
山羊隐花色素2基因真核表达载体的构建及生物信息学分析
2022-08-06 08:19马白荣张海森高登科靳亚平陈华涛
中国兽医杂志 2022年5期
马白荣 , 张海森,2 , 高登科,2 , 李 超,2 , 靳亚平,2 , 陈华涛,2
(1.西北农林科技大学动物医学院 , 陕西 杨凌 712100 ; 2.西北农林科技大学动物医学院农业农村部动物生物技术重点实验室 , 陕西 杨凌 712100)
为了适应地球昼夜、季节等外界环境条件的周期性更替,地球上的生物随之进化出了一系列具有周期性变化规律的内源性调控机制,这种内源性的调控机制被称为生物钟。狭义上的生物钟指的是昼夜节律生物钟(Circadian clock),它能使生物体的行为活动和生理功能表现出以近似24 h为周期的节律性变化[1]。哺乳动物的中枢生物钟位于下丘脑视交叉上核(Suprachiasmatic nucleus,SCN),能够同步外界光照明暗的周期性变化,将其转换为神经内分泌信号,协调同步肝脏、骨骼肌、肾脏、心脏等组织的外周生物钟(Peripheral clocks),以使机体的内源性节律与外界环境变化相适应[1]。哺乳动物分子生物钟系统主要由脑肌类芳烃受体核转位样蛋白1(Brain and muscle aryl hydrocarbon receptor nuclear translocator-like protein 1, BMAL1)、昼夜运动输出周期(Circadian locomotor output cycles kaput,CLOCK)蛋白、周期(Period,PER)蛋白、隐花色素(Cryptochrome,CRY)蛋白等生物钟蛋白形成的转录-翻译正负反馈环路构成,在分子水平与细胞水平呈现以近似24 h为周期的节律性振荡[2]。
隐花色素2(Cryptochrome 2,CRY2)属于蓝光受体基因家族和光酶解家族成员,是生物钟正负反馈环路中重要的负调节因子。细胞质中的CRY与另一负调控因子PER经过酪蛋白激酶CK1ε/δ的磷酸化修饰后进入细胞核内,抑制正调控因子BMAL1-CLOCK二聚体对下游钟控基因的转录调控活性[2]。此外,CRY2在调控哺乳动物生殖和代谢稳态方面具有重要意义。研究发现,CRY2通过抑制c-Myc-BMAL1-MMP2/9通路来抑制滋养层迁移和侵袭,在胚胎着床过程中扮演重要角色[3]。小鼠在禁食状态下,CRY蛋白通过调节环磷腺苷效应元件结合蛋白(cAMP-response element binding protein,CREB)的节律性表达和G蛋白偶联受体的活性来调节肝脏糖异生的昼夜节律性变化[4]。研究表明,CRY2具有棕色脂肪细胞分化增强剂的作用[5],同时在维持软骨细胞外基质稳态方面也具有重要意义[6]。
有关哺乳动物生物钟的研究主要集中于小鼠和大鼠等啮齿类动物,对牛羊等反刍动物的研究则相对较少。山羊作为支撑我国畜牧业发展的重要经济动物,深入探究山羊生物钟调控生殖和代谢等过程的具体机制对提高畜牧生产效益至关重要。然而,山羊生物钟的分子调控机制目前尚不清楚,有待进一步研究。本试验旨在克隆山羊CRY2基因编码序列(Coding sequence,CDS)并构建其真核表达载体,利用生物信息学软件分析和预测CRY2基因及其编码蛋白的基本理化特性,为后续探究山羊生物钟的生物学功能提供前期基础和关键材料。
1 材料与方法
1.1 主要试剂 DMEM培养基,购自美国Hyclone公司;胎牛血清和0.25%胰蛋白酶,均购自美国Gibco公司;无内毒素小提中量试剂盒和DNA纯化回收试剂盒,均购自北京天根生化科技有限公司;Total RNA提取试剂盒和QuickCutEcoR I,均购自日本TaKaRa公司;反转录试剂盒、高保真PCR用DNA聚合酶和SYBRPremixExTaqTMII试剂盒,均购自日本ToYoBo公司;ClonExpress®II One Step Cloning Kit,购自南京诺唯赞生物科技股份有限公司;琼脂糖,购自美国HydraGene公司;蛋白提取试剂,购自上海碧云天生物技术有限公司;BCA蛋白浓度检测试剂盒,购自江苏凯基生物公司;兔抗CRY2多克隆抗体,购自美国Abcam公司;鼠抗β-actin抗体,购自中国三箭公司;HRP共轭羊抗兔抗体,购自中国中杉金桥公司。
1.2 主要仪器 CO2细胞培养箱和核酸蛋白浓度检测仪,均购自美国ThermoFisher公司;倒置荧光显微镜,购自日本Nikon公司;CFX-96型荧光定量PCR仪、普通PCR仪和电泳仪,均购自美国Bio-Rad公司;凝胶成像系统,购自英国Syngene公司。
1.3 组织样本、质粒、菌种与细胞 山羊卵巢组织取自杨凌区杨沛屠宰场。pcDNA3.1-Puro-N-3HA质粒,购自美国Promega公司。大肠杆菌DH5α感受态细胞,购自北京天根生化科技有限公司。人体肾脏细胞系HEK293T细胞,由中国科学院细胞库提供。
1.4 试验方法
1.4.1 山羊卵巢组织总RNA的提取及cDNA的合成 称取20 mg山羊卵巢组织置于离心管中,加入TRIzol裂解液,采用酚-氯仿抽提法提取总RNA,测定浓度后按照试剂盒说明书将其反转录为cDNA。反应体系(10 μL):模板总RNA 5 μL,5×RT Master Mix 2 μL,去离子水3 μL。反应程序:37 ℃,15 min;50 ℃,5 min;98 ℃,5 min。将获得的cDNA置于-20 ℃保存备用。
1.4.2 引物的设计与合成 根据NCBI数据库中山羊CRY2基因(XM_018059193.1)的CDS区信息,使用Primer Premier 5.0软件进行引物设计,设计完成后添加同源臂(小写字母),即在CRY2的上下游PCR扩增引物的5′端分别加上与pcDNA3.1-Puro-N-3HA载体的酶切位点互补配对的同源序列(下划线表示EcoR I酶切位点),Primer F:agcaagctttcta-gagaattcATGGCGGCGGCGGCAGCGGCGA,Primer R:accggatccgatatcgaattcTCAGACGCCCCTGCTCGGCAGT,引物由西安擎科生物科技有限公司合成。
1.4.3 山羊CRY2基因CDS区的克隆 以卵巢组织的cDNA为模板,通过PCR反应扩增带有同源臂的山羊CRY2基因的CDS区。PCR扩增体系(50 μL):cDNA模板4 μL(50 ng/μL)、上下游引物各1 μL(10 μmol/L),预混液25 μL,添加ddH2O补至50 μL。PCR反应程序:98 ℃预变性5 min;98 ℃变性10 s,55 ℃退火15 s,72 ℃延伸105 s,循环35次;72 ℃总延伸5 min。将PCR产物进行琼脂糖凝胶电泳,并利用普通琼脂糖凝胶DNA回收试剂盒对符合预期的条带进行胶回收。
1.4.4 山羊CRY2基因真核表达载体的构建及鉴定 利用限制性内切酶EcoR I对空载体pcDNA3.1-Puro-N-3HA进行单酶切,经核酸电泳鉴定后,通过胶回收试剂盒回收线性化载体片段。根据同源重组试剂盒说明书,按照一定比例将线性化载体片段与带有同源臂的目的基因片段混合,37 ℃反应30 min获得重组质粒。反应体系(20 μL):线性化载体0.88 μL(71.64 ng),目的基因片段1.36 μL(104.22 ng),5×Buffer 4 μL,ExnaseⅡ 2 μL,ddH2O 11.76 μL。将重组质粒转化大肠杆菌DH5α感受态细胞中,涂板培养后挑取单克隆菌落,在37 ℃条件下摇菌12 h,使用无内毒素质粒小提中量试剂盒提取重组质粒,并对获得的重组质粒进行酶切鉴定,经公司测序后将得到的阳性质粒命名为pcDNA3.1-3HA-gCRY2。
1.4.5 重组质粒转染HEK293T细胞 复苏冻存的HEK293T细胞并进行传代培养,将细胞以密度1×106个/皿接种到60 mm培养皿中,待细胞密度达60%时,使用Turbofect转染试剂将pcDNA3.1-3HA-gCRY2和pcDNA3.1-Puro-N-3HA空质粒分别转染至HEK293T细胞中,转染48 h后分别收取细胞样品用于RNA和蛋白提取,检测山羊CRY2基因在HEK293T细胞中的表达变化。
1.4.6 实时荧光定量PCR检测山羊CRY2基因在mRNA水平的表达 转染HEK293T细胞48 h后,分别提取pcDNA3.1-3HA-gCRY2转染组和pcDNA3.1-Puro-N-3HA转染组HEK293T细胞的总RNA,反转录合成cDNA,按照实时荧光定量PCR(qPCR)试剂盒说明书配制反应体系并完成上机检测。qPCR反应体系(20 μL):cDNA 4 μL,PremixExTaqII 10 μL,上下游引物各 1 μL,ddH2O补至20 μL。反应程序:95 ℃预变性3 min;95 ℃变性10 s,60 ℃延伸30 s,共40个循环;95 ℃变性15 s,60 ℃延伸15 s,95.5 ℃变性5 s。根据引物设计原则,使用Primer Premier 5.0设计出能同时扩增山羊和人CRY2基因CDS区的qPCR引物,以人的GAPDH作为内参基因,引物序列见表1,采用2-ΔΔCt相对定量法对样品的Ct值进行定量分析。

表1 引物序列信息Table 1 Primer details
1.4.7 蛋白质免疫印迹(Western blot,WB)检测山羊CRY2基因在蛋白水平的表达 转染48 h后,分别提取pcDNA3.1-3HA-gCRY2转染组和pcDNA3.1-Puro-N-3HA转染组HEK293T细胞的总蛋白,使用BCA检测试剂盒测定其蛋白浓度,并将其与上样缓冲液混匀,煮沸10 min进行蛋白变性以获得蛋白上样样品。WB的操作步骤:配制10%分离胶与5%浓缩胶,置于电泳槽后拔出梳子,从左到右依次加入蛋白Marker和20 μg蛋白样品进行SDS-PAGE;电泳结束后将蛋白转移至硝酸纤维素(PVDF)膜,10%脱脂奶粉中封闭2 h,TBST溶液洗涤4次,每次5 min;4 ℃过夜孵育一抗,TBST溶液洗涤4次,每次5 min;室温下孵育二抗2 h,TBST溶液洗涤4次,每次5 min;将PVDF膜浸泡于配制好的ECL显色液中,利用G:BOX凝胶成像系统曝光,拍照并记录结果。
1.4.8 生物信息学分析 利用NCBI数据库查找并下载山羊(Caprahircus)、小鼠(Musmusculus)、大鼠(Rattusnorvegicus)、人类(Homosapiens)、黑猩猩(Pantroglodytes)、绵羊(Ovisaries)、牛(Bostaurus)、马(Equuscaballus)、鸡(Gallusgallus)和斑马鱼(Daniorerio)CRY2基因的CDS区,利用DNASTAR软件分析各物种CRY2基因CDS区的序列相似性,并应用MEGA 7软件构建系统进化树;利用ExPASy在线软件分析山羊CRY2蛋白的分子质量、分子式、等电点、氨基酸数目、半衰期和不稳定系数等理化性质;利用ProtScale在线软件预测山羊CRY2蛋白氨基酸序列的亲疏水性;分别使用SingalP 5.0和TMHMM-2.0在线工具预测山羊CRY2蛋白氨基酸序列的信号肽区域和跨膜区域;通过SOPMA和SWISS-MODEL在线软件分析预测山羊CRY2蛋白的二级结构和三级结构,并使用STRING工具分析相互作用蛋白。生物信息学相关软件网址及数据库见表2。
表2 生物信息学分析在线软件Table 2 Online software for bioinformatics analysis
1.5 统计学分析 试验结果以平均值±标准误方式表示。采用GraphPad Prism 6.0软件对试验数据进行统计学分析,使用t检验进行差异显著性检验。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 山羊CRY2基因CDS区的成功克隆 以山羊卵巢的cDNA为模板,通过PCR扩增获得CRY2基因带有同源臂的CDS区片段(山羊CRY2基因CDS区为1 791 bp,上下游引物5′端同源臂42 bp),琼脂糖凝胶电泳结果如图1所示,可见一清晰明亮的条带,条带大小与预期结果相符(1 833 bp)。

图1 山羊CRY2基因CDS区的PCR扩增Fig.1 PCR amplification of goat CRY2 gene CDS regionM:DNA分子量标准; 1:PCR扩增产物M:DL5 000 DNA Marker; 1:PCR amplification product
2.2 山羊CRY2基因真核表达载体的成功构建 使用EcoRⅠ酶对pcDNA3.1-Puro-N-3HA空载体和pcDNA3.1-3HA-gCRY2重组质粒进行酶切,并以pcDNA3.1-3HA-gCRY2为模板扩增CRY2基因的CDS区片段,分别将以上获得的3种产物按顺序加样进行琼脂糖凝胶电泳。结果如图2所示,酶切后pcDNA3.1-Puro-N-3HA空载体与pcDNA3.1-3HA-gCRY2重组质粒的第1条带位置相同(5 211 bp);酶切后pcDNA3.1-3HA-gCRY2的第2条带与以pcD-NA3.1-3HA-gCRY2为模板扩增获得的目的基因的条带大小相同(1 791 bp)。将鉴定正确的重组质粒送至公司测序,测序结果与NCBI数据库中XM_018059193.1序列信息完全一致。上述结果表明,pcDNA3.1-3HA-gCRY2真核表达载体构建成功。

图2 酶切鉴定结果Fig.2 Results of enzymatic identificationM:DNA分子量标准; 1:pcDNA3.1-Puro-N-3HA酶切产物; 2:pcDNA3.1-3HA-gCRY2酶切产物; 3:PCR产物M:DL15 000 DNA Marker; 1:Restriction enzyme digested product of pcDNA3.1-Puro-N-3HA; 2:Restriction enzyme digested product of pcDNA3.1-3HA-gCRY2; 3:PCR product
2.3 山羊CRY2基因在HEK293T细胞中的过表达 将空载体和重组质粒转染至HEK293T细胞,分别使用qPCR和WB方法检测CRY2基因在mRNA和蛋白水平的表达变化。与pcDNA3.1-Puro-N-3HA转染组相比,pcDNA3.1-3HA-gCRY2转染组中山羊CRY2基因的mRNA相对表达水平升高约600倍(P<0.001)(图3A)。同时,WB检测到CRY2-3HA融合蛋白在pcDNA3.1-3HA-gCRY2转染组中的表达,而在pcDNA3.1-Puro-N-3HA转染组中未检测到条带(图3B)。

图3 山羊CRY2基因在mRNA(A)和蛋白(B)水平的表达变化Fig.3 Relative changes of goat CRY2 gene expression at mRNA and protein levels*** :P<0.001
2.4CRY2基因在不同物种间的序列相似性比对及系统进化树构建 不同物种CRY2基因CDS区的相似性比对结果如图4A所示,山羊的CDS区与人类、黑猩猩、牛、马、小鼠和大鼠的相似性分别为90.7%、90.5%、93.4%、92.0%、86.5%和86.5%,与绵羊的相似性高达99.2%,而与鸡和斑马鱼的相似性分别为77.1%和68.9%。系统进化树结果如图4B所示,山羊CRY2基因与绵羊、牛、马的遗传距离最近,与鸡和斑马鱼的遗传距离最远。
2.5 山羊CRY2蛋白的理化性质 ExPASy在线工具分析结果显示,山羊CRY2蛋白的理论分子量为66 804.33 Da,分子式为C2 999H4 636N844O847S23,由596个氨基酸组成。各种氨基酸的数量及其出现的频率如表3所示,其中出现频率较高的氨基酸依次为Ala(10.2%)、Leu(9.9%)、Arg(8.4%)和Ser(8.4%)。山羊CRY2蛋白氨基酸序列的理论等电点(pI)为8.90,半衰期为30 h,不稳定系数为48.87,属于不稳定蛋白。使用ProtScale在线软件分析氨基酸序列的亲疏水性,结果如图5所示,山羊CRY2蛋白氨基酸序列大部分位于亲水区域,故推测该蛋白为亲水性蛋白,其中位于序列第522位和第525位的氨基酸疏水性最强,分数均为1.900,位于序列第446位的氨基酸亲水性最强,分数为-2.311。
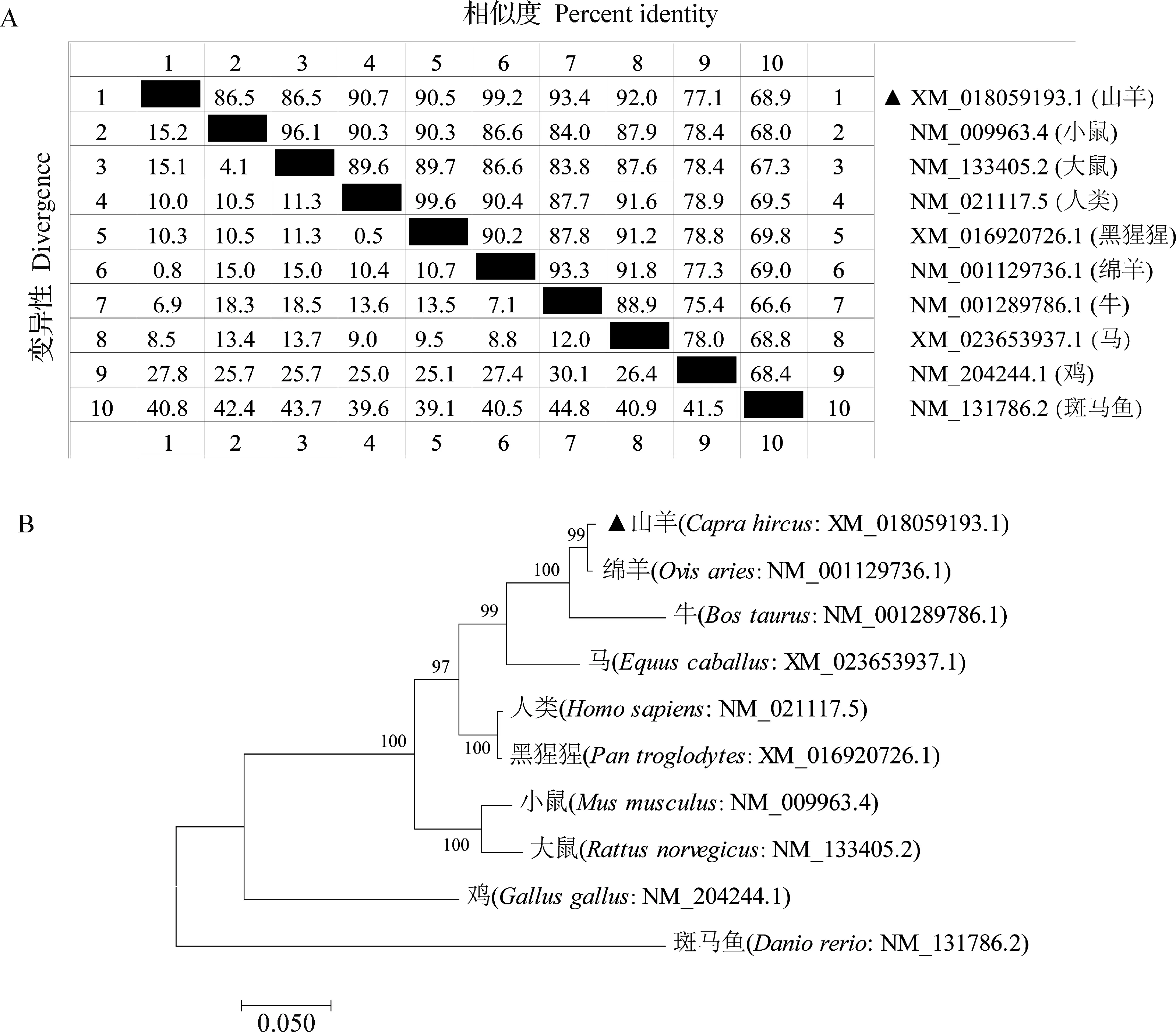
图4 不同物种间CRY2基因CDS区序列相似性比对(A)和系统进化树构建(B)Fig.4 Sequence similarity comparison (A) and construction of phylogenetic trees (B) of CRY2 CDS region in different species▲:本试验获得的山羊CRY2基因▲:Goats CRY2 gene obtained in this study

表3 山羊CRY2蛋白的氨基酸组成Table 3 Amino acid composition of goat CRY2 protein

图5 山羊CRY2蛋白的亲疏水性预测Fig.5 Hydrophilicity and hydrophobicity prediction of goat CRY2 protein
2.6 山羊CRY2蛋白的信号肽及跨膜区域预测 利用SingalP 5.0在线软件预测山羊CRY2蛋白的信号肽区域,结果显示,该蛋白不存在信号肽。利用TMHMM-2.0在线软件对山羊CRY2蛋白进行跨膜区域预测,山羊CRY2蛋白不存在跨膜区。
2.7 山羊CRY2蛋白的二/三级结构预测 使用SMOPA在线分析软件对山羊CRY2蛋白进行二级结构预测,结果如图6A所示,该蛋白二级结构主要包含α-螺旋、β-转角、延伸链和无规卷曲,占比分别为38.26%、5.20%、9.90%和46.64%,其中无规卷曲占比最多。此外,使用同源建模法(SWISS-MODEL软件)对山羊CRY2蛋白的三级结构进行预测,选择模型质量评估得分最高的模型,结果如图6B所示。

图6 山羊CRY2蛋白的二级结构(A)和三级结构(B)预测Fig.6 Prediction of secondary structure (A) and tertiary structure (B) of goat CRY2 protein
2.8 山羊CRY2的互作蛋白预测 使用STRING在线数据库预测山羊CRY2的互作蛋白,预测结果如图7所示,主要包括ARNTL、PER2、FBXL3、PER1、CLOCK、ENSCHIP00000026821、NR1D1、NPAS2、CSNK1D和BHLHE41。

图7 山羊CRY2蛋白的互作蛋白网络Fig.7 Interactive protein networks of goat CRY2 proteinARNTL:芳香烃受体核转位体样蛋白; PER2:周期蛋白2; FBXL3:F-box和富含亮氨酸的重复蛋白3; PER1:周期蛋白1;CLOCK:昼夜运动输出周期蛋白; ENSCHIP00000026821:含蛋白激酶域的蛋白质,属于蛋白激酶超家族;NR1D1:核受体家族1 D组成员1; NPAS2:神经元PAS结构域蛋白2;CSNK1D:酪蛋白激酶1δ,属于蛋白激酶超家族;BHLHE41:基本螺旋-环螺旋家族成员e41ARNTL:Aryl hydrocarbon receptor nuclear translocator-like protein;PER2:Period circadian regulator 2; FBXL3:F-box and leucine rich repeat protein 3; PER1:Period circadian regulator 1; CLOCK:Circadian locomotor output cycles kaput; ENSCHIP00000026821:Protein kinase domain-containing protein,belong to the protein kinase superfamily; NR1D1:Nuclear receptor subfamily 1 group D member 1; NPAS2:Neuronal PAS domain protein 2; CSNK1D:Casein kinase 1δ,belongs to the protein kinase superfamily; BHLHE41:Basic helix-loop-helix family member e41
3 讨论
生物钟是地球上的生物为适应外界环境条件的周期性变化所产生的一种内源性调控机制,可以接收来自环境中的光照、温度以及进食等周期性变化信号,以协调机体能量代谢、激素分泌、血压、睡眠等与环境变化相适应[7-8]。哺乳动物分子生物钟由BMAL1、CLOCK、CRY1/2、PER1/2/3、RORs、REV-ERBs等核心生物钟基因共同构成的正反馈回路组成,其中,BMAL1-CLOCK异源二聚体进入细胞核内与PERs和CRYs基因启动子区域的E-box结合,促进PERs和CRYs基因的转录激活与翻译;当细胞质中的PERs和CRYs蛋白积累到一定程度时,进入细胞核后通过抑制BMAL1-CLOCK异二聚体的转录活性进而抑制其自身的转录,从而形成一个负反馈回路。此外,核受体RORs和REV-ERBs可以结合到BMAL1启动子区域的RORE位点,分别激活和抑制BMAL1的转录[2]。
前期研究发现,CRY2等核心生物钟基因在哺乳动物卵巢中呈现节律性表达,在下丘脑—垂体—性腺轴调控激素释放和靶组织敏感性等进程中发挥重要作用[9-10]。在中枢生物钟SCN中,精氨酸加压素(Arginine vasopressin)的表达受BMAL1-CLOCK二聚体调控,而精氨酸加压素与雌激素协同刺激Kisspeption神经元,进而兴奋GnRH神经元,从而通过垂体—门脉系统调控黄体生成素(Luteinizing hormone,LH)和促卵泡激素(Follicular stimulating hormone,FSH)的释放[11]。在卵巢中,卵泡膜细胞、颗粒细胞和卵母细胞均存在生物钟基因的表达。在大鼠卵巢中,生物钟基因的表达受卵泡发育阶段影响,发育早期的卵泡未检测到生物钟基因的节律性表达,当卵泡发育到窦状卵泡后和成熟卵泡时,其生物钟基因表达则会表现出节律性变化[12-13]。在发育到窦状卵泡期之后的小鼠卵泡中,白天BMAL1和CLOCK的表达水平较高,夜间PER1、PER2、CRY1和CRY2的表达水平较高[14]。此外,在人和小鼠的成熟卵泡颗粒细胞中,StAR、CYP11A1、CYP19A1、HSD3B2等类固醇生成基因的表达都具有昼夜节律性变化[15-17]。上述研究结果表明,CRY2等生物钟基因在啮齿动物和人类的生殖内分泌调控、卵泡发育和类固醇激素合成过程中发挥重要作用,但其调控牛羊等反刍动物生理功能的作用机制尚不清楚,有待进一步的深入研究。
前期研究发现,羊卵巢中的BMAL1、CLOCK、CRYs等核心生物钟基因的表达具有显著的昼夜节律性变化[18],且持续黑暗诱导的生物钟紊乱会改变母羊和胎羊的促卵泡激素、黄体生成素、孕酮和雌二醇的分泌[19]。本课题组前期研究发现,山羊CLOCK基因和BMAL1基因在山羊睾丸间质细胞具有节律性表达,并构建了山羊CLOCK基因和BMAL1基因的真核表达载体,随后通过BMAL1 siRNA干扰和BMAL1过表达等技术证明了生物钟基因BMAL1通过调节山羊睾丸间质细胞中的类固醇生成相关基因的转录来调控睾酮的产生[20-21]。然而,目前关于山羊CRY2的生理功能及其在分子生物钟的具体作用机制仍不清楚,有待进一步探究。本试验利用山羊卵巢组织的cDNA,克隆了CRY2基因的CDS区,成功构建了pcDNA3.1-3HA-gCRY2真核表达载体,并在HEK293T细胞中验证了其表达。HEK293T细胞作为一种质粒辅助细胞,可以使带有SV40启动子的pcDNA3.1质粒具有较高的表达效率[22-23]。本试验将山羊的CRY2基因的CDS区片段克隆至pcDNA3.1载体,并将重组质粒转染至HEK293T细胞,最终实现CRY2基因的高效表达。生物信息学分析发现,山羊CRY2基因CDS区与人和小鼠相比,具有较高的保守性和同源性,表明山羊CRY2基因可能存在与人和小鼠相似的生理功能。蛋白互作分析预测结果显示,山羊CRY蛋白主要与ARNTL、PER2、FBXL3、PER1和CLOCK等蛋白存在相互作用。而在小鼠的研究中发现,CRYs蛋白和PERs蛋白主要作为抑制因子抑制ARNTL、CLOCK对一些生物钟控制基因的转录激活作用,FBXL3可以促使CRY2蛋白发生泛素化并使其降解[24]。由此推测,在山羊生物钟的分子调控网络中,CRY2蛋白与其他蛋白之间的互作可能与小鼠类似,但具体的作用机制有待进一步的探究。
本试验成功克隆了山羊CRY2基因的CDS区,采用同源重组方法构建了pcDNA3.1-3HA-gCRY2真核表达载体,并在HEK293T细胞中实现山羊CRY2蛋白的过表达,对山羊CRY2蛋白的基本理化特性进行了初步预测,为进一步探究山羊生物钟系统的调控机制提供了科学依据。
猜你喜欢
疯狂英语·新读写(2022年6期)2022-11-22
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
今日农业(2021年15期)2021-11-26
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
奥秘(2018年1期)2018-07-02
语文世界(小学版)(2018年2期)2018-03-08
作文周刊·小学一年级版(2017年46期)2018-01-16
小学阅读指南·低年级版(2017年4期)2017-04-24
