
密码子优化的吡喃糖氧化酶基因在毕赤酵母中的表达
2022-06-09 08:48王玥高庆华董聪罗同阳王庆庆
生物技术通报 2022年4期
王玥 高庆华 董聪 罗同阳 王庆庆
(河北省微生物研究所有限公司,保定 071051)
吡喃糖氧化酶(pyranose oxidase,PROD,简称P2O)又名吡喃糖-2-氧化酶或葡萄糖-2-氧化酶(pyranose-2-oxidase,glucose-2-oxidase),是以黄素蛋白为辅酶、以六元环吡喃单糖类化合物为底物的氧化还原酶类(EC 1.1.3.10),具有多底物催化等特征[1]。序列与结构分析比对发现,吡喃糖氧化酶与葡萄糖氧化酶、胆固醇氧化酶、胆碱氧化酶和甲醇氧化酶一样同属于葡萄糖-甲醇-胆碱(glucosemethanol-choline,GMC)氧化还原酶家族[2-3]。
研究发现,吡喃糖氧化酶在很多方面都起到重要作用,可降解木质纤维素[4],参与碳水化合物的生物转化合成[5],还应用于生物燃料电池、生物传感器以及临床诊断分析中[6]。尤其是在临床诊断分析中,P2O可以用于检测糖尿病人血糖控制的重要指标1,5-脱水-D-葡糖醇。血清1,5-AG检测的敏感性、特异性均较高,且不受忌、进食(餐前、餐后均可测)及性别、年龄、药物等因素干扰,因此P2O具有更好的研究前景。
目前,有一些编码吡喃糖氧化酶的基因序列被克隆且成功异源表达在大肠杆菌中,如隔孢伏 革 菌 属 Peniophora sp.[7]、 多 色 栓 菌 属 Trametes multicolor[4,8]、 黄 孢 原 毛 平 革 菌 Phanerochaete chrysosporium[9]、 彩 绒 革 盖 菌 Coriolus versicolor[10]等白腐真菌中的吡喃糖氧化酶,还有来源于密褐褶孔菌Gloeophyllum trabeum[11]和松口蘑菌根菌Tricholoma matsutake[12]等褐腐真菌的吡喃糖氧化酶。但是这些表达的吡喃糖氧化酶的酶活都比较低,因此提高P2O的酶活就成为研究重点。
研究发现,许多外源基因在大肠杆菌表达体系中高效表达时,往往不能形成有一定空间结构的特定生物功能的蛋白质,而是以一种不可溶、无生物活性的沉淀即包涵体的形式存在于细胞内[13-15]。包涵体形成的原因有很多。研究发现,在表达量低时很少形成包涵体,表达量越高越容易形成包涵体[16]。原因可能是重组蛋白的合成速度太快,表达量短期内太大,大肠杆菌无法及时分泌出蛋白后期加工所需的酶类及辅助因子,以至于蛋白不能进行折叠,二硫键不能正确的配对,过多的蛋白进行非特异性结合,蛋白质无法达到足够的溶解度等[16-17]。通常情况下,重组蛋白的氨基酸组成中含硫氨基酸越多越容易形成包涵体[17]。培养条件不佳,如温度过高或胞内pH值接近蛋白的等电点时容易形成包涵体[18-19]。另外,重组蛋白是大肠杆菌的异源蛋白,由于缺少真核生物中翻译后修饰所需酶类及辅助因子,如折叠酶、分子伴侣等,致使中间体大量积累,容易形成包涵体沉淀[20]。
在真核表达系统中,毕赤酵母分泌表达系统能对许多蛋白质产物进行分泌,便于产物提纯。而且毕赤酵母表达系统具有可诱导的强启动子AOX1,启动外源基因高效表达,还有生长迅速等优点[21]。由于毕赤酵母对密码子的偏爱性,为了消除稀有密码子对蛋白表达量的影响,对目的基因密码子序列进行改造十分必要[22]。目前研究发现,吡喃糖氧化酶只在大肠杆菌中成功异源表达,其在毕赤酵母中表达有待研究。
本研究以NCBI中登录号XM_008046051.1的变色栓菌 pyranose 2-oxidase基因序列为基础,根据毕赤酵母的密码子偏爱性对该基因的密码子序列进行优化,委托公司进行基因合成。经过转化子的筛选,得到了高产吡喃糖氧化酶的重组菌株。在10 L发酵罐中扩大培养,并对其部分酶学性质进行了研究。为吡喃糖氧化酶在真核表达系统中的表达提供试验及应用依据的同时,避免了在大肠杆菌中表达产生的弊端,开辟了一条全新的吡喃糖氧化酶的工业化高效生产途径。
1 材料与方法
1.1 材料
1.1.1 菌株及质粒 吡喃糖氧化酶基因由安徽通用生物公司合成,合成的基因连接在pPIC9K表达载体上;大肠杆菌E.coli DH5α购自上海生工;酵母表达菌株GS115购于优宝生物。
1.1.2 主要试剂 质粒提取试剂盒购自全式金公司;Not I-HF酶、EcoR I-HF酶和Sac I酶均购自NEB 公司;DNA Marker购自康为公司;G418购自phytotechlab公司;ABTS购自Biotopped Life Sciences公司;辣根过氧化物酶购自源叶生物公司;其他试剂为国产分析纯。
1.1.3 培养基 LB培养基、YPD培养基、YPDS培养基、BMGY培养基、YPCS培养基、BSM发酵培养基、PTM1微量溶液的配制均参照文献[23]。
1.2 方法
1.2.1 吡喃糖氧化酶基因序列的密码子优化及合成 参照NCBI中登录号为 XM_008046051.1的变色栓菌 pyranose 2-oxidase基因序列,利用密码子优化软件(http://gcua.schoedl.de/)分析,发现吡喃糖氧化酶基因中存在多处毕赤酵母的稀有密码子,利用密码子优化软件(http://www.jcat.de/)对该基因进行密码子优化,在不改变氨基酸序列的基础上,获得优化后的吡喃糖氧化酶基因序列。在序列的5′端加上EcoR I限制性酶切位点,EcoR I限制性酶切位点为GAATTC;3′端加上6个组氨酸标签后再加上Not I限制性酶切位点,Not I限制性酶切位点为GCGGCCGC,将优化构建的基因序列送安徽通用生物公司合成,合成的基因连接到pPIC9K表达载体上,得到重组表达质粒pPIC9K-P2O。
1.2.2 重组表达载体的验证 安徽通用生物公司提供的质粒总量约为 2-5 μg,用 20-50 μL 灭菌后的双蒸水或 pH 8.0 的 1×TE 缓冲液来溶解,溶解后的质粒浓度约为 100 ng/μL。同时提供含有目的基因的重组质粒的穿刺菌约 500 μL 备用。拿到的穿刺菌可以看到明显的穿刺线,挑取穿刺线上的菌接种到LB固体平板,37℃过夜培养。利用全式金质粒提取试剂盒提取阳性菌株的质粒,通过EcoR I和Not I双酶切验证重组质粒pPIC9K-P2O。
1.2.3 阳性转化子的获得 将验证好的重组质粒pPIC9K-P2O利用限制性内切酶Sac I进行线性化后,电转表达宿主毕赤酵母Pichia pastoris GS115,构建重组表达菌株GS115/pPIC9K-P2O,涂布在终浓度250 μg/mL G418的YPDS固体平板上,30℃培养3 d后获得阳性转化子。
1.2.4 重组蛋白的诱导表达和SDS-PAGE检测 将重组菌株GS115/pPIC9K-P2O挑取单菌落接种于5 mL YPCS(1%酵母提取物,2%蛋白胨,0.5%酪蛋白水解物,0.5%山梨醇)试管液体培养基中,14-18 h后加1%(V/V)甲醇,24 h和48 h均各加1%(V/V)甲醇诱导,72 h收菌,常温12 000 r/min离心5 min收集上清,检测吡喃糖氧化酶的酶活,同时进行SDS-PAGE检测。所有试验均设置3个重复。
1.2.5 吡喃糖氧化酶的酶活测定 吡喃糖氧化酶的酶活测定方法参照文献[4,24]。在反应总体积为1 mL的体系中,依次添加浓度为50 mmol/L、pH 6.5的磷酸钾缓冲液580 μL,浓度为0.01 mol/L的ABTS溶液100 μL,浓度为 1 mol/L 的 D-葡萄糖溶液 100 μL,浓度为100 U/mL的辣根过氧化物酶储液20 μL,添加200 μL酶液启动反应,测定该反应在吸收波长为420 nm处3 min之内的连续变化。
1.2.6 吡喃糖氧化酶的酶学性质分析 酶的最适作用pH和温度以及pH稳定性和热稳定性,金属离子的影响和底物特异性的测定参照文献[24-25]。所有试验均设置3个重复。
1.2.7 10 L发酵罐放大表达P2O 选取试管水平上吡喃糖氧化酶活性最高的转化子研究其发酵罐条件下的表达。挑取单克隆接种于5 mL的YPD液体培养基培养10 h,按 3% 接种量转接于 150 mL BMGY培养基至 OD600≈6接种于发酵罐中。10 L发酵罐的装液量为5.4 L的BSM培养基,灭菌后以10%发酵体积接种,以浓氨水控制pH值,培养温度为30℃。发酵过程分为4个阶段:菌体培养阶段、碳源饲喂阶段、饥饿培养阶段和诱导表达阶段具体内容参照文献[21]。具体方法详见Invitrogen 操作手册。每隔12 h测定表达的吡喃糖氧化酶的活性,并同时进行SDS-PAGE监测表达量的累积。
2 结果
2.1 吡喃糖氧化酶基因的密码子优化及人工合成
在保持氨基酸序列不变的基础上,合成优化的基因序列中共替换了421个碱基。优化后的基因序列通过人工合成的方法获得。基因序列如图1所示,大小为1 904 bp。

图1 密码子优化后的基因序列Fig.1 Gene sequence after the optimization of codons
2.2 重组表达载体的验证
挑取穿刺菌保存管中穿刺线上的菌接种到LB固体平板上,37℃ 过夜培养。利用全式金质粒提取试剂盒提取阳性菌株的质粒,通过EcoR I和Not I双酶切对重组质粒pPIC9K-P2O进行验证。
由图2可知,重组表达质粒pPIC9K-P2O经过EcoR I和 Not I双酶切得到9 kb和1.9 kb左右的条带,分别为载体片段和优化后的目的基因片段,与预期结果一致。

图2 pPIC9K-P2O质粒双酶切验证Fig.2 Verification of pPIC9K-P2O digested by double enzymes
2.3 试管水平检测吡喃糖氧化酶的表达
将获得的阳性转化子GS115/pPIC9K-P2O(图3)按照上述试验条件进行吡喃糖氧化酶在试管中的发酵。利用HRP-ABTS的方法测定吡喃糖氧化酶的酶活力。一个单位的吡喃糖氧化酶酶活(U)定义为每分钟氧化2 μmol的ABTS所需的吡喃糖氧化酶的量。

图3 转化子的获得Fig.3 Acquirement of transformants
首先对不同的单菌落进行了颜色筛选,反应体系添加酶液后由无色变为红棕色,且颜色越深说明酶活越高。随后选择了20个颜色变化明显的单菌落,测定其试管水平上的酶活。结果(图4)显示,有6个阳性转化子诱导72 h后表达产物的酶活在18 U/mL以上,其中4号阳性转化子的酶活较高且稳定,最高酶活约24 U/mL。因此,选择菌株GS115/pPIC9KP2O-4进行下一步试验研究。

图4 筛选P2O高表达量的重组菌株Fig.4 Screening of recombinant P.pastoris with high-yield P2O
将离心后的发酵上清液进行SDS-PAGE蛋白电泳,结果如图5所示,pPIC9K-P2O表达载体转化的重组毕赤酵母在85-100 kD之间有一条条带,且pPIC9K空载体转化的毕赤酵母没有相似条带,说明吡喃糖氧化酶在重组毕赤酵母中实现了外源分泌表达。前期优化基因序列时在3′端有6个组氨酸标签,上清液超滤浓缩后采用Ni2+亲和层析柱亲和纯化蛋白,并进行SDS-PAGE蛋白电泳,结果如图6。

图5 重组蛋白的SDS-PAGE结果Fig.5 SDS-PAGE result of recombinant protein

图6 重组蛋白纯化结果Fig.6 Purification results of recombinant protein
2.4 10 L发酵罐表达吡喃糖氧化酶
将重组菌P.pastoris GS115/pPIC9K-P2O-4按照上述试验条件进行10 L发酵罐放大培养,结果如图7所示。初始发酵温度控制在30℃,用浓氨水控制pH 为5.0,通过调节转速和通气量控制溶氧。等到BSM培养基中的甘油耗尽,溶氧上升后,以12 mL/(h·L)的速度流加含有1.2%(V/V)PTM1的50%(W/V)甘油,待OD600长到150-180 时停止流加甘油,然后饥饿培养0.5-2 h,溶氧开始上升。这时,以3 mL/(h·L)的速度流加含有1.2%(V/V)PTM1 的100%甲醇,用浓氨水控制pH 为6.0,调节转速和通气量控制溶氧。
随着甲醇的添加,吡喃糖氧化酶的酶活逐渐升高,诱导132 h时酶活达到约220 U/mL(图7-A)。对不同诱导时间的发酵上清液进行SDS-PAGE 蛋白电泳,结果(图7-B)显示,随着诱导时间的增加,在85-100 kD处的条带变浓重,与酶活的增长趋势相对应。

图7 10 L 发酵罐高密度发酵生产 P2OFig.7 Production of P2O by high-density fermentation in 10 L bioreactor
2.5 重组吡喃糖氧化酶的酶学性质研究
重组吡喃糖氧化酶作用葡萄糖的最适pH为6.5(图8-A)。在pH 5的柠檬酸盐缓冲液和pH 9的甘氨酸-氢氧化钠缓冲液中,P2O表现出约60%的相对活力。在pH 5-9范围内,其相对酶活高于50%。当pH低于5或高于9时,酶活开始随着pH的降低或升高急剧下降(图8-A)。P2O在较广的pH范围内都表现出较高的稳定性,尤其是对碱性条件有较强的耐性。在pH 4-9的缓冲液中放置24 h,酶活仍在80%以上(图8-B)。

图8 最适pH及pH稳定性Fig.8 Optimal pH and pH stability
P2O氧化葡萄糖的最适温度为55℃(图9-A),曲线呈典型的“钟罩型”。温度高于或低于55℃都导致其酶活下降。在30 min内,该酶在不超过60℃条件下热稳定性良好(图9-B)。当温度升高至80℃时,30 min内 P2O完全失活。
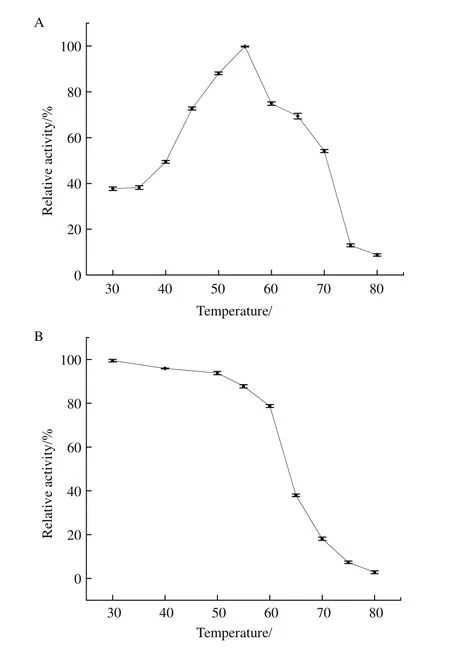
图9 最适温度及温度稳定性Fig.9 Optimal temperature and temperature stability
不同浓度的金属离子对吡喃糖氧化酶的酶活影响如图10所示。金属离子Cu2+对该酶酶活的抑制作用较大,与周亚萍报道的研究结果一致[26-27]。在金属离子浓度为 10 mmol/L 时,Mn2+、Co2+对其有激活作用(图10),分析其原因可能是该浓度下的这两个金属离子与酶结合改变了酶的空间结构(尤其是活性部位的结构),对酶的活性有了促进作用,但其调控机制有待进一步深入研究。P2O的底物专一性较好,以葡萄糖为最适底物(图11),对麦芽糖也有一定活性,对其他单糖或二糖不显示催化活性。

图11 P2O的底物特异性Fig.11 Substrate specificity of P2O
3 讨论
吡喃糖氧化酶(P2O)是以黄素蛋白(FAD)为辅酶的氧化还原酶类,具有多底物催化特性,可催化葡萄糖、山梨糖、木糖等多种糖类化合物。近年来吡喃糖氧化酶越来越受到重视,该酶在降解木质素[2,25]、食品技术[2,28]、制作生物传感器[29]等方面有重要作用。P2O还可以用于检测糖尿病人血糖控制的重要指标 1,5-脱水葡萄糖醇(1,5-AG)[30]。葡萄糖氧化酶和己糖激酶只能作用于β-D-葡萄糖,而吡喃糖氧化酶能作用于α-和β-D-葡萄糖,对葡萄糖有更高的亲和性,因此,吡喃糖氧化酶无需变旋即可检测血液中或食物中的葡萄糖[31]。
目前,P2O已经在大肠杆菌中得到了异源表达,在真核表达系统中尚无报道。并且P2O在大肠杆菌中进行重组表达时容易形成包涵体,表达量低,不易分离,给工业应用带来一定的限制,因此,利用基因工程技术实现吡喃糖氧化酶的大量生产成为了研究热点。毕赤酵母表达系统是目前应用较广、外源蛋白表达效果较理想的表达系统之一[21],与原核表达系统相比,具有很多优点,如生长迅速,遗传操作简单,有可诱导的强启动子AOX1,启动外源基因高效表达,并能正确翻译和翻译后加工,被广泛用于各种具有工业用途的重组蛋白的生产。
研究发现,P2O广泛分布于白腐真菌中,本研究选用的担子菌纲变色栓菌(Trametes versicolor)是白腐真菌的一种,常用于实验和科研检测。我们在NCBI中选取了几个含有吡喃糖氧化酶基因序列的真菌,分别对不同的基因序列进行密码子优化,转化到毕赤酵母中表达,通过酶活力和稳定性的检测发现,本研究选用的基因序列优化后表达的吡喃糖氧化酶的酶活力较高,且稳定性较好。
本研究利用毕赤酵母表达系统表达吡喃糖氧化酶,通过分析影响蛋白表达量的因素,根据毕赤酵母密码子偏爱性对该基因序列进行优化,提高该酶的表达量。合成基因序列的过程中,在该基因序列的3′端加入6个组氨酸标签,可以采用Ni2+亲和层析柱亲和纯化蛋白。在10 L发酵罐发酵培养时,经过132 h酶活达到了220 U/mL。毕赤酵母可通过高密度发酵培养大量生产重组蛋白,同时有利于简化纯化步骤,降低生产成本。本实验还对重组吡喃糖氧化酶的部分酶学性质进行了研究,为该酶的应用和改造提供一系列必要的初始参数。尤其是在血糖检测用酶,要求所用的酶有较高的催化效率,较窄的底物谱,同时需要有良好的热稳定性。
本研究在为吡喃糖氧化酶在真核表达系统中的表达提供了试验及应用依据,同时,有效避免了在大肠杆菌中表达产生的弊端,开辟了一条全新的吡喃糖氧化酶的工业化高效生产途径。
4 结论
本研究利用毕赤酵母表达系统实现了吡喃糖氧化酶的分泌表达,根据毕赤酵母的密码子偏好性对pyranose 2-oxidase基因进行密码子优化,将优化后的基因序列克隆到毕赤酵母中,经过转化子筛选,得到了高产P2O的重组毕赤酵母菌株。最佳培养条件下吡喃糖氧化酶酶活在试管水平达到24 U/mL,在10 L发酵罐中对重组毕赤酵母菌株放大培养,甲醇诱导132 h吡喃糖氧化酶酶活可达220 U/mL。进一步研究了其酶学性质,pH和温度的稳定性良好,并且底物专一性较好。
猜你喜欢
食品与发酵工业(2022年6期)2022-04-01
食品与发酵工业(2022年3期)2022-02-23
河南工业大学学报(自然科学版)(2021年5期)2021-11-15
昆明医科大学学报(2020年12期)2021-01-26
生物学通报(2020年11期)2020-10-22
发明与创新·中学生(2019年6期)2019-06-26
浙江化工(2019年4期)2019-05-13
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年8期)2018-09-10
中成药(2018年7期)2018-08-04
