(P<0.05)。CX 处理组中pH 和DM 损失平均值均显著低于CE 和XE 组(P<0.05),青贮30 d,CX 组的pH 达到最低值(4.36)。
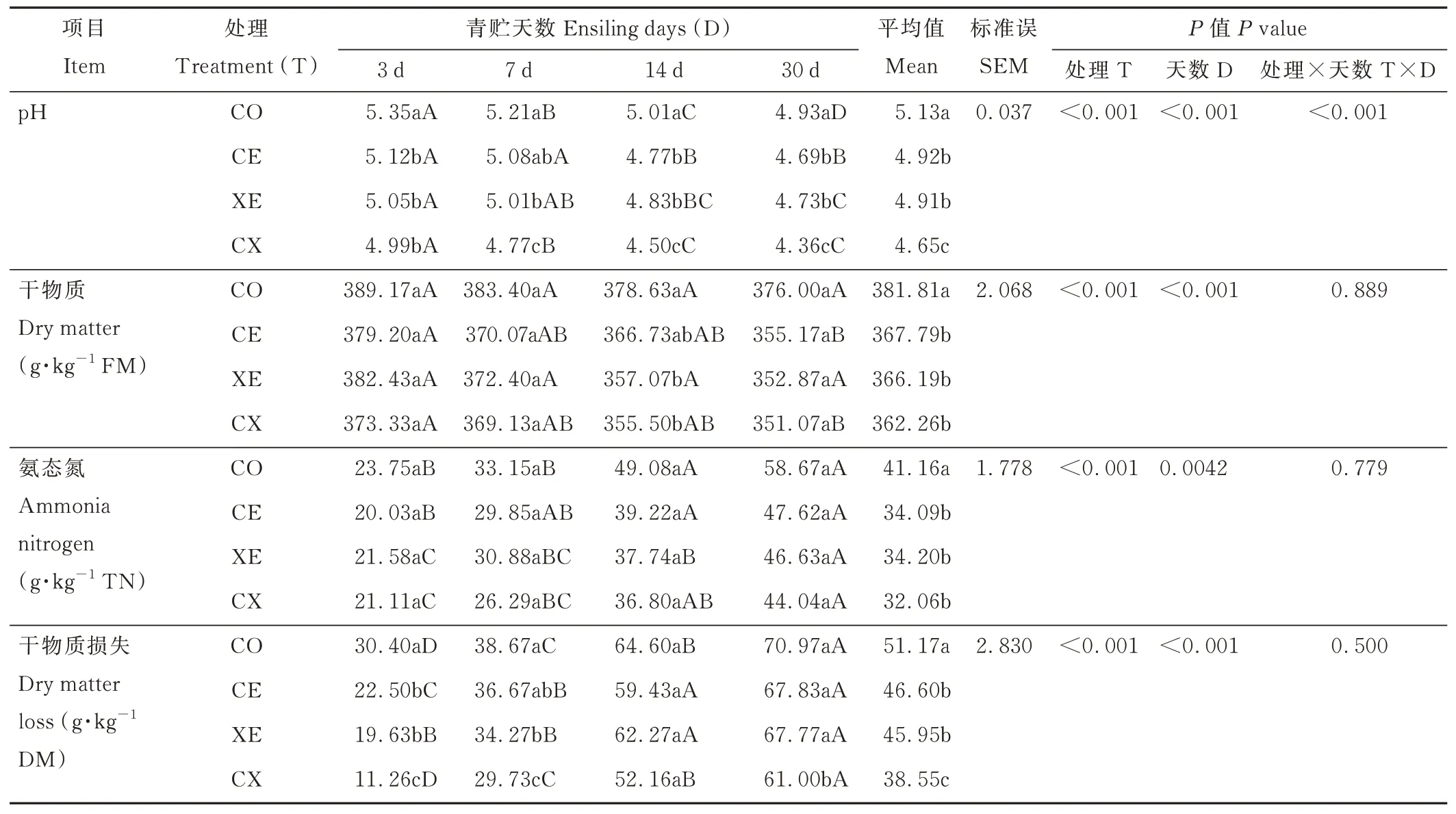
表3 酶制剂对水稻秸秆青贮过程中pH、干物质、干物质损失和氨态氮含量的影响Table 3 Effects of enzymes on pH,DM,DM loss and NH3-N contents of rice straw during ensiling
2.2 酶制剂对水稻秸秆青贮过程中碳水化合物含量的影响
2.2.1酶制剂对水稻秸秆青贮过程中结构性碳水化合物含量的影响 酶制剂、青贮天数及交互作用显著影响了水稻秸秆ADF、NDF 和CL 含量(P<0.05)。随着青贮时间的延长,各处理组ADF、NDF、CL 和HC 含量逐渐降低(表4),青贮14 d NDF 含量显著低于青贮3 和7 d(P<0.05);ADF 含量显著低于青贮3 d(P<0.05);XE 组青贮14 d 后HC 含量显著低于青贮3 和7 d(P<0.05);CE 和CX 组青贮30 d CL 含量显著低于青贮3 d(P<0.05)。青贮30 d,CE、XE 和CX 组NDF、ADF 和CL 含量显著低于CO 组(P<0.05);XE 组HC 含量显著低于CO 组(P<0.05),CE 和CO 之间无显著差异(P>0.05);CX 组CL 含量显著低于XE 组(P<0.05),CX 与CE 之间无显著差异(P>0.05);CX 组NDF 和ADF 含量平均值显著低于CE 和XE 组(P<0.05);各处理组间ADL 含量差异不显著(P>0.05)。
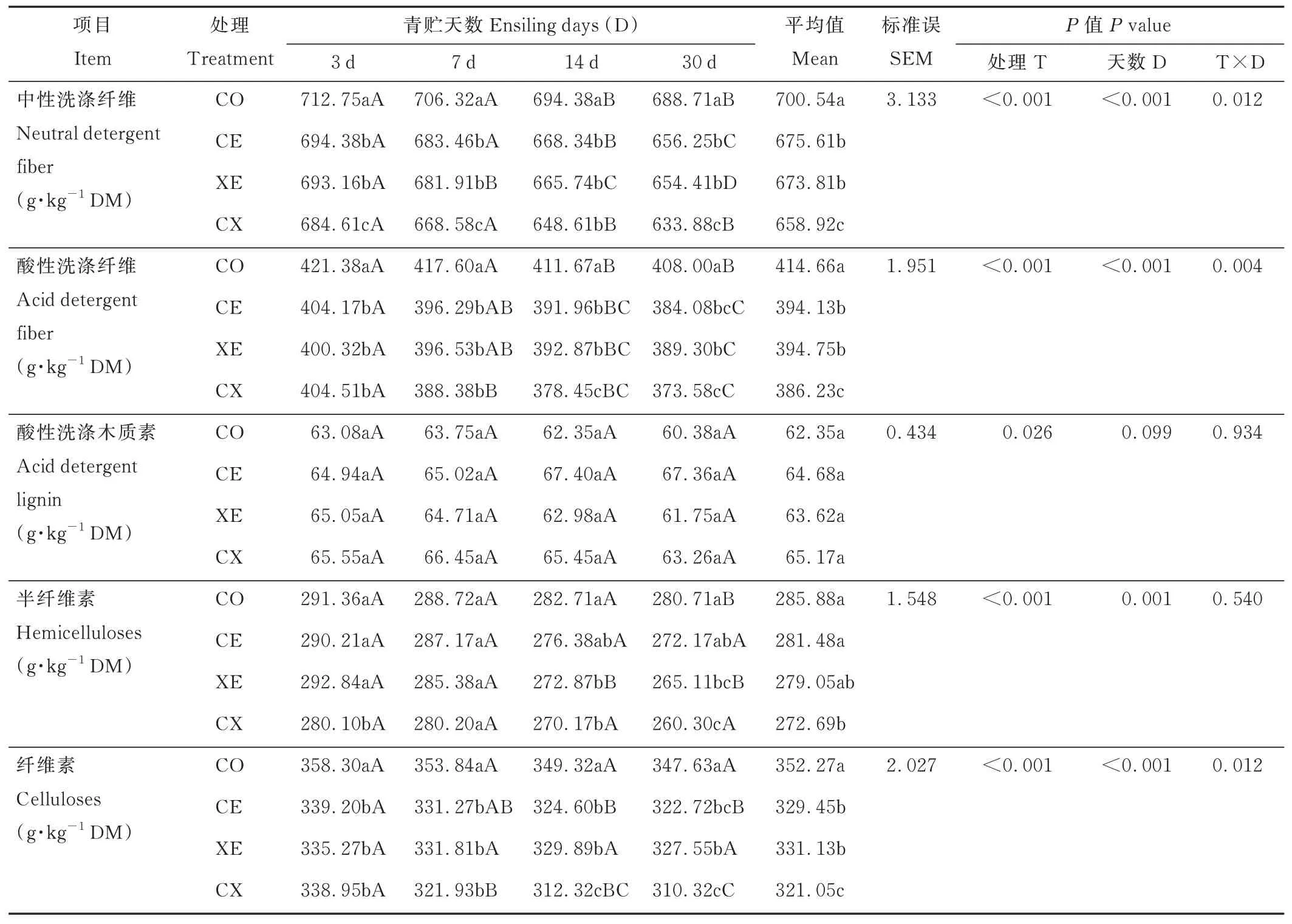
表4 酶制剂对水稻秸秆青贮过程中结构性碳水化合物含量的影响Table 4 Effects of enzymes on structural carbohydrate contents of rice straw during ensiling
整个青贮过程中,CE、XE 和CX 处理组NDF/ADL、ADF/ADL、HC/ADL 和CL/ADL 均低于CO 组,CX 组的此几项值均最低(P>0.05)。青贮7 d 后,CX 组ADF/ADL、HC/ADL 和CL/ADL 均显著低于CO 组(P<0.05);青贮30 d,CX 组中NDF/ADL 显著低于CO 组(P<0.05)(图1)。

图1 酶制剂对水稻秸秆青贮过程中结构性碳水化合物与ADL 比值的影响Fig.1 Effects of enzymes on ratio of structural carbohydrates to ADL of rice straw during ensiling
2.2.2酶制剂对水稻秸秆青贮过程中水溶性碳水化合物含量的影响 酶制剂、青贮天数及交互作用均显著影响了水稻秸秆青贮过程中WSC、葡萄糖、果糖和蔗糖含量(P<0.05)(图2)。整个青贮过程中,各处理组中水溶性碳水化合物含量均显著下降(P<0.05)。CE、XE 和CX 处理组中WSC 含量先提高后下降,而CO 组中WSC 含量持续下降。青贮过程中CE、XE 和CX 组的WSC、葡萄糖、果糖和蔗糖含量均显著高于CO 组(P<0.05)。青贮3 和7 d CX 组葡萄糖和果糖含量均显著高于CE 和XE 组(P<0.05);除青贮14 d 外,整个青贮过程中CX 组中的蔗糖含量均显著高于其他各组(P<0.05)。
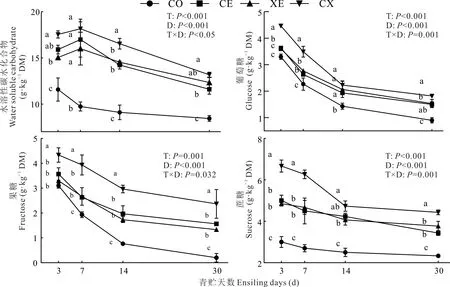
图2 酶制剂对水稻秸秆青贮过程中水溶性碳水化合物含量的影响Fig.2 Effects of enzymes on water soluble carbohydrate contents of rice straw during ensiling
2.3 酶制剂对水稻秸秆青贮饲料体外消化率和产气参数的影响
酶制剂显著提高了水稻秸秆青贮饲料的理论最大产气量、产气速率常数、24、48 和72 h 的累积产气量(图3)和干物质体外消化率(图3),缩短了产气延滞时间(P<0.05)(表5)。CE、XE 和CX 组的理论最大产气量、24、48和72 h 累积产气量和IVDMD 均显著高于CO 组(P<0.05),且CX 组均为最高,分别为68.27、45.90、61.71、66.34 mL 和61.31%;与CO 相比,CE 和CX 组显著缩短了产气延滞时间(P<0.05),CE 和CX 组之间差异不显著(P>0.05);CX 组显著提高了产气速率常数(P<0.05)。各处理组间IVNDFD 和IVADFD 差异不显著(P>0.05)。
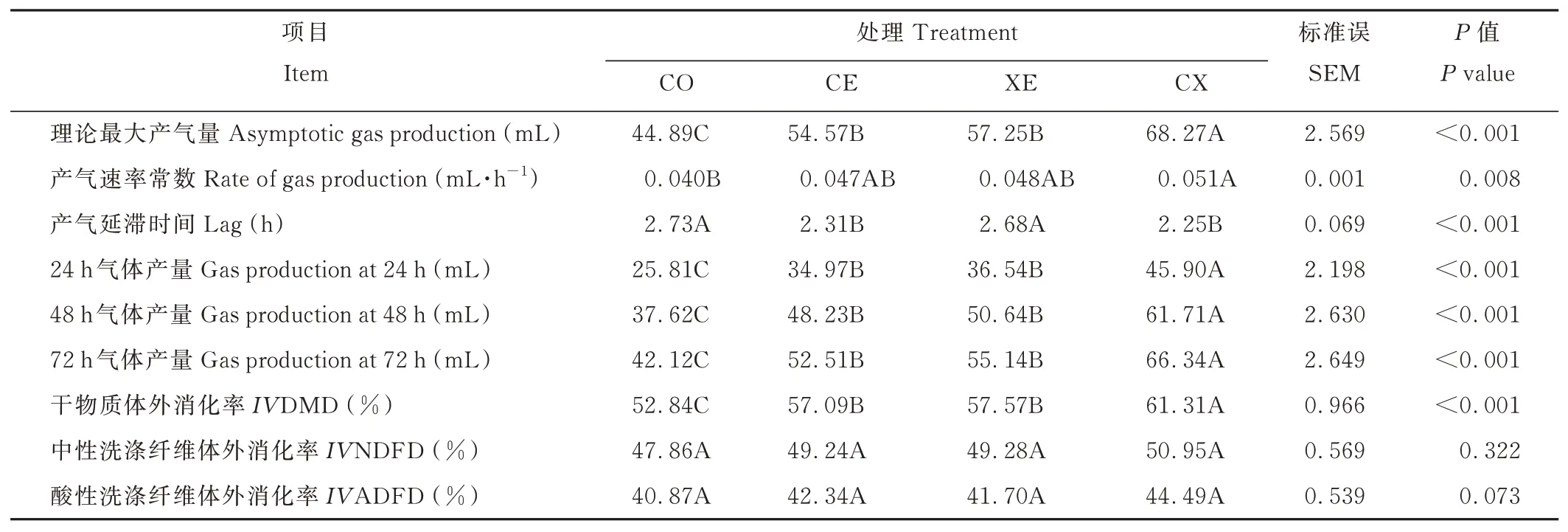
表5 酶制剂对青贮30 d 后水稻秸秆青贮饲料的体外消化率和产气参数的影响Table 5 Effects of enzymes on in vitro digestibility and gas production profile of 30 days rice straw silage
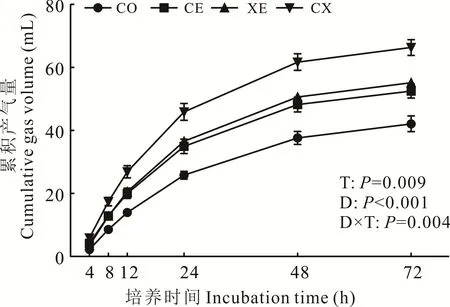
图3 酶制剂对青贮30 d后水稻秸秆青贮饲料体外产气量的影响Fig. 3 Effects of enzymes on in vitro gas production profile of 30 days rice straw silage
3 讨论
3.1 酶制剂对水稻秸秆青贮发酵品质的影响
随着青贮时间的延长,各处理组pH 随LA 含量提高逐渐下降。品质优良的青贮饲料要求pH 在3.80~4.20[20],虽然青贮30 d 各处理组的pH 均高于4.35,但水稻秸秆干物质含量高(393.25 g·kg-1FM),植物细胞渗透压高,趋于生理干旱状态的有害微生物活性被抑制,pH 即使高于4.20 也能使青贮饲料得到良好的保存[21]。赵金鹏[4]在水稻秸秆青贮中添加酶制剂后pH 高于4.45,主要是因为水稻秸秆表面附着的乳酸菌数量(≈4.54 lg cfu·g-1FM)较少所致[13]。CE、XE 和CX 组LA/AA 值逐渐提高,青贮14 d 达到最大值后逐渐下降,说明青贮前期以同型乳酸发酵为主,LA 含量迅速提高,随着青贮时间的延长水溶性碳水化合物含量不足,pH 下降速度缓慢,而耐酸的异型发酵乳酸菌逐渐取代同型发酵乳酸菌成为优势菌群,产生AA,也可将LA 转化为AA,提高了AA 含量[22]。 AA 具有较强的抗真菌能力,青贮饲料中适量的AA 有利于提高饲料的有氧稳定性[14]。PA 和BA 含量过高会影响青贮饲料的适口性,是青贮饲料品质优劣的重要参考指标[21]。李莉等[23]在象草(Pennisetum purpureum)青贮中添加酶制剂,未检测到BA。本试验中,与CO 相比,CE、XE 和CX 组具有较低的BA 和PA 含量,表明水稻秸秆青贮过程中添加酶制剂加快了pH 的降低,有效地抑制了其他有害微生物的活性。
青贮饲料中AN 由植物酶和有害微生物共同作用下,蛋白质和氨基酸等含氮化合物的降解产生[24]。优质青贮饲料AN 含量应低于100.00 g·kg-1TN[23],青贮30 d,各处理组的AN 含量为44.04~58.67 g·kg-1TN,均远小于100.00 g·kg-1TN,可能是因为CE、XE 和CX 组较低的pH 有效抑制了有害微生物对水稻秸秆青贮饲料中蛋白质的降解。
水稻秸秆干物质含量较高难以压实,且节间中空易残留氧气,导致青贮前期有害微生物增殖[4],降低了DM 含量,提高了DM 损失。青贮前期CX 组pH 的迅速降低抑制了有害微生物对水稻秸秆青贮饲料营养物质的分解利用,使DM 损失较小(61.00 g·kg-1DM),这也与CX 组有 较 低的总VFAs(AA、PA 和BA)含量相印证。
CX 组的发酵品质优于CE 和XE 组,庄苏[11]研究发现纤维素酶与木聚糖酶组合处理象草比单一酶处理组LA/AA 值和LA 含量更高,pH、AA、PA、BA、AN 含量和DM 损失更低,发酵品质更好。水稻秸秆中粗纤维含量高,WSC 含量低(≈46.32 g·kg-1DM),单一酶降解结构性碳水化合物,提高青贮发酵品质效果有限[8]。木聚糖酶可降解半纤维素,提高纤维素酶对纤维素的可及性[8],纤维素酶和木聚糖酶协同降解水稻秸秆细胞壁多糖,产生较多WSC,为乳酸菌提供了更多的底物,促进了乳酸发酵,迅速降低pH,抑制了有害微生物活性,提高了发酵品质[11]。
3.2 酶制剂对水稻秸秆青贮过程中碳水化合物组分含量的影响
CE 组纤维素含量显著低于CO 组,XE 组半纤维素含量显著低于CO 组(P<0.05),表明纤维素酶和木聚糖酶对纤维素和半纤维素的酶解作用直接引起了水稻秸秆青贮饲料粗纤维组分含量的降低。Liu 等[25]在高水分苜蓿(Medicago sativa)青贮中添加纤维素酶显著降低了NDF 和ADF 的含量(P<0.05),本试验结果与其一致。CX 组中木聚糖酶降解了部分半纤维素,提高了纤维素的平均孔径和比表面积,从而增加了酶对纤维素的可及性[8],进一步降低了纤维素含量,这与CX 组有更高的WSC 含量相印证。青贮30 d,与CO 组相比,CX 组中NDF/ADL、ADF/ADL、CL/ADL 和HC/ADL 显著下降(P<0.05),证明纤维素酶和木聚糖酶协同作用效果优于单一酶处理。Tian 等[26]研究表明在紫花苜蓿青贮中添加酶制剂对结构性碳水化合物组分影响不显著(P>0.05),本试验结果与其不一致,可能与酶添加量和底物不同有关。此外,本试验中所用酶最适pH 为5.0 和5.5,青贮过程中各处理组最低pH 高于4.36,青贮前14 d 各处理组pH 始终高于4.50,在一定程度上保证了酶的作用时间。
CE、XE 和CX 组的葡萄糖、果糖、蔗糖和水溶性碳水化合物含量在整个青贮过程中均高于CO 组,这主要归因于两个方面,一是纤维素酶和木聚糖酶促进了纤维素和半纤维素的降解,释放了部分葡萄糖、果糖和蔗糖等[27];二是添加纤维素酶和木聚糖酶,迅速降低pH,抑制了有害微生物对水溶性碳水化合物的消耗,节约了更多的水溶性碳水化合物供乳酸菌利用[4]。葡萄糖和果糖是乳酸菌优先代谢的底物[28],青贮开始后乳酸菌增殖,代谢利用葡萄糖和果糖,产生乳酸,葡萄糖和果糖含量快速降低[4]。CX 组水溶性碳水化合物含量始终高于CE 和XE 组,纤维素、半纤维素含量低于CE 和XE 组,也间接证明了纤维素酶和木聚糖酶能协同降解水稻秸秆青贮饲料中结构性碳水化合物,提高水溶性碳水化合物的含量。
3.3 酶制剂对水稻秸秆青贮饲料体外消化特性的影响
许多研究表明[18,29],青贮饲料IVDMD 与各种粗纤维组分含量呈负相关,粗纤维组分含量的降低有利于IVDMD 和产气量的提高。酶制剂可降解青贮饲料细胞壁中结构性碳水化合物,降低纤维素和半纤维素含量,使之结构相对松散,易于进一步被瘤胃微生物附着并降解[8],本研究中CE、XE 和CX 组水稻秸秆青贮饲料中较低纤维素和半纤维素含量使IVDMD 提高了8.04%~16.03%;与CE 和XE 相比,CX 组更低的纤维素和半纤维素含量,使IVDMD 进一步提高。此外,有研究表明酶制剂与瘤胃微生物存在协同作用,能促进瘤胃微生物的增殖和提高对底物的附着能力,提高瘤胃微生物对粗饲料的消化率[11]。IVDMD 的提高表明添加酶制剂能有效提高水稻秸秆青贮饲料的消化率。Agustinho 等[30]研究表明,在全株玉米青贮中添加木质纤维素酶可提高体外消化率。
瘤胃产气主要来自瘤胃微生物作用于瘤胃中的粗蛋白和碳水化合物,单位时间内产气量的多少能反映出反刍动物对青贮饲料中营养物质的消化率[31-32]。瘤胃微生物和青贮饲料特性影响瘤胃产气量,本试验中酶制剂降解了青贮饲料中部分纤维素和半纤维素,改变了水稻秸秆青贮饲料表面结构,有利于瘤胃中纤维降解微生物对青贮饲料粘附和定殖,提高了瘤胃微生物对细胞壁多糖的降解和利用[8,13,33],因此CE、XE 和CX 组理论最大产气量、24、48 和72 h 累积产气量显著高于CO 组(P<0.05)。与CE 和XE 相比,CX 组理论最大产气量(68.27 mL)和产气速率常数(0.051)均有所提高,且在72 h 累积产气量最大(66.34 mL),与CO、CE 和XE 相比分别增加57.50%、26.34%和20.31%,归因于CX 组最低的粗纤维组分含量和最高的IVDMD。 酶制剂有利于促进瘤胃中纤维降解菌和其他类型微生物的生长繁殖[7],提高了消化率和瘤胃菌体蛋白含量,也在一定程度上提高了体外产气量。Morgavi 等[33]研究发现瘤胃微生物酶与外源酶具有协同作用,添加纤维素酶和木聚糖酶可以显著提高纤维素和半纤维素在瘤胃中的消化率。Chen 等[34]报道,添加酶制剂显著提高了芦苇(Phalaris arundinacea)粗纤维组分的体外消化率。丁良[35]和Li 等[36]发现,青贮饲料中可发酵营养成分越多,体外产气量越高,产气参数的改善表明酶制剂提高了水稻秸秆青贮饲料的营养价值。
4 结论
水稻秸秆青贮中添加纤维素酶或木聚糖酶可降解结构性碳水化合物,释放水溶性碳水化合物,提高青贮发酵品质、IVDMD(8.04%~16.03%)和体外产气量(24.67%~57.50%),改善了水稻秸秆的营养价值。组合添加纤维素酶和木聚糖酶能进一步降解结构性碳水化合物,提高水溶性碳水化合物含量、体外消化率、产气量及发酵品质。
猜你喜欢
林产化学与工业(2022年4期)2022-09-13 09:47:12
现代畜牧科技(2021年8期)2021-10-13 07:21:44
湖南饲料(2019年5期)2019-10-15 08:59:12
湖南饲料(2019年5期)2019-10-15 08:59:10
猪业科学(2018年5期)2018-07-17 05:55:56
猪业科学(2018年4期)2018-05-19 02:04:36
广东饲料(2016年2期)2016-12-01 03:43:05
中国酿造(2016年12期)2016-03-01 03:08:21
化工进展(2015年6期)2015-11-13 00:27:42
中国医科大学学报(2015年10期)2015-03-01 02:09:50
