
留胚米脂质稳定化及其贮藏关键技术研究进展
2022-07-02 03:49:38王丽群国立东陈凯新郑先哲卢淑雯
食品科学 2022年11期
王丽群,国立东,周 野,陈凯新,郑先哲,卢淑雯,*
(1.黑龙江省农业科学院食品加工研究所,黑龙江 哈尔滨 150086;2.黑龙江中医药大学药学院,黑龙江 哈尔滨 150040;3.东北农业大学工程学院,黑龙江 哈尔滨 150030)
我国是稻米生产大国也是稻米消费大国,水稻的种植和生产直接关乎国家粮食安全。一直以来,我国稻米加工以初级精白米产品为主,不仅带来了严重的资源浪费,而且精白米营养缺失会直接导致以此为主食人群患2型糖尿病的风险增加20%。留胚米最早起源于日本,流行病学研究表明,将其替代精白米作为主食能显著改善以稻米为主食人群的身体机能。稻粒纵向剖面结构及不同类型稻米的结构组成分布如图1所示。其中,糙米为仅去除稻壳后未经碾磨的稻米籽粒;留胚米为适度去除糙米表层蜡质、种皮及部分糊粉层后制得的加工产品;精白米则只保留了胚乳部分,是碾磨加工程度最高的一种稻米产品。由于稻米含有的人体必需营养素中90%以上都存在于稻米外层和胚中,稻米经逐级碾磨后,精白米营养流失最大,糙米虽保留了全部营养但因其口感和蒸煮性较差而不易被人们接受。为兼顾营养和消费需求,留胚米是一种以糙米为原料经适度碾磨后获得的活性稻米产品,除口感与精白米相近外,还含有丰富的脂质、蛋白质、矿物质、维生素及抗氧化物质(如生育酚、酚酸、谷维素)等营养成分。
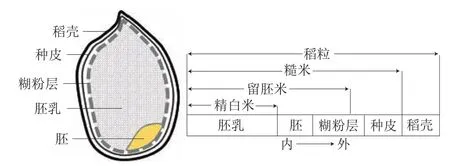
图1 稻粒纵向剖面结构及不同类型稻米的结构组成Fig.1 Longitudinal cross-sectional structure of rice seeds and structural composition of different types of rice
在通过糙米制备留胚米的过程中,糙米的天然保护外层被破坏、脂肪球破裂,这些加工过程为脂肪代谢相关酶与脂质的接触创造了条件。在发生一系列生理活性反应后,留胚米制品的品质最终发生劣变。因此,尽管留胚米在营养指标、感官指标和品质指标上优势明显,但由于其贮藏特性差且无法实现常温长期市场流通,很多企业不会大量生产留胚米,最终导致留胚米只能成为概念米,在市场上占有较小份额。此外,由于稻米类制品对感官品质要求严格以及稻米中脂质劣变诱因的特殊性,至今还缺少理想的留胚米品质劣变解决方案。鉴于此,本文对留胚米中脂质的稳定性进行了系统的阐述与分析,旨在为开展留胚米品质劣变机理研究提供参考,同时为突破留胚米高品质贮藏的产业难题提供新思路。
1 留胚米脂质劣变的理论研究
从稻米籽粒的生理结构上看,留胚米虽然受加工影响比较大,但其营养组成和酶系分布与糙米仍具有一致性。糙米因其表面蜡质及种皮结构的保护作用而能够实现常温条件下的经年保藏,在稻米脂质劣变的理论研究方面,现有研究多以糙米为基础原料,但从糙米外层结构的破坏到加工成为精白米的过程是稻米内部脂质发生劣变的前提条件,而留胚米是加工程度介于糙米和精白米之间的产品类型,因此糙米中脂质劣变理论研究同样适用于留胚米。
1.1 脂质劣变途径
完整稻米籽粒营养全面丰富,但各类营养物质在稻米籽粒中的分布并不均匀,且集中存在于外层结构和胚中,这种分布会导致稻米外层结构的保留程度与稻米品质劣变具有显著相关性。与稻米中蛋白质等营养素的流失和变性不同,稻米脂质的水解与氧化通常会带来不愉快的酸败味或陈化味,直接影响稻米的感官品质,进而导致稻米产品不再适合消费。稻米中脂质质量分数约为1%~4%,与影响稻米食味品质的直链淀粉含量具有相关性。按在稻米中的存在形式,稻米脂质可分为淀粉脂质和非淀粉结合脂质两种类型,其中淀粉脂质是指与淀粉通过单酰基与羰基稳定结合,分布于稻米胚乳中的脂类,含量约为稻米总组分的0.2%~0.7%;而非淀粉结合脂质是指以脂肪球体形式存在于糊粉层(50%~55%)、胚芽(14%~18%)及内胚乳(12%~19%)中的脂类,含量约为稻米总组分的2.9%~3.4%,在磷脂形成的单层膜结构保护下,三酰甘油作为脂质的主要成分稳定存在于脂肪球体内。稻米脂质劣变多以非淀粉结合脂质为底物,且历经3个变化阶段(图2):第一阶段是在机械破坏或磷脂酶水解的条件下,脂肪球发生破裂释放甘油三酯;第二阶段是释放出来的脂质在脂肪酶的作用下水解生成甘油和脂肪酸;第三阶段是脂质氧化,此阶段包括酶促氧化过程和非酶自动氧化过程,脂肪酸经氧化后最终分解生成挥发性的醛和酮等羰基化合物,同时不饱和脂类也会在有氧条件下自发地发生氧化反应,生成过氧化物,并进一步氧化成醛、酮、酸等化合物,产生陈米臭。稻米经氧化反应生成的脂肪酸和过氧化物会与营养成分结合,也会引起稻米蛋白质结构的变化和聚集体的形成,导致稻米的食用和营养价值严重下降。

图2 稻米中非淀粉脂质劣变途径Fig.2 Pathways for non-starch lipid deterioration in rice seeds
1.2 脂质酶促氧化
稻米中天然存在很多参与脂质水解和氧化分解的酶,如脂肪酶、脂肪氧化酶、磷脂酶、过氧化氢酶等。对于未经外力作用的稻米脂肪球体来说,磷脂酰胆碱作为脂肪球膜的主要成分能够在磷脂酶D的作用下降解生成磷脂酸,也有研究认为脂肪球膜降解是脂质降解和酸败的触发器。脂肪球膜破坏后,限制在脂肪球体内的三酰甘油经脂肪球膜渗出,并在脂肪酶的作用下水解,生成以亚油酸为主的次级代谢产物。随后,亚油酸在特异性脂肪氧化酶作用下氧化生成氢过氧化物,在碳链裂开后形成更小的挥发性化合物,如环氧醛酮类、内酯类和呋喃类化合物,具有不愉快气味,造成稻米的酸败。显然,脂肪代谢相关酶的活力在稻米品质劣变过程中发挥了重要作用。在以糙米为原料加工米制品的过程中,脂质代谢相关酶的活力会因加工方式的不同而发生变化,如经超高压和高强度超声技术处理后米制品的脂质水解和氧化能力提高,但发芽处理却可起到延缓脂质劣变进程的作用。此外,脂质酶促氧化可以受金属离子调节,例如脂肪酶活性能被Ca激活,但会被Mn和Cu抑制,当加工处理带来稻米内部矿物质离子的移动与释放时,元素释放的种类、空间分布及处理条件会直接影响稻米脂质劣变进程。研究发现糙米发芽时富集于种皮和糊粉层之间的Ca能迅速移动到胚,而胚中Mg和Mn的含量同时发生显著下降。由此可见,矿物质释放与脂质代谢酶活力之间存在着明显的互作关系,今后研究如能进一步明晰这些活动因子的定量关系以及不同加工处理对酶蛋白金属结合位点特异性损伤的影响,则能从调控酶促氧化反应方向的角度为留胚米脂质劣变问题提供解决思路。
1.3 脂质自动氧化
在稻米品质劣变的研究中,对非酶氧化过程的关注度较低。脂质自动氧化是一个在诱发剂(如温度、单线态氧、光敏剂或辐射)的作用下激活,并由起始、增殖和终止过程组成的连锁反应。从已有报道来看,稻米脂质自动氧化的起始自由基来源无法确定,它可能是一种脂肪酸自由基HL·,也可能是一种完全不同的自由基R·。至今鲜见与稻米脂质劣变起始诱发因子相关的公开研究报道。金属离子,特别是二价态的金属离子(包括Fe、Cu、Mn及Mg),除能担当酶促反应的催化剂外,也是常见的脂肪酸自动氧化引发剂,可以通过单电子转移来降低起始阶段的反应活化能。活性稻米加工过程会对金属离子间的相互作用产生影响,进而调节脂质的氧化和矿物质元素的释放。有研究指出,活性稻米体系中释放的金属离子具有调节脂质氧化的作用,如活性稻米体系内的金属离子螯合剂(如乙二胺四乙酸及其盐类)螯合金属离子后,其氧化能力降低。而硒富集发芽糙米在贮藏期间脂肪酸值和羰基含量均较普通发芽糙米发生显著降低,深入研究发现,硒富集发芽糙米中存在参与脂质代谢的差异蛋白能延缓长期贮藏期间发芽糙米的氧化酸败,硒元素对稻米贮藏期间品质的劣变同样具有抑制作用。由此可见,金属元素在作为脂质氧化反应引发剂及代谢相关蛋白差异表达诱发剂方面的研究也有待展开,而以糙米为原料经适度加工后获得的留胚米制品中金属元素的分布与残留量研究也应得到同样的重视。
2 留胚米品质稳定化技术
2.1 育种稳定化技术
鉴于活性稻米贮藏期间产生米臭多以LOX的代谢产物为主,在基因及其同工酶基因定位研究的基础上,研究者们倾向于开展更多缺失突变稻株的筛选及LOX同工酶基因敲除稻株的研究。尽管水稻中已鉴定出很多与脂质合成相关的基因,但已公布的与脂质代谢相关的基因却较少,且都集中于的研究,这导致基因成了稻米分子育种中遗传修饰的重要靶点。稻米中LOX以营养贮藏蛋白的形式存在于籽粒中,据蛋白质序列解析,LOXs可分为3种类型,而稻米籽粒中常见的LOX多为Ⅲ型LOX的同工酶,即LOX-1、LOX-2和LOX-3。RoyChowdhury等利用RNA干扰诱导基因表达技术分析稻米基因的特异性功能,发现Ⅲ型LOXs的两种同工酶编码基因更偏向于在稻米籽粒中表达,当稻米籽粒萌发时,LOXs以脂质为底物启动代谢反应,在种子成熟、幼苗生长及胁迫应激过程中发挥重要作用;活性稻米贮藏期间,水稻基因又可起到适当延缓稻米籽粒衰老过程的作用。Suzuki等发现缺乏的稻米籽粒耐贮性会提高,其在贮藏期间脂质过氧化程度降低,己醛、戊醛和戊醇等化合物积累导致的陈腐风味减少,而张瑛等通过缺失水稻品种培育,发现和基因缺失在延长稻米种子的寿命方面起着关键作用。从上述报道来看,虽然脂类代谢相关酶类是严重影响稻米陈化和营养品质下降的重要因素之一,但缺失稻米对留胚米加工产业来说并没有带来突破性的改变,除与稻米脂质代谢相关的其他酶类基因的研究需要进一步扩展外,现有基因缺失稻米的营养品质及感官品质变化也需要经实验测定以保证其商品特性。
2.2 酶稳定化技术
研究表明,碾磨制得的留胚米在温度25℃、相对湿度65%的储藏条件下,储藏20 d后脂肪酸值和丙二醛含量明显增高,30 d后留胚米中脂肪酸值已经超过25 KOH mg/100 g干基,米臭味明显,其品质已不适宜食用,由此可以看出,留胚米劣变反应发生迅速且以脂质氧化为主。与留胚米不同,普通稻米的品质劣变虽也以脂质的氧化和水解过程为主,但其劣变时间需要2 年以上,且通常伴有蛋白质和淀粉等组分的规律性改变。在稻米品质稳定方面,为改善稻米酸败对稻米品质产生的严重负面影响,很多研究者陆续开展了与稻米脂质劣变调控相关的研究,探索了很多与稻米脂质代谢反应酶相关的稳定化技术。按照加工方式划分,酶稳定化技术可以分为热加工技术和非热加工技术。在对不同碾磨方式处理后稻米进行微波辐照热加工钝酶的研究中发现,微波辐照后稻米中脂肪酶活性、LOX活性及贮藏后稻米中游离脂肪酸释放程度显著降低,且其降低程度随微波功率的提高而增强,这表明微波加热钝酶技术可延缓稻米中脂质水解;但也有研究发现,热处理会使活性稻米中一些细胞膜结构解体,使更多极性脂质暴露于活性酶之下,因此即使稻米中酶活性有所降低,但其产生的挥发性氧化产物含量反而会更多。此外,因热加工后稻米外观品质破坏严重,一些非热加工技术也已应用到活性稻米钝酶加工工艺中,如低温等离子体、高压钝酶、脉冲电场钝酶技术等。研究表明,各类非热加工技术能获得很好的钝酶效果,如电子束辐照处理后的稻米货架期明显延长;高压处理也能抑制脂肪酶和LOX活性,降低贮藏期内稻米酸败程度;超高静水压处理具有明显提升糙米在贮藏期内抗氧化物水平的作用。但对于留胚米来说,由于稻米商品性要求较高,经非热钝酶处理的留胚米商品性难以保证,截至目前,并没有理想的灭酶技术能够应用于留胚米工业化生产。
2.3 包装稳定化技术
为实现贮藏期稳定,留胚米的包装条件必须充分考虑到物理、化学、生理和生化等多方面反应。由于留胚米易变质,目前市场上流通较多的是小包装加真空或充CO包装,其中充CO包装能充分抑制留胚米的呼吸作用,CO不断地被米粒吸附,一段时间后呈现出与真空包装同样紧密的状态。此外,选择包装材料及方式也可提高留胚米贮藏稳定性,如采用纳米材料包装留胚米,研究认为该包装可以使留胚米酶活性基因表达能力下降,降低留胚米脂肪氧化反应速率,延缓留胚米产品贮藏期间脂肪酸含量升高和米臭味的产生,但目前此项技术尚未得以应用,由于纳米包装调控基因和蛋白表达的作用机理还不够深入,仍需开展后续研究。一项添加吸氧剂的包装实验研究发现保质期内稻米中游离脂肪酸、总生育酚含量及多不饱和脂肪酸绝对含量都有增加,但这种包装处理却能有效抑制脂肪酸的进一步氧化。另有研究发现,贮藏期内颗粒状产品的脂质氧化产物含量显著低于同类粉末状产品,这表明脂质代谢相关酶类及脂肪酸的接触表面积等因素都会影响稻米脂质的劣变程度。但通过隔绝氧气的方式来预防稻米脂质氧化,只能在脂质氧化阶段发挥作用,并没有在本质上阻断脂质在脂肪酶作用下发生水解反应。
3 影响留胚米脂质稳定性的其他关键科学问题
脂质是稻米内部的主要贮藏物质之一,也是稻米细胞的重要组成成分。在水稻生长发育、逆境胁迫、衰老过程中,脂质代谢对其生长适应性和生理效应发挥着重要作用。完整稻谷或糙米中的脂质劣变速度非常缓慢,其原因在于:一是稻米脂质集中分布于籽粒的种皮层、糊粉层和胚中,脂肪酶主要分布于稻谷种皮的横断层中,脂肪酶和脂质因处于不同的空间区域和稳定的活力状态而不利于脂质氧化反应进行;二是休眠的稻米籽粒处于自然生理平衡状态,代谢活动(包括氧化还原反应)亦处于稳定的休眠期。对于留胚米来说,碾磨或铣削等外力破坏了稻米籽粒表层细胞和生理结构,在损伤胁迫的压力下,活性稻米将产生一系列生理应激及稻米自身抗氧化系统失衡问题。解析这些科学问题有助于从本质上提出更先进的控制留胚米贮藏过程中脂质变化的技术手段。
3.1 机械(碾磨)胁迫的影响
脂质特异性探针染色和显微观察实验显示稻米中脂质主要分布于米粒表面,碾磨或抛光处理破坏了稻米籽粒细胞,导致脂肪球体内的三酰甘油释放,虽然脂质代谢相关酶类的分布位置尚无明确报道,但三酰甘油的暴露确为稻米脂质的水解和氧化劣变提供了底物条件。此外,留胚米剧烈的加工过程打破了稻米籽粒的休眠状态,并激发了其生理活性。研究表明,植物细胞在损伤胁迫过程中会发生应答反应,互补DNA(complementary DNA,cDNA)异源表达实验已经证实,植物在经历损伤胁迫时其LOX类化合物生成量增加,而通过LOX代谢途径释放出的活性氧、脂质衍生羰基、丙二醛等多种自由基会进一步增加细胞膜的损伤和细胞死亡。不过,在植物处于损伤压力时,细胞外LOX能通过触发植物防御应答,加速植物愈伤激素(如正己醛、()-2-己烯醛、()-2-己烯醇等)的迅速合成。在对拟南芥中4种13个类型的LOXs(LOX2、LOX3、LOX4和LOX6)的分析研究中发现,LOX2和LOX6能在受伤的叶片中产生大量植物防御激素,如茉莉酸类物质,此类物质是与抗性密切相关的植物生长物质,当植物经历机械伤害时,其含量显著增加,进而诱导一系列与抗逆有关的基因表达。因此,尽管活性氧分子是脂质发生链式反应的关键诱发剂,还是需要对留胚米加工过程中脂肪代谢相关酶类的胁迫应答反应做进一步探究,且已有部分研究已经发表。如冷冻胁迫能显著降低48 h发芽处理后水稻胚中的活性氧水平,尤其是超氧阴离子水平,同时发现很多参与玻璃化-冷冻处理影响氧化应激反应的关键标志基因(/、、、、、和)。此外,高盐胁迫条件下稻米中的抗氧化酶会产生诱导应激,而低湿度条件下,低水分含量与低环境温度都具有延缓糙米中脂肪酸含量增加的作用。以上这些研究均为具有损伤性的胁迫反应直接影响活性稻米中活性氧水平和脂质代谢相关酶类的活力提供了重要证据。但在活性稻米贮藏机制的研究方面,条件胁迫与留胚米应激反应有关的研究鲜见报道,仍需进一步研究明确参与脂质代谢相关酶类是稻米籽粒天然贮藏的还是因结构破坏而应激产生的。
3.2 留胚米抗氧化体系与脂质氧化之间的互作关系
脂质氧化的起始阶段是在引发因子启动下,不饱和脂肪酸分子亚甲基基团RH形成自由基R·,随后进入链传递阶段并形成大量氢过氧化物游离基,即链的传递阶段;当反应生成的自由基产物已不能维持链传递,或者当抗氧化剂或自由基清除剂与链传递过程产生的自由基发生反应时,就到达了脂质氧化的终止阶段。活性氧是有氧代谢过程中不可避免出现的副产物,在正常植物代谢过程中必须维持在亚致死水平。因此,植物体自身发展出一种内在的抗氧化防御系统,通过酶和非酶抗氧化剂来保护自身免受氧化损伤。对于活细胞而言,细胞内氧化还原反应的平衡,除生理维持外,天然抗氧化剂发挥了重要作用。非酶性抗氧化剂,无论是亲水的如抗坏血酸和谷胱甘肽,还是亲脂的如-生育酚和类胡萝卜素,都可以淬灭各种ROS。因此,受到加工损伤的活性稻米,其自身抗氧化防御系统发挥的作用值得研究者们开展深入研究。
3.2.1 稻米中的内源性抗氧化剂
稻米中很多植物化学成分都具有抗氧化活性,如VE、-谷维素、植物甾醇、叶黄素、玉米黄质等类胡萝卜素,槲皮素、芦丁等类黄酮化合物,以及阿魏酸、没食子酸、香草酸、丁香酸等酚酸类物质。白米米糠中酚酸含量最高,其中游离酚酸占到总酸的41%。这些内源性抗氧化剂主要存在于稻米的胚及糊粉层中,不同品种及不同部位含量差异较大。贮藏期内,当酶和非酶抗氧化剂水平发生改变以及产生过多自由基或活性氧/氮时,活性稻米籽粒细胞中这些抗氧化物质则有能力对抗各种氧化应激,保护细胞免受损伤。一项基于13个水稻品种的研究,发现水稻胚芽中VE含量比米糠中高出5 倍,而米糠中-谷维素含量比胚芽中高出5 倍。有研究者在米糠中发现了可能对抗氧化剂提供保护作用的泛醇,并经流行病学和生化试验证实,该物质是一种具有细胞内抑制脂质过氧化作用且能再生的抗氧化剂。目前,活性稻米中内源抗氧化剂的类型及抗氧化方式仍是谷物抗氧化研究的热点。
3.2.2 加工方式对稻米内源性抗氧化剂的影响
稻米加工方式除影响大米营养品质外,也是影响其内源抗氧化剂含量的重要因素。Liu Lei等以粳稻和籼稻为研究对象,发现随着碾磨程度增加,粳稻和籼稻糙米中槲皮素、阿魏酸和香豆酸等9种内源性抗氧化物的含量明显降低;采用碳水化合物降解酶(如纤维素酶)处理稻米可提高酚酸的提取含量;此外,远红外辐射处理技术能起到增加稻壳提取物抗氧化活性和总酚含量的作用;而蒸煮工艺却能降低泰国非糯性紫米品种的花青素和酚类化合物水平及其抗氧化活性。由于游离酚类物质比结合酚类物质具有更强的抗氧化活性,在稻谷陈化和稻米贮藏期间,游离酚类化合物因参与抗氧化反应而使得其抗氧化能力降低。在稻米贮藏研究中,研究者通过加速实验发现稻米中与清除活性氧自由基相关的酶类含量在贮藏期间呈减少趋势,而这些酶类主要集中存在于稻米胚芽中,包括超氧化物歧化酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶等,抗氧化酶类含量的降低加速了稻米中氧化还原反应的失衡进程。因此,过氧化氢酶、过氧化物酶、多酚氧化酶活力可以作为稻米储藏期间品质变化的评价指标。通常情况下,稻谷中过氧化氢酶、过氧化物酶和多酚氧化酶的活性较低,但当稻谷脂质过氧化产物丙二醛得以大量积累时,会刺激稻谷自身氧化还原酶系统,引起酶活性的小幅度增强。由此可知,不同活性稻米中含有的天然抗氧化剂不同,加工方式不同也会对稻米内源抗氧化剂的抗氧化活性产生不同影响。
3.2.3 稻米内源性抗氧化剂对脂质劣变的抑制作用
稻米中天然抗氧化剂与稻米脂质代谢之间存在明显互作关系。同样含有稻米外层结构及胚的米糠中总酚、总黄酮及总花青素的含量与其水解酸败程度呈负相关,这表明米糠中的酚类化合物通过抑制贮藏期间脂肪酶活性,而起到抑制米糠酸败的作用;体外酶活性测定实验发现,黑米中花青素与酚类物质具有抑制猪胰脂肪酶活性的能力;在脱脂米糠中也发现了一种具有1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)清除能力的蛋白类单体或低聚物,该物质能有效清除脂质氧化初期产生的自由基,但对传递期产生的自由基并没有清除作用;此外,在一项红麸皮基因型稻米加工后水解酸度变化的研究中,也证实单宁类天然抗氧化剂与脂肪酶活性的抑制作用有关。从已发表的报道来看,稻米内源抗氧化物对自身脂质抗氧化作用的研究成果仍然很少,活性稻米自身抗氧化体系对其自身脂质的抗氧化能力评价方面还需进行系统的研究并加以解释。在稻米自抗氧化系统的防御调节能力方面,有研究发现添加海藻糖能提高稻米发育细胞的氧化应激能力,进而降低细胞内活性氧含量,但关于此类自抗氧化能力调节实验在加工活性稻米中的研究鲜见公开报道。鉴于稻米抗氧化系统对其脂质酸败的抑制效果,开展活性稻米中内源性抗氧化剂与脂质氧化之间的定量关系和抑制机制的研究,可为开发活性稻米的长期高品质贮藏技术提供重要理论支持。
4 结 语
留胚米是经适度碾磨后获得的一种营养丰富的活性稻米产品。稻米品种、脂质组成、加工(储藏)条件及其抗氧化防御系统等因素均可能影响保质期内留胚米的品质稳定性,但多年来取得的成果尚不能突破留胚米加工产业的技术壁垒,留胚米贮藏难题似乎已无法解决。本文对近年来与活性稻米脂质劣变相关的报道进行归纳整理发现,在留胚米贮藏技术开发方面,留胚米加工带来的损伤胁迫能够激发活性稻米籽粒的应激反应和自抗氧化体系应答反应,但在各种加工条件下,活性稻米脂质代谢反应方向及其抗氧化体系的组成变化仍不清楚,稻米脂质代谢产物与其内源抗氧化剂间的定量关系及稻米内源抗氧化剂延缓脂质酸败的机制也未得到探究。如能对这些问题进行系统地研究与阐释,则可能深入揭示留胚米脂质劣变机制,进而在加工过程中规避留胚米脂质劣变的敏感条件,开发出切实可行的留胚米经年保藏技术或大包装常温保藏技术,最终实现营养丰富、口感良好的留胚米制品安全流通。
猜你喜欢
保健与生活(2023年9期)2023-05-19 21:07:36
美食(2022年5期)2022-05-07 22:27:35
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
中国粮油学报(2019年4期)2019-07-12 09:06:40
中国粮食经济(2018年5期)2018-12-27 08:57:26
中国药理学与毒理学杂志(2015年3期)2015-12-16 09:11:42
时代英语·高三(2014年5期)2014-08-26 16:55:49
应用化工(2014年9期)2014-08-10 14:05:08
食品工业科技(2014年13期)2014-03-11 18:16:55
草食家畜(2012年2期)2012-03-20 13:22:35
