
不同泌乳期羊乳和牛乳的高通量定量乳清蛋白质组学
2022-06-02 08:42:44吴欣雨
食品科学 2022年10期
张 荣,吴欣雨,贾 玮
(陕西科技大学食品与生物工程学院,陕西 西安 710021)
随着乳制品市场多样化和家庭膳食结构的改善,全球羊乳消费保持稳定增长,促进了人们对羊乳营养特性的探索。羊乳及其制品富含维生素、矿物质、高质量蛋白质、低胆固醇和较小的脂肪分子等营养素,被认为是一种易于消化的低致敏性天然营养源,是牛乳基婴幼儿配方乳粉和母乳的潜在乳源替代物。
羊乳乳清因含有高营养价值蛋白质、大量低聚糖、乳糖和矿物质被认为是功能性成分,具有免疫调节功能和抗菌活性,并可延缓心血管疾病、骨质疏松症、癌症和肥胖症的发病或改善其状况。目前,基于乳清蛋白的配方乳粉可替代普通婴幼儿配方乳粉,避免婴幼儿对全脂牛乳或大豆蛋白的胃肠道不耐受。Sousa等发现,与其他反刍动物乳相比,羊乳中的低聚糖与母乳更相似,可作为一种重要营养素添加至牛乳配方中,以达到抗致病性、抗黏附性和益生元作用。De Souza Araújo等研究表明,羊乳乳清能抑制促炎细胞因子的合成,具有显著抗炎作用。同时,母乳为婴幼儿时期的最佳营养来源,但目前由于女性健康和工作等原因导致母乳喂养率较低。羊乳和牛乳中蛋白质作为婴幼儿生长发育的必要物质基础和母乳替代品中的重要营养素,目前对其初乳阶段蛋白质分析较少,缺少不同阶段羊乳和牛乳基乳粉的蛋白质精确配比信息。
蛋白质组学分析因其具有鉴别和量化数千种蛋白质的能力,已广泛应用于食品营养特性的深入研究。有研究利用同位素标记相对和绝对定量蛋白质组学技术对牛初乳和牛常乳的乳清蛋白进行差异分析,发现与信号传导相关的9 种差异蛋白。曹雪妍等在牛初乳和牛常乳乳清中鉴定出129 种糖蛋白和190 个差异表达糖基化位点,参与了补体与凝血级联、金黄色葡萄球菌感染和溶酶体等通路。在关中山羊和荷斯坦奶牛的乳清蛋白中鉴定出114 种显著差异表达蛋白,主要参与补体和凝血级联反应。这些结果丰富了牛乳和羊乳的乳清蛋白生物学特性信息,但鲜有用于区分羊乳和牛乳之间差异的分析流程,在小样本快速、可重复和高灵敏分析的情况下基于蛋白质组学分析乳清蛋白差异的方法更少。
为更好地了解羊乳与牛乳的乳清蛋白组成并评价其差异,本研究对羊乳和牛乳在不同泌乳期的乳清蛋白组成进行比较分析。采用非标记定量(lable free quantitative method,LFQ)蛋白质组学方法,基于成本效益和时间最大值算法,系统研究不同泌乳期羊乳和牛乳乳清蛋白质组的组成和变化。利用生物信息学分析进行基因功能注释和参与通路分析,用于评估乳清蛋白质潜在的生物活性功能差异,以期更好地了解牛乳和羊乳的蛋白质组成差异,为羊乳基乳制品的研发提供理论指导。
1 材料与方法
1.1 材料与试剂
羊乳和牛乳样品取自陕西省西安市关中奶山羊和荷斯坦奶牛养殖场。采集产后0~5 d羊初乳和牛初乳和产后1~6 个月的羊常乳和牛常乳样品各10 份,在4 ℃贮藏,并在2~4 h内运送至实验室。分析前混合以消除喂养、胎次及个体差异对初乳和成熟乳乳清蛋白的影响,分为3 份进行重复分析。
蛋白酶抑制剂(生化试剂) 瑞士Roche公司;胰蛋白酶(生化试剂) 美国Promega公司;BCA快速蛋白定量试剂盒 上海碧云天生物技术有限公司;所有分离用有机溶剂均购于美国Sigma-Aldrich公司。
1.2 仪器与设备
EASY-nLC 1000纳升级液相色谱系统串联四极杆-静电场轨道阱(Q-Exactive)高分辨质谱(配有加热电喷雾离子源及Xcalibur 4.1数据处理系统) 美国Thermo Fisher Scientific公司;FD-5真空冷冻干燥机 美国GOLD-SIM公司。
1.3 方法
1.3.1 样品预处理
将1 mmol/L蛋白酶抑制剂分别添加到50 mL生牛乳和50 mL生羊乳中,并于4 ℃、12 000 r/min离心40 min,获得乳脂肪层、乳清层和固体颗粒。在乳清层加入10%乙酸溶液调节pH值至4.6,在4 ℃、12 000 r/min离心30 min去除酪蛋白,收集上层清液即为乳清蛋白。使用BCA快速蛋白定量试剂盒分别测定羊乳和牛乳的乳清蛋白含量。
1.3.2 超滤辅助样品制备消化
将10 μL 100 mmol/L二硫苏糖醇溶液加入至100 μL蛋白样品溶液中,56 ℃孵育1 h,以保持蛋白质巯基处于还原状态,避免蛋白质中半胱氨酸之间形成二硫键。冷却至室温后,加入终浓度50 mmol/L碘乙酰胺,室温下避光孵育40 min使巯基烷基化。然后用100 μL UA缓冲液洗涤乳清样品两次,50 mmol/L NHHCO溶液洗涤一次。将加有胰蛋白酶的NHHCO溶液加入10 kDa超滤离心管,酶与底物质量比为1∶20,37 ℃过夜酶解。用体积分数1%甲酸溶液终止消化后使用C柱进行脱盐,冻干,最终复溶在40 μL体积分数0.1%甲酸溶液中进行质谱鉴定。
1.3.3 液相色谱-质谱鉴定及数据分析
色谱条件:将5 μL待测样品溶液注入C反相色谱柱(100 mm×75 μm,3 μm)中,以IntelliFlow控制流速为250 nL/min,用流动相A(水+体积分数0.1%甲酸+4 mmol/L甲酸铵)和流动相B(80%乙腈+体积分数0.1%甲酸+4 mmol/L甲酸铵)进行样品洗脱。洗脱梯度:0~50 min,95%~65% A、5%~35% B;50~95 min,65%~0% A、35%~100% B;95~100 min,0% A、100% B;100~110.1 min,0%~95% A、100%~5% B;110.1~120 min,95% A、5% B。
质谱条件:加热电喷雾电离离子源,喷雾电压3.8 kV,RF透镜电压55 V,毛细管温度275 ℃,数据依赖采集模式dd/MS,范围/300~1 700,正离子模式下一级扫描分辨率70 000(半峰宽/200),二级扫描分辨率17 500,最大离子注入时间120 ms,在线性离子阱中对前20 个响应最高的前体离子进行碎裂,隔离窗口/2,碰撞能量27 eV。前体离子动态排除时间40 s。
1.4 数据分析
质谱原始数据由MaxQuant软件处理,在选定胰蛋白酶作为消化酶,设置N-端蛋白质乙酰基和氧化为可变修饰、脲甲基化修饰为固定修饰、LFQ定量算法、匹配时间窗口0.7 min和对齐时间窗口20 min等参数,生成蛋白质的定量数据矩阵。SIMCA 14.1软件建立偏最小二乘判别分析(partial least squares-discriminant analysis,PLS-DA)模型增强聚类效果并识别潜在的生物标志物。利用DAVID生物信息学资源6.8进行基因功能富集分析。
2 结果与分析
2.1 蛋白质谱鉴定及表达差异分析
由于质谱仪鉴定肽段具有一定的长度限制,胰蛋白酶产生酶切肽段的C-末端与具有高度特异性的精氨酸和赖氨酸结合,产生长度适当的碱性肽。图1为鉴定肽段的长度分布图,显示鉴定的肽段包含氨基酸数目大部分为7~20,均在肽段长度合理范围内,且与质谱鉴定的分子质量范围(/300~1 700)一致。肽段的精确鉴定可显著降低假阳性结果出现概率。本实验使用的Q-Orbitrap质谱仪一级和二级扫描的质量精确度都小于5×10,但为提高鉴定结果的精确性,在Uniprot数据库搜索肽段匹配误差阈值设置为0.05 Da。所有已鉴定肽的质量误差在±5×10范围内,证明所建立方法的高可靠性和准确性。本研究中,通过肽段在牛乳和羊乳乳清中共鉴定出489 种乳清蛋白,乳清样品中已鉴定蛋白质的LFQ强度跨越近5 个数量级。
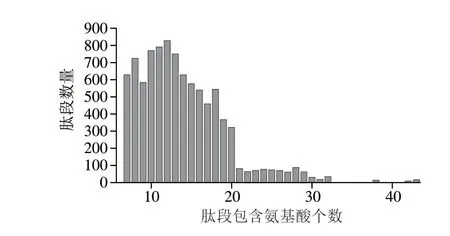
图1 已鉴定肽段的长度分布Fig.1 Length distribution of identified peptides
对所有已鉴定蛋白质进行Venn分析,分别在牛初乳、牛常乳、羊初乳和羊常乳中鉴定到170、161、240 种和291 种乳清蛋白,其中特征蛋白分别为34、45、85 种和97 种。4 种乳清蛋白质组中47 种乳清蛋白共同表达,包括血凝蛋白、补体成分3和血清白蛋白等。牛常乳为目前应用最广泛的乳源,与羊初乳和牛初乳相比,牛常乳中鉴定出57 种特征乳清蛋白;与羊常乳相比,牛常乳中鉴定出85 种特征乳清蛋白,包括血清淀粉样蛋白、-HS-糖蛋白和不溶性弹性结构蛋白等。同时,羊常乳为目前替代牛乳最合适且营养丰富的乳源,与羊初乳和牛初乳相比,羊常乳乳清蛋白中鉴定出109 种特征乳清蛋白。与牛常乳相比,羊常乳中鉴定出110 种特征乳清蛋白,包括-2-糖蛋白1、MAP34-A蛋白和骨桥蛋白等。
2.2 不同泌乳期羊乳和牛乳乳清蛋白化学计量学分析
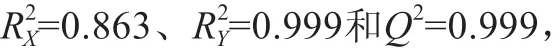

图2 4种乳清样品的PLS-DA模型Fig.2 PLS-DA model for four whey samples
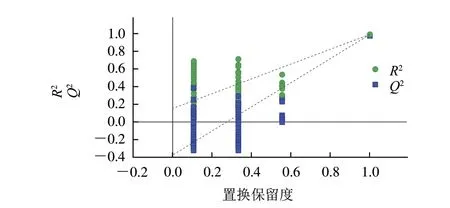
图3 PLS-DA模型置换检验图Fig.3 Permutation test plot for validation of PLS-DA model
为确定4 组乳清蛋白质组间的生物标志物进行变量投影重要性(variable important in projection,VIP)分析。由图4可知,组织蛋白酶2、白细胞介素20受体亚单位、PKS-ER结构域蛋白、纤维蛋白原链、膜联蛋白、核结合蛋白2、黑素转铁蛋白、乳脂肪球表皮生长因子8蛋白和血小板反应蛋白1这9 种蛋白VIP值大于1,且在羊乳中含量较高,可用于区分牛初乳、牛常乳、羊初乳和羊常乳。膜联蛋白是一种钙依赖性磷脂结合蛋白,参与细胞膜和细胞骨架的形成,具有稳定脂质双层膜骨架作用。同时,膜联蛋白可通过与甲酰肽受体结合,抑制中性粒细胞滚动,减少中性粒细胞黏附和渗出,从而减少炎症部位中性粒细胞浸润和炎性细胞因子释放。纤维蛋白原是肝脏合成凝血系统的核心蛋白质,在凝血酶作用下形成凝固性纤维蛋白凝块发挥凝血作用,而纤维蛋白原链的翻译和转录是合成整个纤维蛋白原分子的限速步骤。CD5分子样蛋白、丝氨酸蛋白酶抑制剂A3-5、线粒体ATP合成酶亚单位、桥粒蛋白、含FABP结构域的蛋白质和内质网伴侣蛋白在其他3 组乳中含量较高。其中桥粒蛋白在牛初乳中含量最高,为细胞内桥粒结构的基本组成部分,可通过影响钠离子通道功能属性或Cx43蛋白表达分布导致心律失常。本实验结果表明,与羊乳相比,桥粒蛋白在牛乳清中表达更为广泛。

图4 4 组乳清蛋白质组间生物标志物VIP得分Fig.4 VIP scores of biomarkers among whey proteomes from bovine and goat colostrum and mature milk
2.3 不同泌乳期羊乳和牛乳乳清蛋白生物信息学分析
利用DAVID生物信息学资源6.8,根据分子功能、细胞成分和生物过程对牛乳和羊乳中的乳清蛋白进行分类。胞外区、胞外间隙、血液微粒和胞外体是乳清蛋白定位的主要细胞成分,说明这些低丰度蛋白组分是血清中蛋白质在乳腺细胞的紧密连接处通过细胞间或跨细胞传递到乳清中。其他蛋白与角蛋白丝、局灶性黏连、血小板颗粒等功能有关。乳清蛋白的主要分子功能有结构分子活性、丝氨酸型内肽酶抑制剂活性以及糖蛋白结合。丝氨酸型肽链内切酶抑制剂活性、丝氨酸型肽链内切酶活性和结构分子活性在羊常乳中最为丰富,其次是半胱氨酸型肽链内切酶抑制剂活性、糖蛋白结合和过氧化还原酶活性。因此,根据蛋白质组学的基因功能富集分析,乳清蛋白可能参与了人类健康相关途径,而其生物活性在羊乳和牛乳的初乳和常乳中有所不同。
乳清蛋白参与的生物学过程以视网膜稳态和内肽酶活性负调控最为显著,急性期反应、纤溶和糖酵解过程也占很大比例。羊常乳中,与视网膜稳态、纤溶、先天免疫应答有关的蛋白质数量明显较多;牛常乳中,与抗菌体液反应相关的蛋白质数量显著增加。参与糖酵解过程、肽激素分泌的阳性调节和血管收缩阳性调节的蛋白质在羊初乳中表达较高;参与内肽酶活性负调控、急性期反应、抗原处理和通过MHC I类表达肽抗原以及免疫应答的蛋白质在牛初乳中表达较高。进一步研究羊初乳和牛初乳的乳清蛋白差异发现,有170 种蛋白丰度存在显著差异。牛初乳中有136 种乳清蛋白未被鉴定或含量极低,包括-烯醇化酶、血管紧张素原和补体成分3等。羊初乳中的高表达蛋白与补体激活、内肽酶活性负调节、炎症反应、细胞迁移、脂质储存和生物素代谢过程有关;这些特征蛋白的分子功能包括磷脂酰化酶活性、磷脂结合、糖蛋白结合和丝氨酸型肽链内切酶抑制剂活性。此外,肌抑制素、凝溶胶蛋白、内质网伴侣BiP、桥粒胶蛋白、黏着素亚单位SA-1等34 种常见蛋白在羊初乳中的丰度明显高于牛初乳,这些常见蛋白主要与免疫应答、内肽酶活性负调控、视网膜稳态、细胞黏附、骨化和急性期反应相关。这些蛋白在羊初乳中的分子功能包括丝氨酸型内肽酶活性、肝素结合、钙离子结合、细胞外基质结合和蛋白结合。说明初乳中高水平的抗菌蛋白和免疫活性蛋白可能有助于消除肠道中的致病菌,减少肠道炎症和败血症。羊初乳中急性期反应蛋白的丰度高于牛初乳,可能是由于哺乳初期初生羔羊面临从无菌环境到微生物环境的胁迫所致。初乳是帮助新生儿建立自身免疫系统的主要营养来源,羊初乳中较高丰度的急性期反应蛋白有助于新生儿建立抗微生物感染的免疫系统。
与其他乳清相比,羊常乳中乳清蛋白共有107 个蛋白丰度存在显著差异,其中54 种蛋白在羊常乳中特异性表达,包括肌抑制素、中性粒细胞明胶酶相关脂蛋白、凝溶胶蛋白、血凝蛋白、尿苷磷酸化酶2、蛋白S100-A8、黄嘌呤脱氢酶等。羊常乳中,13 种蛋白质丰度显著高于其他乳,包括肌抑制素、内质网伴侣BiP、肌球蛋白7和前纤维蛋白1等;11 种蛋白质丰度极低或未在羊常乳中鉴定,包括CD5分子样蛋白、角苔藓素、纤维连接蛋白、乳胶蛋白和核结合蛋白-1等。值得注意的是,羊常乳中的96 种高表达蛋白中有17 种与免疫应答有关,包括血凝蛋白、中性粒细胞明胶酶相关脂蛋白、骨桥蛋白、补体成分3和S100-A9蛋白等。骨桥蛋白是一种由破骨细胞、成骨细胞、肾细胞和内皮细胞以及活化免疫细胞(如T细胞和巨噬细胞)分泌的磷酸化糖蛋白,参与骨形态发生和调节炎症细胞动员,在炎症和组织重塑部位显著上调,可作为促炎和抗炎细胞因子。中性粒细胞明胶酶相关脂蛋白是一种分子质量25 kDa的分泌糖蛋白,在细菌感染时由先天免疫细胞分泌产生,可通过螯合载铁分子限制细菌的生长速率。这些结果都表明羊常乳可以有效提高机体免疫力。
黄嘌呤脱氢酶、肌球蛋白7、U6-snRNA相关的Sm样蛋白LSm4、-乳清蛋白和载脂蛋白A1等17 种羊常乳中高表达的蛋白质与一系列代谢过程有关。在哺乳动物中黄嘌呤脱氢酶和黄嘌呤氧化酶是黄嘌呤氧化还原酶的两种互换形式。黄嘌呤脱氢酶为其天然形式,主要存在于细胞内部,可通过巯基氧化或有限的蛋白水解作用转化为黄嘌呤氧化酶。在黄嘌呤脱氢酶到黄嘌呤氧化酶的转化过程中,具有NAD依赖性脱氢酶和O依赖性氧化酶活性的中间产物可能产生HO和超氧阴离子自由基等活性氧,在过渡金属存在下通过Haber-Weiss和Fenton反应生成高细胞毒性的羟自由基。黄嘌呤脱氢酶通过产生影响氧化还原平衡的氧化剂参与代谢过程。载脂蛋白A1是高密度脂蛋白的载体蛋白,将胆固醇从周围组织运输到肝脏代谢,从而清洁血管壁。血清高密度脂蛋白胆固醇水平越高,人体将血清胆固醇转运到肝脏代谢的能力越好,从而有效降低动脉粥样硬化发生的可能性。而载脂蛋白A1水平的升高会增加高密度脂蛋白的合成,因此载脂蛋白A1可通过参与代谢过程降低罹患心血管疾病的风险。
5 种蛋白(内质网伴侣BiP、凝溶胶蛋白、转录因子AP-2、双特异性蛋白磷酸酶和rho-GDP解离抑制剂2)参与了细胞凋亡过程。内质网伴侣BiP是多功能蛋白质,其能与靠近内质网的免疫球蛋白结合蛋白、凝集素钙蛋白和钙网蛋白的多肽产生相互作用。对内质网和线粒体Ca串扰的调节是其附加功能之一,这种功能依赖于内质网伴侣BiP与内质网Ca处理机制的相互作用,这种新的附加功能允许内质网伴侣BiP直接控制线粒体凋亡,无需通过未折叠蛋白反应上调促凋亡转录因子。内质网蛋白质折叠机制通过控制线粒体氧化磷酸化速率微调ATP的输入。内质网伴侣BiP在调节内质网线粒体Ca流量中的作用表明,在分泌大量蛋白质的细胞类型中,分泌蛋白折叠的过程是细胞存活和死亡的中心调节器;而在其他类型细胞中,内质网蛋白折叠可能是一种前哨机制,监测细胞的健康状况,以控制细胞代谢和凋亡。凝溶胶蛋白是一种内源性细胞质蛋白和分泌性血浆蛋白,可以调节肌动蛋白组装和分解,其裂解产物在多种细胞类型中的表达导致细胞分离、聚集和核碎裂。已有报道通过HEK293T细胞的高转染效率研究电压依赖性阴离子通道(voltage-dependent anion-selective channel,VDAC)蛋白与凝溶胶蛋白之间的相互作用,并证明VDAC与凝溶胶蛋白的G5结构域结合,而不与其他结构域结合。凝溶胶蛋白的G5段显示VDAC与HIV-Vpr重叠、竞争性结合,G5结构域通过阻断Vpr与VDAC的结合抑制HIV-Vpr诱导的T细胞凋亡。双特异性蛋白磷酸酶通过差异表达和对MAPK信号通路的调节在多种类型人类癌症中发挥作用,其特异性抑制肿瘤坏死因子诱导的p38/MAPK活化,促进细胞凋亡,降低核因子(nuclear factor,NF-)活性。同时双特异性蛋白磷酸酶在DU145细胞中的过度表达也会通过阻断p65/NF-核移位促进细胞凋亡和降低NF-活性,从而抑制癌症发生或延缓发病。
羊乳和牛乳的初乳和常乳中蛋白质显著表达的前10 条通路如图5所示。补体与凝血级联、抗生素生物合成和碳代谢是最显著的途径。-抗蛋白酶、补体成分3和补体因子H参与补体与凝血级联途径。丙酮酸激酶、-烯醇化酶、核苷二磷酸激酶和补体因子H与抗生素的生物合成有关。PALP结构域蛋白、葡萄糖-6-磷酸异构酶和异柠檬酸脱氢酶参与碳代谢。其他主要途径有Rap1信号途径、氨基酸生物合成和溶酶体。根据通路分析,实验鉴定的乳清蛋白与多种代谢途径有关。在乳清蛋白中,补体与凝血级联途径是天然免疫的介质,可产生具有促炎症特性的补体裂变产物。在病理生理学中,凝血级联激活后损伤部位凝血酶的形成可以提供抵御微生物入侵的物理屏障,并触发补体系统。此外,通过局部刺激产生的过敏毒素可激活细胞免疫应答。Amara等分析体内和体外凝血系统中多种因素与中枢补体成分之间的多重联系。本实验中,羊乳清中补体裂变产物的含量高于牛乳,而其中补体因子H有助于胎儿免疫保护,补体成分3和补体成分7可作为防御因子。羊乳和牛乳中乳清蛋白对糖代谢的影响可能与其活性乳清蛋白有关。葡萄糖-6-磷酸异构酶是一种胞浆型非限速酶,能支持特定胚胎脊髓神经元的存活和培养,并介导HL-60细胞分化为终末单核细胞。异柠檬酸脱氢酶作为一种新的角膜晶体蛋白,在促进角膜透明性方面发挥结构和催化作用。Nam等报道随异柠檬酸脱氢酶表达的减少,3T3-L1前脂肪细胞的脂肪生成减少,葡萄糖摄取和脂肪合成减少。此外,异柠檬酸脱氢酶表达的抑制导致NOX4活性抑制,从而降低分化脂肪细胞的水平。羊乳清中PALP结构域蛋白、葡萄糖-6-磷酸异构酶和异柠檬酸脱氢酶的含量越高,羊乳的降血糖作用越强。

图5 不同泌乳期羊乳和牛乳蛋白质富集的通路分析Fig.5 KEGG pathway enrichment analysis of proteins in goat and bovine colostrum and mature milk from
3 结论
在牛初乳、牛常乳、羊初乳和羊常乳中分别鉴定到170、161、240 种和291 种乳清蛋白,47 种蛋白质在4 组乳样中共同表达,说明虽然羊乳和牛乳之间存在显著差异,但羊乳在一定程度上可用于替代牛乳。对所鉴定乳清蛋白的LFQ强度建立PLS-DA模型,发现牛初乳和牛常乳在蛋白质组成上比羊初乳和羊常乳更相似,筛选了羊乳中丰度较高的9 种蛋白质用作标志物区分这4 组乳样。经生物信息学分析发现,鉴定的乳清蛋白主要参与的生物过程为视网膜稳态和内肽酶活性负调控,定位的主要细胞成分是胞外区、胞外间隙、血液微粒和胞外体,主要分子功能为结构分子活性、丝氨酸型内肽酶抑制剂活性和糖蛋白结合。通路分析发现,补体与凝血级联、抗生素生物合成和碳代谢是最显著的途径。在羊常乳中筛选出96 种高丰度蛋白质,大部分与免疫应答、代谢过程及细胞凋亡有关。羊初乳中34 种常见蛋白的丰度明显高于牛初乳,主要与免疫应答、内肽酶活性负调控和急性期反应相关。研究结果发现羊乳的特征蛋白组成及差异蛋白可能参与的生物过程,加深对羊乳蛋白的认识,对牛乳及其制品的营养改良和母乳替代品的生产具有重要意义。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30 08:22:06
自我保健(2020年8期)2020-01-01 21:12:03
食品安全导刊(2019年36期)2019-01-04 16:20:36
食品工业科技(2014年15期)2014-03-11 18:17:43
食品工业科技(2014年5期)2014-03-11 18:14:08
食品科学(2013年17期)2013-03-11 18:26:49
食品科学(2013年15期)2013-03-11 18:25:41
食品科学(2013年6期)2013-03-11 18:20:09
健康必读(2012年10期)2012-04-29 00:44:03
小小说大世界(2009年1期)2009-04-03 09:48:46
