
小麦苗期铅耐受性的全基因组关联分析
2022-05-16 08:49职蕾者理孙楠楠杨阳DaurenSerikbay贾汉忠胡银岗陈亮
中国农业科学 2022年6期
职蕾,者理,孙楠楠,杨阳,Dauren Serikbay,2,贾汉忠,胡银岗,陈亮
小麦苗期铅耐受性的全基因组关联分析
职蕾1,者理1,孙楠楠1,杨阳1,Dauren Serikbay1,2,贾汉忠3,胡银岗1,陈亮1*
1西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,中国陕西杨凌 712100;2赛富林农业技术大学,哈萨克斯坦努尔苏丹 010011;3西北农林科技大学资源与环境学院,中国陕西杨凌 712100
【目的】随着工业化的推进,重金属尤其是铅对耕地的污染已成为世界性问题。小麦作为主要粮食作物,其健康生产对保障粮食安全意义重大,筛选铅耐受性强和铅低积累小麦品种、挖掘相关调控基因或QTL区间,为耐铅种质创新和揭示小麦铅耐受性遗传机制奠定基础。【方法】采用140 mg·kg-1的硝酸铅溶液对102份小麦品种(系)进行苗期胁迫试验,以3个重复下的最大根长、根生物量和生长速率的耐铅系数的加权隶属函数值(值)来评价小麦对铅的耐受性。结合小麦660K SNP芯片的335 438个高质量SNP标记对小麦铅耐受性进行全基因组关联分析(genome-wide association study,GWAS),挖掘铅耐受性候选基因。【结果】小麦品种(系)之间的铅耐受性表现出丰富的变异,变异系数为44.8%—46.2%,相关系数介于0.87—0.97(<0.001);铅耐受性强的品种呈现出铅低积累特性。基因分型结果显示SNP多态性信息含量(polymorphic information content,)为0.28—0.32,群体结构分析将供试材料分为7个亚群;2种GWAS分析方法共检测到20个与小麦铅耐受性显著关联的SNP(≤0.001)和8个候选区间,分别分布在1B、2A、2D、3A、3B、5A和7A染色体上,单个位点可解释15.33%—19.75%的表型变异,其中10个位点和8个候选区间在2个及以上环境被检测到。分析稳定检测的显著关联位点及区间的候选基因,发现其功能主要与跨膜运输、蛋白修饰以及氧化应激反应有关,包括7个与转运蛋白相关基因(、、等)、2个与泛素化与去泛素化相关的候选基因(和)、3个跨膜蛋白基因(、和)和1个过氧化物酶相关的候选基因()。【结论】筛选出铅耐受性强的种质材料7份,检测到与小麦铅耐受性显著关联的20个SNP位点及8个候选区间,筛选出13个与小麦铅耐受性相关的候选基因。
小麦;铅耐受性;全基因组关联分析;候选基因分析
0 引言
【研究意义】在矿石开采和能源利用过程中,重金属会随着以矿渣、粉尘、废水等污染介质进入环境,并在大气、水体、土壤和植物中迁移,造成严重的面源污染及重金属污染[1]。每年由于重金属污染造成的粮食产量损失达1 000万t以上,导致直接经济损失超200亿元[2-3]。农业面源污染及重金属污染已成为保障粮食安全亟待解决的问题,国家科技计划也高度重视农业面源及重金属污染的综合防治[4]。铅作为土壤重金属污染的主要元素之一,对动植物及人体的生理、生化等多方面均有消极影响[5-6]。铅污染耕地、空气、水等已引发多起群体血铅中毒事件[7]。环境保护部和国土资源部2014年发布的《全国土壤污染状况调查公报》显示,中国农田土壤污染点位超标率为19.4%,其中,铅是土壤污染中风险较高的金属元素之一,呈现从西北到东南、从东北到西南逐渐升高的态势。铅对小麦的营养和生殖生长均存在不同程度的抑制作用[8-9],而且部分小麦产区已面临严重的铅污染,给小麦生产带来了新的挑战;配合耕地土壤污染修复,在中、轻度污染耕地种植耐受性强、低积累的小麦品种是保障耕地效益和生产优质粮食的理想途径。因此,为保障污染农田的健康生产,筛选耐铅的小麦种质资源、挖掘铅积累相关遗传位点十分必要[10]。【前人研究进展】根据重金属在植物生态系统中的作用,可以将其分为必需金属和非必需金属。铜、铁、锰、镍和锌等属于必需金属,适量的必需金属对植物生长发育有促进作用。而铅、镉、砷、汞等非必需金属在植物中没有已知功能,且对植物生长发育有毒害作用。在拟南芥中,、、、可以激活ATP结合盒(ATP-binding cassette,ABC)转运蛋白的表达[11-13],从而增强植物对铅的耐受性;编码含有酰基辅酶A结合域的蛋白质,参与植物铅耐受性调控[14]。在烟草和大麦中,NtCBP4(质膜钙调蛋白结合转运蛋白)和HvCBT1(Hordeun vulgare CaM结合蛋白)编码的离子转运蛋白通过调控阳离子的非选择性运输,从而使植株表现出对重金属的耐受性[15-16]。全基因组关联分析是一种检测复杂性状遗传因素的有效分析方法,已广泛用于水稻、小麦、玉米等作物重要性状调控基因的挖掘[17-19],并在植物镉耐受性[20-21]以及油菜耐铅分子机制研究[22]等方面取得了重大进展。【本研究切入点】铅抑制植物的生长发育,最显著的是抑制根系的生长[23-24],目前,水稻、油菜及柳树的耐铅基因型被挖掘应用[25-27],但鲜有耐铅小麦种质筛选及其分子机制解析相关报道。【拟解决的关键问题】本研究参照植物盐胁迫中采用耐盐系数及加权隶属函数值对耐盐性进行评价的方法[28-30],依据根部2个性状和地上部生长速率等3个性状的耐铅系数及加权隶属函数值综合评价小麦对铅的耐受性,结合660K SNP芯片进行基因型分析,通过GWAS分析挖掘小麦耐铅的调控位点及候选基因。通过筛选小麦耐铅种质,挖掘小麦铅耐受性相关遗传位点及候选基因,为小麦耐铅种质创新及机理解析奠定基础。
1 材料与方法
1.1 材料
选用102份小麦品种(系),其中42份来自河南省(40.19%),21份来自陕西省(21.57%),9份来自山东省(9.80%),其余小麦品种(系)来自山西省、江苏省、安徽省、宁夏自治区、河北省、甘肃省、北京市及国外(电子附表1)。
1.2 试验方法及指标测定
1.2.1 试验方法 供试材料于2020年8—10月,在西北农林科技大学旱区作物逆境生物学国家重点实验室光照培养间进行水培试验。分为空白对照组和铅胁迫组,空白对照组采用纯水+1/2浓度霍格兰营养液,铅胁迫组培养液采用浓度为140 mg·kg-1Pb(NO3)2+1/2浓度霍格兰营养液[31]。培养方法:将种子用75%乙醇消毒5 min,再用纯水清洗干净后置于铺有滤纸的培养皿(直径为9 cm)上,每组处理30粒种子,2组处理分别加入纯水和140 mg·kg-1Pb(NO3)2进行萌发,3 d后挑选长出第一片叶且长势一致的10株幼苗移入打好孔的泡沫板,进行水培。由于正常浓度的霍格兰营养液与硝酸铅反应产生沉淀,降低了铅离子胁迫浓度,所以本研究采取1/2浓度霍格兰营养液进行水培处理:空白对照组采用纯水进行培养,铅胁迫组采用140 mg·kg-1Pb(NO3)2进行培养,每个培养盒8个品种,加入3 L培养液,培养液每3天更换一次,培养一周后,将空白对照组的培养液换成1/2浓度霍格兰营养液,铅胁迫处理的培养液换成含有140 mg·kg-1Pb(NO3)2的1/2浓度霍格兰营养液。继续培养至三叶期,收获测量性状。设3次重复试验。
1.2.2 测定指标及测定方法 在萌发后的第3天及第9天挑选长势一致的5株小麦,测定其地上部基部到顶部的长度作为苗高,以苗高差与对应天数的比值作为生长速率。
水培至三叶期,收获长势均匀一致的5株小麦,采用直接测量法测定其最大根长,将根系全部剪下,80℃烘干至恒重,用万分之一天平测定根系干物质重量,取平均值计算每株小麦的根生物量。
1.3 表型数据分析
利用Excel整理最大根长、根生物量、地上部生长速率的观测值,并分别计算耐铅系数[32],利用隶属函数法将各材料3个指标的耐铅系数分别转换为隶属函数值,以某个指标的变异系数在所有指标变异系数中所占的比率为该指标耐盐系数的权重,进而计算各材料在各重复下的加权隶属函数值(值)[23]。
耐铅系数=铅胁迫测定值/对照处理测定值×100%
加权隶属函数值(值)的计算公式如下:
(=1,2,3,…,)
=(=1,2,3,…,)
式中,X为各小麦品种(系)基于测定指标的耐铅系数,Xmax、Xmin分别为供试材料中X的最大值和最小值,(X)为各供试材料X的隶属函数值,CV为各供试材料(X)为的变异系数,W表示CV在总变异中所占的比率。
用R包Lme4计算3次重复值的最优线性无偏性值(best linear unbiased estimate,BLUE)。
广义遗传力计算公式如下:
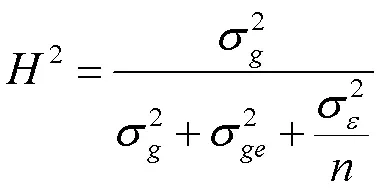
其中,为遗传方差,为基因型与环境互作方差,为环境方差,为环境数。
1.4 重金属含量的测定
自来水冲洗样品,洗去根系分泌物等,再用去离子水冲洗,沥干水分,80℃烘至恒重。对样品的地上部与根部分别进行微波消解(Multiprep-41FC2):称取0.2 g(不足0.2 g则称取全部),加入6 ml硝酸(HNO3),微波消解梯度升温进行消解。消解完成后,用1%硝酸定容到10 ml。样品中的铅(Pb)含量采用火焰/石墨烯原子吸收光谱仪(日立Z-2000)进行测定。
1.5 群体结构与连锁不平衡分析
使用小麦660K SNP芯片对参试小麦材料进行基因分型,对分型结果进行人工质量控制,剔除数据缺失率>50%,杂合率大于20%和最小等位基因频率<0.1的SNP标记,共获得335 438个高质量SNP标记。采用Power Marker V3.25软件分析参试材料的遗传多样性和多态性信息含量(polymorphic information content,),1-(P)2(P表示位点的第个等位变异出现的频率)。从筛选的标记中,随机选取2 000个最小等位基因频率大于10%,且在染色体上均匀分布的SNP,利用Structure 2.3.4软件进行群体结构分析;参数设置为:Length of Burn-Period=10 000,MCMC Reps after Burn-in=100 000,选择Admixture Ancestry模型和Dependent Allele Frequencies模式,令K=2—12,每个K值重复5次。以位点间的相关系数平方(2)衡量多态性位点两两之间的连锁不平衡(linkage disequilibrium,LD),使用PopLDdecay软件进行A、B和D基因组的全基因组连锁不平衡分析,计算2参数设置为:-MaxDist 30000[33-34];LD衰减分析,将覆盖SNP对的30 000 kb LD区间划分为100 kb的区间,以第95百分位的2值作为阈值估测LD衰减距离[35]。
1.6 全基因组关联分析及候选基因筛选
基于筛选到的高质量SNP标记,利用Tassel 5.0软件中的Q+K混合线性模型进行全基因组关联分析,显著关联阈值估计为-log10()=5.5(=1/N,其中N=使用的SNP数量)。此外,为了避免忽略次要位点的影响,还选择阈值为3进行后续分析。判定SNP标记与目标性状关联的显著性,利用R语言绘制关联分析结果的曼哈顿图和Q-Q图[22, 25]。进一步利用FarmCPU(fixed and random model circulating probability unification,http://zzlab.net/FarmCPU/index.html)对Tassel的Q+K混合线性模型的分析结果进行检验。利用多种环境下检测到的与铅耐受性显著关联的SNP对供试材料进行基因分型,将供试材料分为2类(仅选择纯合基因型),并使用SPSS 25的检验分析基因型对表型的效应。使用R包ggplot2绘制箱线图。利用中国春基因组数据库(https://wheat-urgi.versailles.inra.fr/)获取与小麦铅胁迫显著关联的SNP标记序列,在NCBI数据库中进行BLASTx序列比对[36],根据从Ensemble plants数据库(http://plants.ensembl.org/index.html)下载基因注释信息,对QTL物理区间内所包含的基因进行注释,辅助候选基因筛选。
2 结果
2.1 小麦耐铅种质筛选
比较102份小麦材料在0和140 mg·kg-1铅胁迫下生长至三叶期的最大根长、根生物量和生长速率(图1-A—图1-C)。正常处理下的最大根长为4.83—31.87 cm,铅胁迫处理下的最大根长为1.70—9.87 cm,99.9%小麦材料的最大根长受到抑制,遗传力为7.57%;正常处理和铅胁迫处理下根生物量的范围分别为0.0023—0.0152 g和0.002—0.094 g,68.6%小麦材料的根生物量受到抑制,遗传力为35.54%;正常处理和铅胁迫处理下生长速率分别为0.24—2.24 cm·d-1和0.29—1.49 cm·d-1,90.2%小麦材料的生长速率受到抑制,遗传力为25.62%(图1-D)。以上结果表明,大多数小麦材料的最大根长、根生物量及生长速率在铅胁迫下均受到不同程度的抑制。
使用最大根长耐铅系数、根生物量耐铅系数和生长速率耐铅系数的加权隶属函数值(值)综合评价各个品种的铅耐受性。值范围为0.029—0.342,变异系数范围为44.80%—46.20%,遗传力为88.9%(表1)。表明这些材料的铅耐受性存在广泛的遗传变异,通过相关性分析发现这些材料3次重复间的铅耐受性加权隶属函数值(值)显著相关,相关系数超过0.87(图1-E)。根据各品种的值,选择值处于前七位且大于0.3的7个小麦品种为铅耐受性稳定的小麦品种(表2),这7个品种分别来自河南省、陕西省、山西省、山东省等地,均为21世纪以来育成的品种。
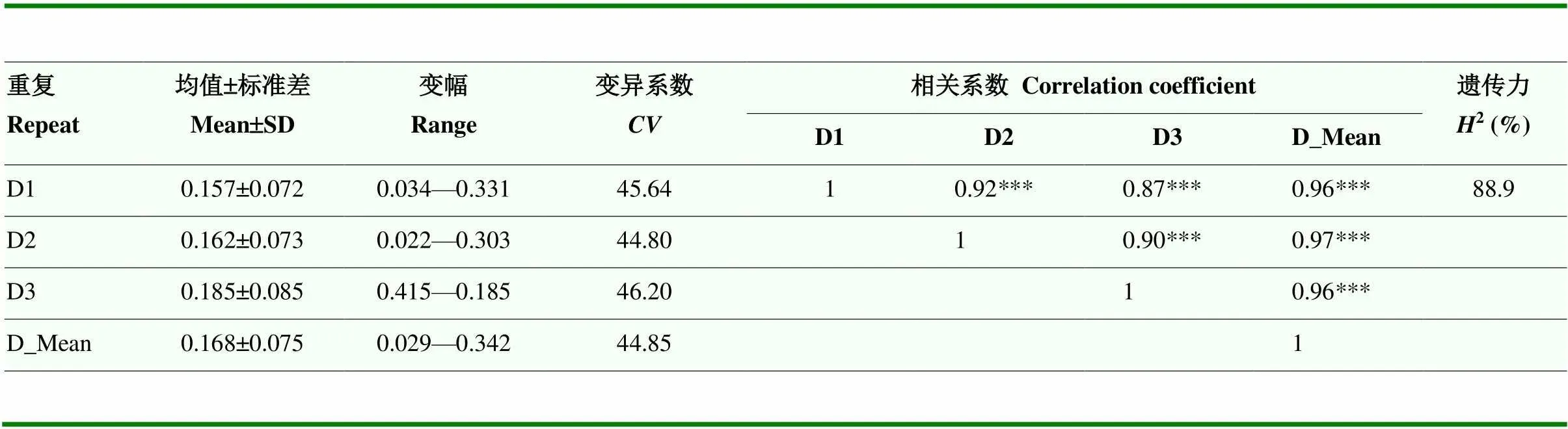
表1 3个重复中102份小麦铅耐受性加权隶属函数值(D值)的统计分析
D1:重复1;D2:重复2;D3:重复3;D_Mean:平均值;SD:标准差;:变异系数;***表示在<0.001水平差异显著。下同
D1: repeat 1; D2: repeat 2; D3: repeat 3; D_Mean: D average; SD: Standard deviation;: Variable coefficient; ***Significant at<0.001.The same as below

A—C:分别为正常处理与铅胁迫下102份小麦的最大根长、生长速率和根生物量的分布;D:铅胁迫对小麦品种(系)的抑制情况;E:最大根长、根生物量和生长速率3个指标的耐铅系数的加权隶属函数值(D值)的频率分布和相关性矩阵,D1、D2、D3、average分别代表重复1、重复2、重复3以及3次重复的平均值。下同
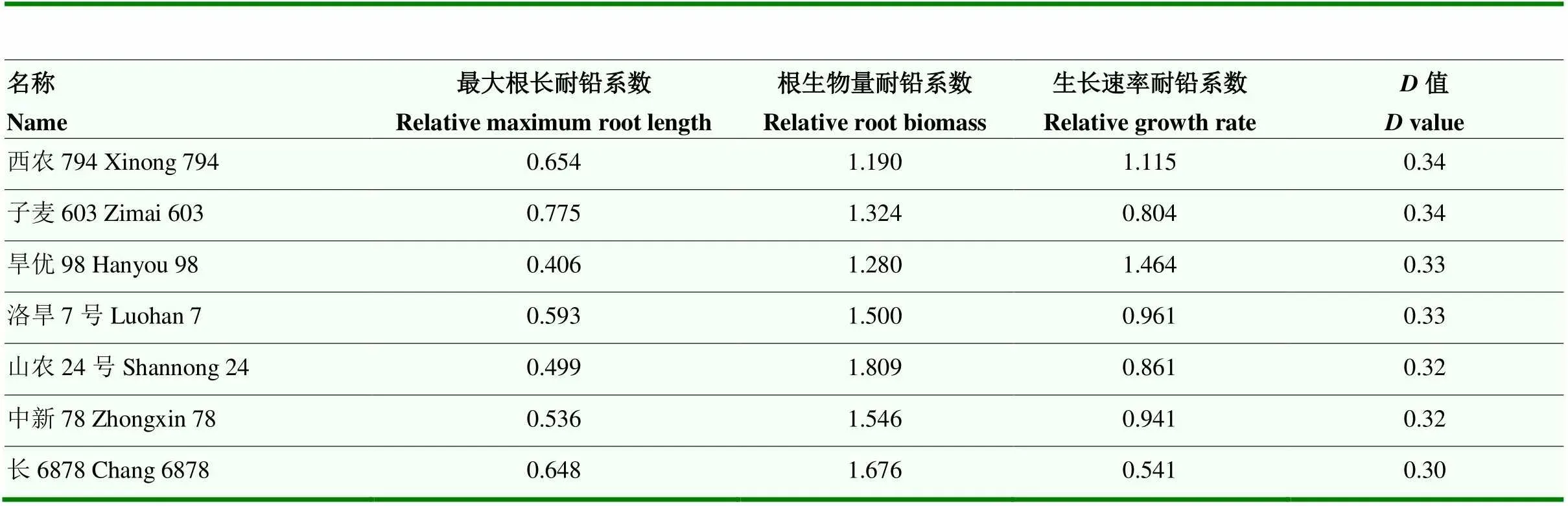
表2 102份小麦材料中筛选出的耐铅性强的种质
取值处于前10位的铅耐受强性的品种和值处于后10位的铅耐受性低的品种,对其地上部与根部的铅含量均值进行独立样本的检验分析(图2),结果表明,对于根部铅含量,小麦铅耐受性强的品种显著低于耐受性低的品种。对于地上部铅含量,耐受性强的品种低于耐受性低的品种,但差异不显著。
2.2 SNP多态性及分布
根据小麦660K SNP芯片的基因分型结果,筛选出供试材料中具有多态性的SNP标记335 438个,分布在A、B和D基因组上的SNP数目分别为124 264(37.05%)、168 168(51.13%)和43 006(12.82%),其中,B基因组的多态性最高,D基因组的多态性远低于A和B(表3)。21条染色体中,3B染色体上SNP标记数目最多(43 683个),4D染色体上的SNP标记数目最少(2 563个)。3个染色体组的表现为A(0.310)>B(0.308)>D(0.305)。
2.3 群体结构与连锁不平衡分析
Structure V2.3.4软件分析结果(图3-A)表明,在K=7时,ΔK值最大,曲线变化程度最大,表明参试材料可分为7个类群(图3-B和图4)。亚群Ⅰ和亚群Ⅴ分别包含6和23个品种(系),主要来自陕西省,亚群Ⅰ中包含少量甘肃省品种,亚群Ⅴ中包含少量河北省、宁夏自治区、江苏省的品种;亚群Ⅱ包含10个品种(系),来自陕西省和河南省;亚群Ⅲ和亚群Ⅳ分别包含26和12个品种(系),主要来自河南省;亚群Ⅵ包含13个品种,主要来自山东省;亚群Ⅶ包含12个品种(系),来自河南省、安徽省和山东省。基于PopLDdecay对102份小麦品种(系)LD衰减距离计算的结果及前人对小麦中全基因组LD衰减距离的分析[28, 37],将物理图谱上前后5 Mb区间内的位点认定为一个候选位点。
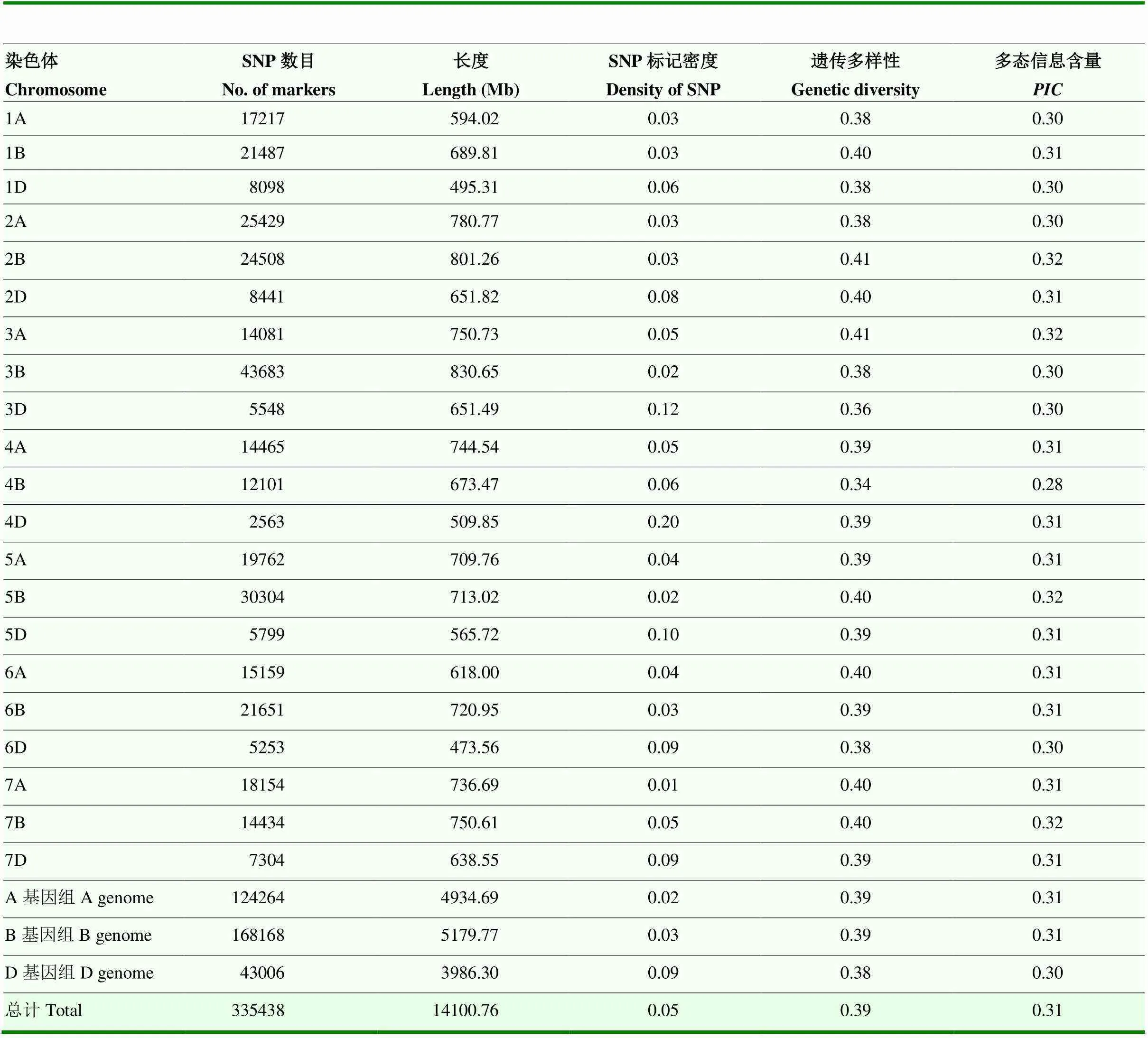
表3 SNP标记的分布、物理图谱长度及其多态性
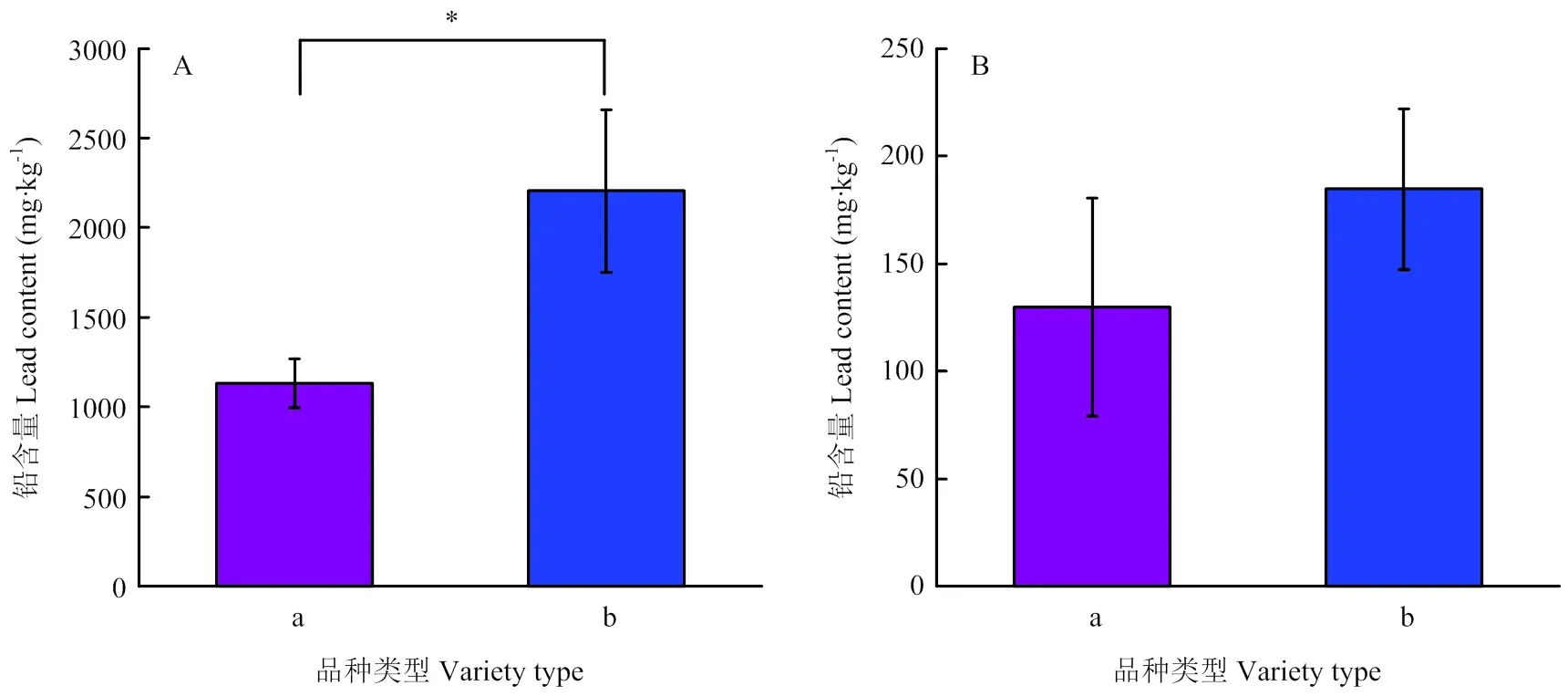
A:根部铅含量;B:地上部铅含量;a:耐受性强品种;b:耐受性低品种;*表示差异显著(P<0.05)
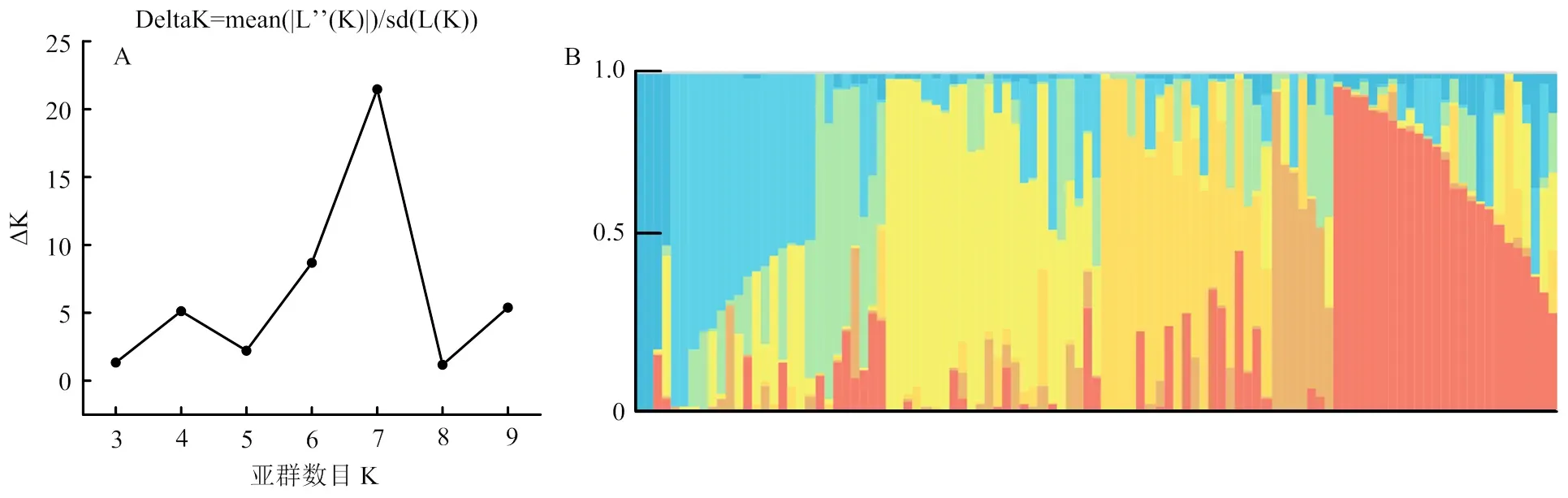
A:群体的∆K值;B:群体结构示意图 A: Estimation of ∆K value in population; B: Group structure
2.4 耐铅相关QTL的检测
以102份小麦品种(系)3个重复及BLUE的铅耐受性评价值(值)和660K SNP芯片检测到的335 438个SNP分别进行全基因组关联分析,采用Q+K混合线性模型,重复D1中检测到14个显著关联位点,分别位于2A、2D、3A、3B、5B、7A等6条染色体上,贡献率为15.61%—20.89%;重复D2中检测到11个显著关联位点,分别位于2A、2D、3A、3B、7A等5条染色体上,贡献率为15.62%—18.69%;重复D3中检测到6个显著关联位点,分别位于2A、3B、5A等3条染色体上,贡献率为15.47%—19.39%。重复BLUE中检测到9个显著关联位点,分别位于2A、2D、7A等3条染色体上,贡献率为14.97%—19.88%。总之,共检测到20个与小麦铅耐受性显著关联的位点(图5和表4),贡献率为15.33%—20.89%。
为了检测Q+K混合线性模型分析的结果,进一步利用质控后得到的335 438个SNP标记,运用R包GAPIT中的FarmCPU模型对3次重复及BLUP共4个环境下小麦铅耐受性进行全基因组关联分析,结合LD衰减距离,共找到8个与小麦铅耐受性相关QTL区间,包含201个显著关联位点,分别位于1B、2A、2D、3B、5B、7A和7D共7条染色体上(图5和表5)。Q+K混合线性模型分析得到的20个SNP中有13个的物理位置映射到FarmCPU的结果QTL区间内或附近,2种方法间存在良好的一致性。
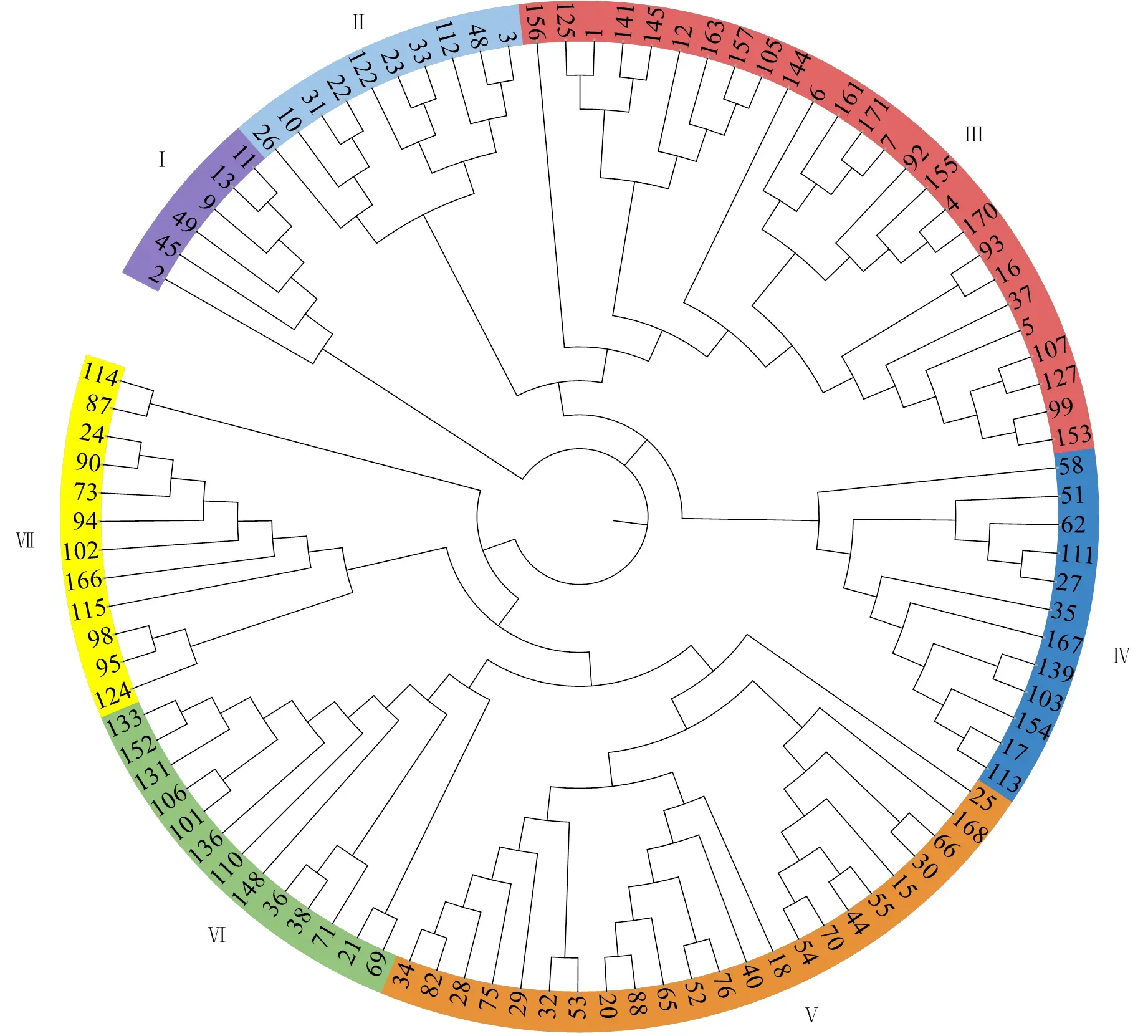
图中编号对应电子附表1中各试验材料的试验编号
2.5 不同基因型表型差异分析
为检验不同基因型对铅耐受性的影响,选取Q+K模型检测到的能够同时在2个及以上的环境中检测到的9个SNP,根据其基因型对群体进行分组,并采用检验分析不同基因型对铅耐受性的效应。结果表明,这9个SNP的2种单倍型间的耐受性值均存在显著或极显著差异(图6)。有5个SNP(AX-109277543、AX-108894641、AX-108776369、AX-110631665、AX-109372076)能够同时在Q+K模型和FarmCPU模型中被检测到,对应区间分别为2A的Pb_nwafu-2、2D的Pb_nwafu-3、3B的Pb_nwafu-4和7A的Pb_nwafu-7区间,表明这些区间可能包含调控小麦铅耐受性的关键候选基因,这些SNP可用于进一步开发小麦铅耐受性的分子标记。
2.6 小麦铅耐受性候选基因预测
综合Q+K混合线性模型筛选出的20个SNP以及FarmCPU筛选出的8个QTL区间,利用SNP序列检索中国春小麦基因组数据库,获得其区间序列,在NCBI数据库中进行BLASTx序列比对,结合从Ensemble Plants数据库下载的最新小麦基因组注释信息(IWGSC RefSeq v1.1),筛选8个QTL区间内可能调控小麦铅耐受性的候选基因。共筛选到13个与可能小麦铅耐受性相关的候选基因(表6)。通过同源比对、家族分析(图7-A)及保守结构域分析(图7-B),结果表明,这些候选基因的功能主要集中在氧化应激反应、蛋白质修饰等途径。

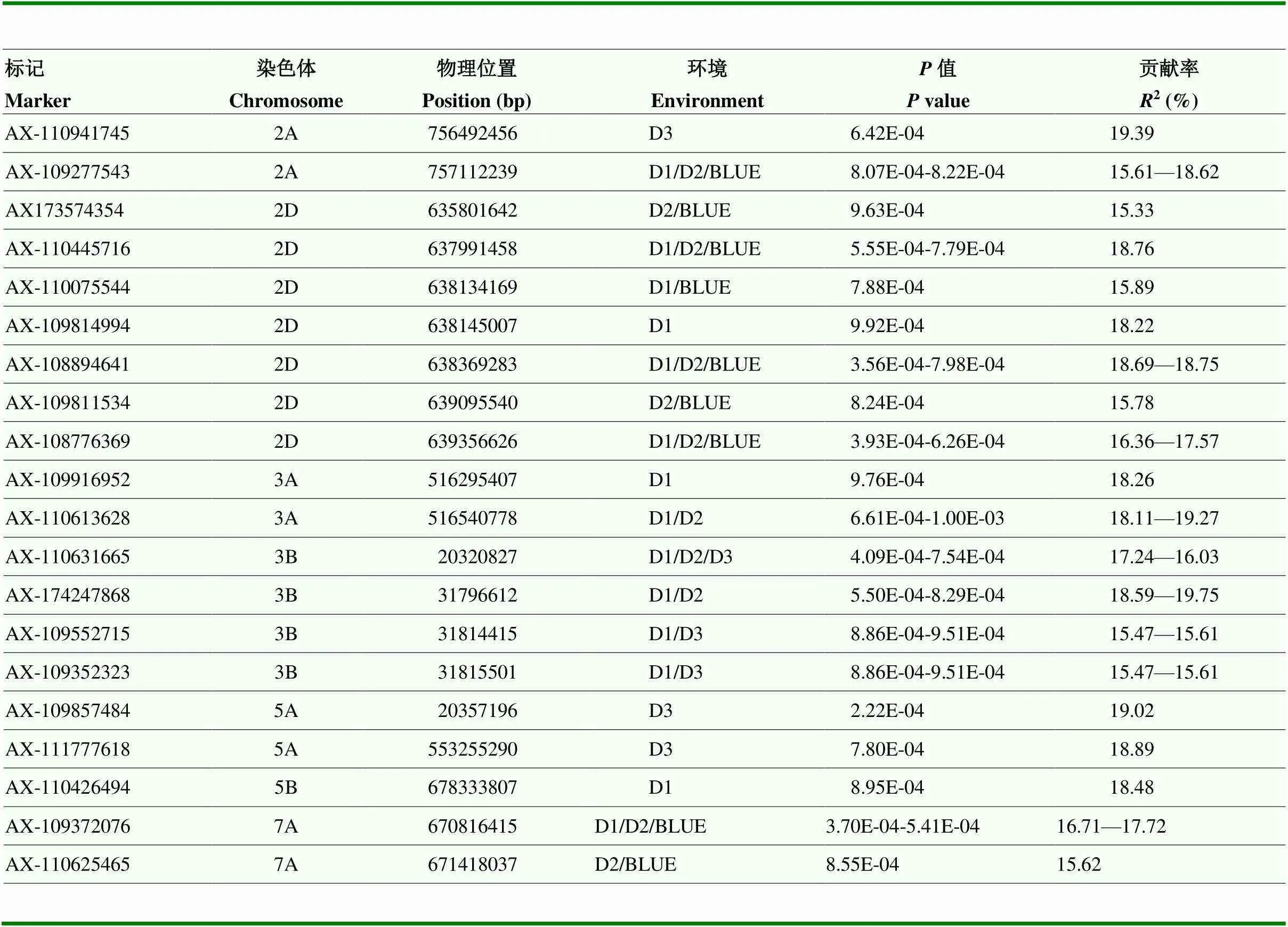
表4 GWAS检测到的小麦铅耐受性显著相关SNP位点
BLUE:最优线性无偏性值。下同 BLUE: best linear unbiased estimate.The same as below

*:在P<0.05水平差异显著;**:在P<0.01水平差异极显著

表5 FarmCPU检测到的小麦铅耐受性显著相关的区间

A:ATP结合盒转运蛋白相关候选基因与拟南芥和小麦的ABCC家族的进化树分析;B:AtMTP3与TraesCS3B02G040900的保守结构域分析
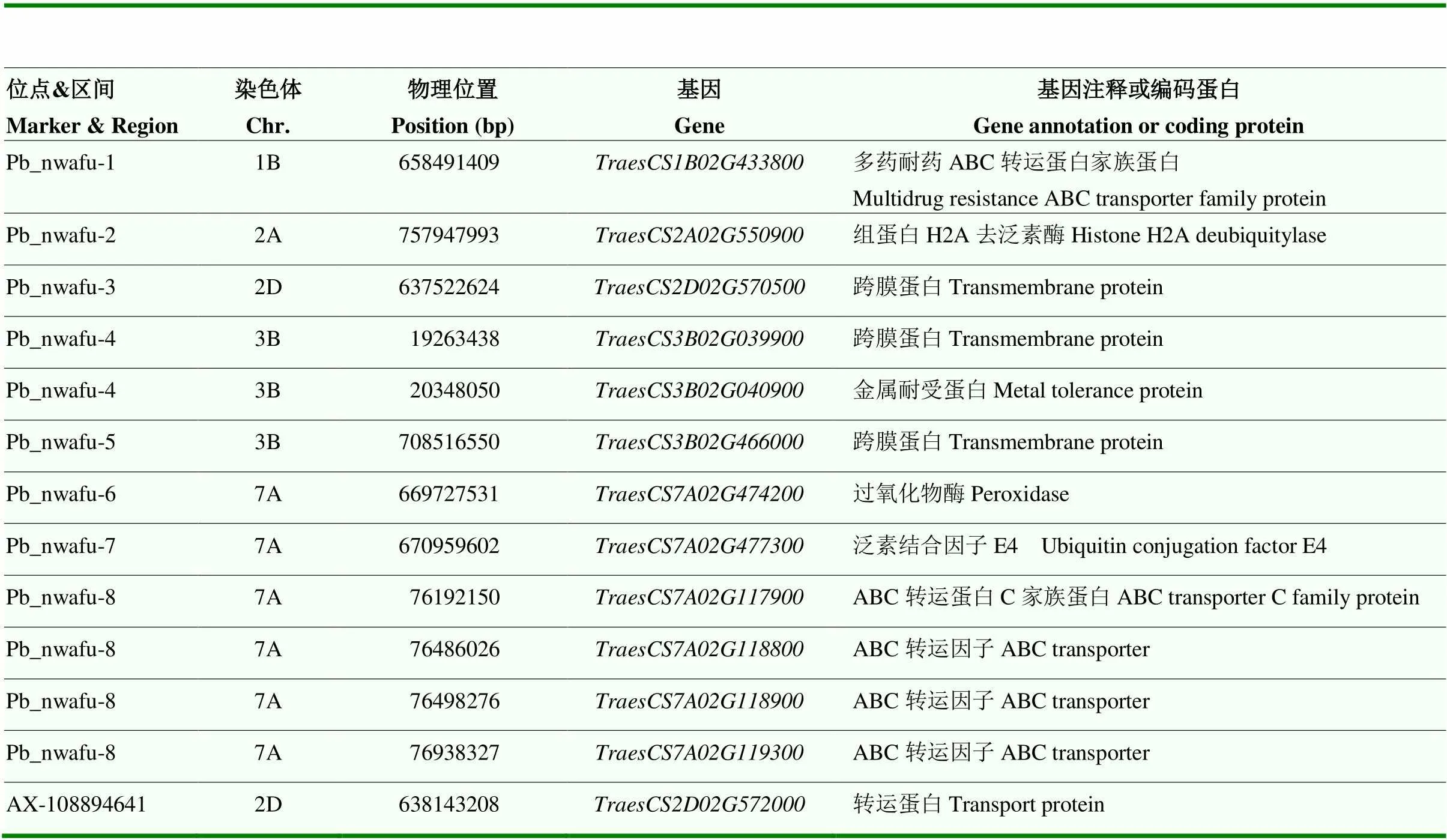
表6 筛选获得候选基因信息
3 讨论
3.1 小麦耐铅性的分析
铅是植物非必需重金属元素,会对植物生长产生一系列负面效应,包括影响光合作用、呼吸和矿物质营养[38]。本研究中,铅胁迫下多数供试材料出现最大根长显著缩短、根生物量明显减小以及生长速率变缓的现象,其中,根部受到的胁迫现象最明显,可能是由于根部直接暴露在铅溶液中。铅耐受性指植物适应和应对铅胁迫的能力,然而,在作物种质资源耐铅性鉴定和耐铅遗传改良的研究中,缺乏统一的标准,多数研究采用植株在胁迫环境下的生长状况来反应耐受性[22, 39-40]。小麦对铅胁迫的耐受性属于数量性状,易受环境影响,本研究通过正常和铅胁迫条件下测定指标的耐铅系数以及各个耐铅系数组成的加权隶属函数值值来评价小麦对铅的耐受性[24],可反映不同材料的综合耐性。本研究供试材料存在广泛的表型变异,根据耐受性值从群体中选择了7个铅耐受性强的小麦材料,可为耐铅小麦种质创新提供资源。
植物修复是目前治理重金属污染土壤的策略之一,利用重金属超富集植物从土壤中吸收重金属从而实现重金属污染土壤的修复[41]。对于中、轻度污染耕地,通过单一的重金属超富集植物进行植物修复存在不利于正常的农业生产,经济效益较低等问题[42-43],故常选择利用超富集植物对土壤进行修复的同时与低积累农作物品种进行间作的方式,目前已在玉米、大葱等农作物中得到了很好的应用[44-46]。因此,急需筛选铅低积累小麦材料,为耐铅小麦种质创新提供资源。本研究筛选的铅耐受性强、铅积累量低的小麦品种为小麦产区中轻度重金属污染农田的间作配套和健康生产提供了选择。有趣的是,在本研究筛选的7个铅耐受性品种中,最大根长耐铅系数以及生长速率耐铅系数的范围分别是0.406—0.775及0.541—1.464,而根生物量耐铅系数的变化范围为1.190—1.809,均大于1,说明铅有可能在抑制最大根长的同时增加了根直径,并最终提高了根系干物质积累量。根据对小麦耐受品种与不耐受品种地上部和根部的铅含量测定结果,发现铅胁迫下小麦根部富集的铅含量远远高于地上部富集的铅含量,这与之前的研究结果[47]一致,耐受品种与不耐受品种地上部铅含量的显著差异也表明耐受性强的小麦品种很可能是通过限制根部对铅离子的吸收、转运或通过对铅离子外排及解毒作用来降低铅积累,并最终提高小麦对铅的耐受性[10]。本研究根据耐受性系数对20个极端小麦品种的铅含量进行分析,发现铅耐受性和根系、地上部的铅积累量存在负相关关系,然而,植物对重金属离子铅的吸收、转运机制比较复杂,后续还需在更大的群体中进行验证,明确铅耐受性与铅积累量的相互关系及协调机制。
3.2 小麦铅耐受性GWAS分析
随着高通量测序和基因芯片技术的快速发展,全基因组关联分析成为一种快速简便分析复杂性状遗传变异的方法,已经成功应用于多种作物基本性状的遗传变异研究,包括小麦品质、水稻产量、大豆光合作用等性状[25, 48]。STICH等[49]研究认为,Q+K混合线性模型是全基因组关联分析中降低假阳性的最好方法之一。本研究采用Q+K混合线性模型对102份供试材料的小麦铅耐受性进行全基因组关联分析,同时,为了提高关联分析的准确率以及避免忽略微效位点,本研究同时还采用FarmCPU方法[50],以验证补充Q+K混合线性模型。目前,利用全基因组关联分析方法研究小麦铅耐受性、挖掘耐性相关基因的报道比较少,本研究通过全基因组关联分析方法共检测到20个与铅耐受性显著关联SNP位点和8个关键QTL区间。其中,候选区间Pb_nwafu-2内有拟南芥的同源基因(小麦2A染色体753 Mb处),该基因对铅离子进入植物细胞的运输途径具有调控作用。候选区间Pb_nwafu-7附近有拟南芥基因的同源基因(小麦7A染色体671 Mb处),该基因编码ATP结合盒转运蛋白,参与拟南芥对铅的解毒作用[51]。因此,候选区间Pb_nwafu-2和Pb_nwafu-7可以作为小麦耐铅分子机制研究的重点区间,可在此区间挖掘相关候选基因,为小麦耐铅胁迫分子机制研究提供参考。同时,为进一步验证部分SNP单倍型与铅耐受性的关系,本研究选择了9个SNP进行检验,其中,有5个SNP的2种纯合基因型间的铅耐受性存在显著差异(<0.05),且这5个SNP能够在多次重复及2种关联分析方法中检测到,可作为小麦铅耐受性改良的优势位点和选择标记。后续研究中,可依据这5个SNP标记的位置及候选基因筛选结果,选择2A、2D、3B、7A染色体上对应的区间作为热点区间,进一步在更大的遗传群体中验证标记的实用性,并深入分析相关候选基因的作用机制。
3.3 铅耐受性相关候选基因预测
本研究共筛选到13个可能与小麦铅耐受性相关的基因,位于2D、3B上的3个基因、、编码跨膜蛋白。位于2D上的基因编码转运蛋白。位于1B、7A上的6个基因编码ATP结合盒转运蛋白,ATP结合盒转运蛋白(ABC)主要通过结合和水解ATP释放能量,导致蛋白质构象变化,从而实现包括重金属螯合物、金属离子在内的多种底物的跨膜运输[52-53]。ABCC多药耐药蛋白家族(ABCC- MRP)是ABC转运蛋白家族的一个亚家族,参与包括细胞稳态、金属解毒、谷胱甘肽结合物转运在内的多种生理过程[54]。本研究候选基因、和与拟南芥和同源性较高,且有研究表明和能够介导拟南芥对镉和汞的耐受性[55],因此,这3个候选基因可以作为今后铅胁迫遗传机制的研究重点。位于3B染色体上的编码金属耐受蛋白,金属耐受蛋白(metal tolerance protein,MTP)可以催化过渡金属离子从胞质外进入亚细胞区,影响植物对重金属的耐性和积累[56],与拟南芥(与金属二价阳离子耐受性相关的基因,图7-B)具有相同的保守结构域,该保守结构与阳离子外排相关[57],且根据小麦公共表达量数据库显示在根中表达。位于7A的基因编码过氧化物酶,重金属胁迫可能会导致小麦体内产生过量的活性氧(reactive oxygen species,ROS),进一步诱导氧化应激反应,进而影响植物对重金属的耐性和积累[10]。位于2A、7A的2个基因、编码分别编码组蛋白H2A去泛素化酶和泛素结合因子,泛素和去泛素化对蛋白质的修饰作用可以减轻重金属对植物的毒害作用[22]。本研究筛选的候选基因编码不同蛋白,在植物抵御重金属胁迫中起直接或间接作用,其中、、和可以作为后续小麦耐铅分子机制研究的重点。
4 结论
通过Q+K混合线性模型获得20个与小麦铅耐受性显著关联的遗传位点,通过FarmCPU模型检测到8个与小麦铅耐受性显著关联的QTL,2种模型的分析结果在物理位置上存在较好的一致性。最终获得13个与小麦铅耐受性直接或间接相关的候选基因,分别编码不同转运蛋白、过氧化物酶、跨膜蛋白以及泛素
与去泛素化,可能通过参与氧化应激反应、蛋白质修饰等途径来调节小麦对铅的耐受性。
[1] ZHAO F J, Ma Y B, ZHU, Y G, TANG Z, McGrath S P.Soil contamination in China: current status and mitigation strategies.Environmental Science & Technology, 2015, 49(2): 750-759.
[2] WILLIAMS P N, LEI M, SUN G X, HUANG Q, LU Y, DEACON C, MEHARG A A, ZHU Y G.Occurrence and partitioning of cadmium, arsenic and lead in mine impacted paddy rice: hunan, China.Environmental Science & Technology, 2009, 43(3): 637-642.
[3] ZHUANG P, MCBRIDE M B, XIA H P, LI N Y, LIA Z A.Health risk from heavy metals via consumption of food crops in the vicinity of dabaoshan mine, south China.Science of the Total Environment, 2009, 407(5): 1551-1561.
[4] 徐长春, 熊炜, 郑戈, 林友华.“农业面源和重金属污染农田综合防治与修复技术研发”专项组织实施进展分析.农业环境科学学报, 2017, 36(7): 1242-1246.
XU C C, XIONG W, ZHENG G, LIN Y H.Progress of the program for research and development on comprehensive prevention and remediation techniques for agricultural non-point source and heavy metal polluted croplands.Journal of Agro-Environment Science, 2017, 36(7): 1242-1246.(in Chinese)
[5] LUO S L, CALDERON U A, YU J H, LIAO W B, XIE J M, Lü J, FENG Z, TANG Z Q.The role of hydrogen sulfide in plant alleviates heavy metal stress.Plant and Soil, 2020, 449(1/2): 1-10.
[6] RUFF H A, MARKOWITZ M E, BIJUR P E, ROSEN J F.Relationships among blood lead levels, iron deficiency, and cognitive development in two-year-old children.Environmental Health Perspectives, 1996, 104(2): 180-185.
[7] 张玉林.资本的秩序与乡村居民铅中毒——关于河南省多个案例的分析.江苏行政学院学报, 2017(3): 67-76.
ZHANG Y L.Capital order and lead poisoning of rural residents— analysis of multiple cases in henan province.Journal of Jiangsu Administration Institute, 2017(3): 67-76.(in Chinese)
[8] 薛涛, 廖晓勇, 王凌青, 张扬珠.镉污染农田不同水稻品种镉积累差异研究.农业环境科学学报, 2019, 38(8): 1818-1826.
XUE T, LIAO X Y, WANG L Q, ZHANG Y Z.Cadmium accumulation in different rice cultivars from cadmium-polluted paddy fields.Journal of Agro-Environment Science, 2019, 38(8): 1818-1826.(in Chinese)
[9] 宋淑艳, 拜丽克孜·买提库尔班.重金属镉、铅对小麦的影响及改良剂缓解效果研究进展专题综述.天津农林科技, 2021(1): 41-43.
SONG S Y, BERIKZI M.A review of the research progress on the effects of heavy metals cadmium and lead on wheat and the mitigation effects of improvers.Tianjin Agriculture and Forestry Science and Technology, 2021(1): 41-43.(in Chinese)
[10] YAN A, WANG Y M, TAN S N, YUSOF M L M, GHOSH S, CHEN Z.Phytoremediation: a promising approach for revegetation of heavy metal-polluted land.Frontiers in Plant Science, 2020, 11: 359.
[11] JIANG L, WANG W, CHEN Z, GAO Q, XU Q, CAO H.A role for APX1 gene in lead tolerance in.Plant Science, 2017, 256: 94-102.
[12] KIM D Y, BOVET L, NOH E W, MARTINOIA E, LEE Y.AtATM3 is involved in heavy metal resistance in.Plant Physiology, 2006, 140(3): 922-932.
[13] KIM D Y, BOVET L, MAESHIMA M, MARTINOIA E, LEE Y.The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance.The Plant Journal, 2007, 50(2): 207-218.
[14] XIAO S, GAO W, CHEN Q F, RAMALINGAM S, CHYE M L.Overexpression of membrane-associated acyl-CoA-binding protein ACBP1 enhances lead tolerance in.The Plant Journal, 2008, 54(1): 141-151.
[15] ARAZI T, SUNKAR R, KAPLAN B, FROMM H.A tobacco plasma membrane calmodulin-binding transporter confers Ni2+tolerance and Pb2+hypersensitivity in transgenic plants.The Plant Journal, 1999, 20(2): 171-182.
[16] SCHUURINK R C, SHARTZER S F, FATH A, JONES R L.Characterization of a calmodulin-binding transporter from the plasma membrane of barley aleurone.Proceedings of the National Academy of Science of the United States of America, 1998, 95(4): 1944-1949.
[17] HINDU V, PALACIOS-ROJAS N, BABU R, SUWARNO W B, RASHID Z, USHA R, SAYKHEDKAR G R, NAIR S K.Identification and validation of genomic regions influencing kernel zinc and iron in maize.Theoretical and Applied Genetics, 2018, 131(7): 1443-1457.
[18] PANG Y L, WU Y Y, LIU C X, LI W H, ST AMAND P, BERNARDO A, WANG D F, DONG L, YUAN X F, ZHANG H R, ZHAO M, LI L Z, WANG L M, HE F, LIANG, Y L, YAN Q, LU Y, SU Y, JIANG H M, WU J J, LI A F, KONG L R, BAI G H, LIU S B.High-resolution genome-wide association study and genomic prediction for disease resistance and cold tolerance in wheat.Theoretical and Applied Genetics, 2021, 134(9): 2857-2873.
[19] ZHONG H, LIU S, SUN T, KONG W L, DENG X X, PENG Z H, LI Y S.Multi-locus genome-wide association studies for five yield- related traits in rice.BMC Plant Biology, 2021(21): 364.
[20] PAN X W, LI Y C, LIU W Q, LIU S X, MIN J, XIONG, H B, DONG Z, DUAN Y H, YU Y Y, LI X X.QTL mapping and candidate gene analysis of cadmium accumulation in polished rice by genome-wide association study.Scientific Reports, 2020, 10: 11791.
[21] QIN P, WANG L, LIU K, MAO S S, LI Z Y, GAO S, SHI H R, LIU Y X.Genome-wide association study oftraits under seedling-stage cadmium stress.Crop Journal, 2015, 3(5): 405-415.
[22] ZHANG F G, XIAO X, XU K, CHENG X, XIE T, HU J H, WU X M.Genome-wide association study (GWAS) reveals genetic loci of lead (Pb) tolerance during seedling establishment in rapeseed (L.).BMC Genomics, 2020, 21(1): 139.
[23] 张寒, 潘香逾, 王秀华, 李家丽, 姜慧新, 赵岩.苜蓿萌发期耐盐性综合评价与耐盐种质筛选.草地学报, 2018, 26(3): 666-672.
ZHANG H, PAN X Y, WANG X H, LI J L, JIANG H X, ZHAO Y.Comprehensive evaluation of salt tolerance and screening for salt tolerance germplasm of alfalfa (medicago) at germination stage.Acta Agrestia Sinica, 2018, 26(3): 666-672.(in Chinese)
[24] 陈新, 吴斌, 张宗文.燕麦种质资源重要农艺性状适应性和稳定性评价.植物遗传资源学报, 2016, 17(4): 577-585.
CHEN X, WU B, ZHANG Z W.Evaluation of adaptability and stability for important agronomic traits of Oat (spp.) germplasm resources.Journal of Plant Genetic Resources, 2016, 17(4): 577-585.(in Chinese)
[25] YANG Y, CHAI Y M, ZHANG X, LU S, ZHAO Z C, WEI D, CHEN L, HU Y G.Multi-locus GWAS of quality traits in bread wheat: mining more candidate genes and possible regulatory network.Frontiers in Plant Science,2020, 11: 1091.
[26] ZHANG F G, XIAO X, XU K, CHENG X, XIE T, HU J H.Genome-wide association study (GWAS) reveals genetic loci of lead (Pb) tolerance during seedling establishment in rapeseed (L.).BMC Genomics,2020, 21(1): 139.
[27] 戴海芳, 武辉, 阿曼古丽·买买提阿力, 王立红, 麦麦提·阿皮孜, 张巨松.不同基因型棉花苗期耐盐性分析及其鉴定指标筛选.中国农业科学, 2014, 47(7): 1290-1300.
DAI H F, WU H, AMANGULI M, WANG L H, MAIMAITI A, ZHANG J S.Analysis of salt-tolerance and determination of salt-tolerance evaluation indicators in cotton seedlings of different genotypes.Scientia Agricultura Sinica,2014, 47(7): 1290-1300.(in Chinese)
[28] THOMAS C L, ALCOCK T D, GRAHAM N S, HAYDEN R, MATTERSON S, WILSON L, YOUNG S D, DUPUY L X, WHITE P J, HAMMOND J P, DANKU JMC, SALT D E, SWEENEY A, BANCROFT I, BROADLEY M R.Root morphology and seed and leaf ionomic traits in aL.diversity panel show wide phenotypic variation and are characteristic of crop habit.BMC Plant Biology,2016(16): 214.
[29] UTMAZIAN M N D, WIESHAMMER G, VEGA R, WENZEL W W.Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars.Environmental Pollution,2007, 148(1): 155-165.
[30] YUN L, LARSON S R, JENSEN K B, STAUB J E, GROSSL P R.Quantitative trait loci (QTL) and candidate genes associated with trace element concentrations in perennial grasses grown on phytotoxic soil contaminated with heavy metals.Plant and Soil,2015, 396(1/2): 277-296.
[31] 康吉利, 曾志军, 刘玉佩.铅胁迫对小麦种子萌发及幼苗生长的影响.广西农业科学, 2009, 40(2): 144-146.
KANG J L, ZENG Z J, LIU Y P.Effects of lead(Pb2+)stress on seed germination and seedling growth of wheat.Guangxi Agricultural Sciences, 2009, 40(2): 144-146.(in Chinese)
[32] 耿雷跃, 马小定, 崔迪, 张启星, 韩冰, 韩龙植.水稻全生育期耐盐性鉴定评价方法研究.植物遗传资源学报, 2019, 20(2): 267-275.
GENG L Y, MA X D, CUI D, ZHANG Q X, HAN B, HAN L Z.Identification and evaluation method for saline tolerance in rice during the whole growth stage.Journal of Plant Genetic Resources, 2019, 20(2): 267-275.(in Chinese)
[33] ZHANG C, DONG S S, XU J Y, HE W M, YANG T L.PopLDdecay: a fast and effective tool for linkage disequilibrium decay analysis based on variant call format files.Bioinformatics, 2019, 35(10): 1786-1788.
[34] YU S Z, WU J H, WANG M, SHI W M, XIA G M, JIA J Z,KANG Z S, HAN D J.Haplotype variations in QTL for salt tolerance in Chinese wheat accessions identified by marker-based and pedigree- based kinship analyses.Crop Journal, 2020, 8(6): 1011-1024.
[35] 王继庆, 任毅, 时晓磊, 王丽丽, 张新忠, 苏力坛·姑扎丽阿依, 谢磊, 耿洪伟.小麦籽粒超氧化物歧化酶(SOD)活性全基因组关联分析.中国农业科学, 2021, 54(11): 2249-2265.
WANG J Q, REN Y, SHI X L, WANG L L, ZHANG X Z, SULITAN G, XIE L, GENG H W.Genome-wide association analysis of superoxide dismutase (SOD) activity in wheat grain.Scientia Agricultura Sinica,2021, 54(11): 2249-2265.(in Chinese)
[36] 周思远, 毕惠惠, 程西永, 张旭睿, 闰永行, 王航辉, 毛培钧, 李海霞, 许海霞.小麦耐低磷相关性状的全基因组关联分析.植物遗传资源学报, 2020, 21(2): 431-445.
ZHOU S Y, BI H H, CHENG X Y, ZHANG X R, RUN Y X, WANG H H, MAO P J, LI H X, XU H X.Genome-wide association study of low-phosphorus tolerance related traits in wheat.Journal of Plant Genetic Resources, 2020, 21(2): 431-445.(in Chinese)
[37] APPELS R, EVERSOLE K, FEUILLET C, KELLER B, ROGERS J, STEIN N, POZNIAK C J, CHOULET F, DISTELFELD A, POLAND J.Shifting the limits in wheat research and breeding using a fully annotated reference genome.Science, 2018(361): 661.
[38] AHMAD M S A, ASHRAF M, TABASSAM Q, HUSSAIN M, FIRDOUS H.Lead (pb)-induced regulation of growth, photosynthesis, and mineral nutrition in maize (L.) plants at early growth stages.Biological Trace Element Research, 2011, 144(1/3): 1229-1239.
[39] 赵鲁, 叶琰, 刘继远, 刘安辉, 高振新, 王小龙, 李洋, 王震, 佟明恒.添加铅对大豆和小麦生长及铅吸收特征影响的研究.中国土壤与肥料, 2013(6): 83-87.
ZHAO L, YE Y, LIU J Y, LIU A H, GAO Z X, WANG X L, LI Y, WANG Z, TONG M H.Study on the effect of lead addition on the growth and lead absorption characteristics of soybean and wheat.China Soil and Fertilizer, 2013(6): 83-87.(in Chinese)
[40] 厉有为, 梁婵娟.三种油料作物对土壤Pb污染的耐受性与积累.环境化学, 2021, 40(5): 1602-1610.
LI Y W, LIANG C J.Tolerance and accumulation of lead in three oil crops to lead pollution in soil.Environmental Chemistry, 2021, 40(5): 1602-1610.(in Chinese)
[41] Ayangbenro A S, Babalola O O.A new strategy for heavy metal polluted environments: a review of microbial biosorbents.International Journal of Environmental Research and Public Health, 2017, 14(1): 94.
[42] 王玲芬, 谢明.对多起铅污染中毒事件处理的体会与思考.中国卫生监督杂志, 2011, 18(5): 485-487.
WANG L F, XIE M.Experience and thinking on handling multiple lead pollution incidents.Chinese Journal of Health Inspection, 2011, 18(5): 485-487.(in Chinese)
[43] 郭思宇, 王海娟, 王宏镔.重金属污染土壤间作修复的研究进展.中国生态农业学报(中英文), 2021, 29(5): 890-902.
GUO S Y, WANG H J, WANG H B.Advances in the intercropping remediation of heavy metal polluted soil.Chinese Journal of Eco-Agriculture, 2021, 29(5): 890-902.(in Chinese)
[44] 秦丽, 湛方栋, 祖艳群, 孟婧轩, 晋磊, 李元.土荆芥和蚕豆/玉米间作系统中Pb、Cd、Zn的累积特征研究.云南农业大学学报(自然科学), 2017, 32(1): 153-160.
QIN L, ZHAN F D, ZU Y Q, MENG J X, JIN L, LI Y.Accumulation characteristics of Pb,Cd and Zn byL.Intercropping with maize and broad bean.Journal of Yunnan Agricultural University (Natural Science), 2017, 32(1): 153-160.(in Chinese)
[45] 陈国皓, 祖艳群, 湛方栋, 李博, 李元.钝化剂处理对玉米与伴矿景天间作下植株生长及镉累积特征的影响.农业环境科学学报, 2019, 38(9): 2103-2110.
CHEN G H, ZU Y Q, ZHAN F D, Li B, Li Y.Effects of passivators on the growth and cadmium accumulation of intercropped maize and Sedum plumbizincicola.Journal of Agro-Environment Science, 2019, 38(9): 2103-2110.(in Chinese)
[46] WANG S Q, WEI S H, JI D D, BAI J Y.Co-Planting Cd contaminated field using hyperaccumulatorL.through interplant with low accumulation welsh onion.International Journal of Phytoremediation, 2015, 17(9): 879-884.
[47] NIU Z X, SUN L N, SUN T H.Enrichment characteristics of Cd and Pb by four kinds of plant under hydroponic culture.Chinese Journal of Ecology, 2010, 29(2): 261-268.
[48] HUANG X H, ZHAO Y, WEI XH, LI C Y, WANG A, ZHAO Q, LI W J, GUO Y L, DENG L W, ZHU C R, FAN D L, LU Y Q, WENG Q J, LIU K Y, ZHOU T Y, JING Y F, SI L Z, DONG G J, HUANG T, LU T T, FENG Q, QIAN Q, LI J Y, HAN B.Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm.Nature Genetics, 2012, 44(1): 32-39.
[49] STICH B, MELCHINGER A E.Comparison of mixed-model approaches for association mapping in rapeseed, potato, sugar beet, maize, and.BMC Genomics, 2009, 10: 94.
[50] LIU X, HUANG M, FAN B.Iterative usage of fixed and random effect models for powerful and efficient Genome-Wide association studies.PlosOne, 2016, 12: e10059573.
[51] LEE M, LEE K, LEE J, NOH E W, LEE Y.AtPDR12 contributes to lead resistance in.Plant Physiology, 2005, 138(2): 827-836.
[52] GAILLARD S, JACQUET H, VAVASSEUR A, LEONHARDT N, FORESTIER C.AtMRP6/AtABCC6, an ATP-binding cassette transporter gene expressed during early steps of seedling development and up-regulated by cadmium in.BMC Plant Biology, 2008, 8: 22.
[53] THEODOULOU F L, KERR I D.ABC transporter research: going strong 40 years on.Biochemical Society Transactions, 2015, 43(5): 1033-1040.
[54] BHATI K K, SHARMA S, AGGARWAL S, KAUR M, SHUKLA V, KAUR J, MANTRI S, PANDEY A K.Genome-wide identification and expression characterization of ABCC-MRP transporters in hexaploid wheat.Frontiers in Plant Science, 2015, 6: 488.
[55] PARK J, SONG, W Y, KO D, EOM Y, HANSEN T H, SCHILLER M, LEE T G, MARTINOIA E, LEE Y.The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury.The Plant Journal, 2012, 69(2): 278-288.
[56] OVECKA M, TAKAC T.Managing heavy metal toxicity stress in plants: Biological and biotechnological tools.Biotechnology Advances, 2014, 32(1SI): 73-86.
[57] ARRIVAULT S, SENGER T, KRAEMER U.Themetal tolerance protein AtMTP3 maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency and Zn oversupply.The Plant Journal, 2006, 46(5): 861-879.
Genome-Wide Association Analysis of Lead Tolerance in Wheat at Seedling Stage
ZHI Lei1, ZHE Li1, SUN NanNan1, YANG Yang1, Dauren Serikbay1,2, JIA HanZhong3, HU YinGang1, CHEN Liang1*
1College of Agronomy, Northwest A&F University/State Key Laboratory of Crop Stress Biology for Arid Areas, Yangling 712100, Shaanxi, China;2S.Seifullin Kazakh Agro Technical University, Nursultan 010011, Kazakhstan;3College of Resources and Environment, Northwest A&F University, Yangling 712100, Shaanxi, China
【Objective】With the advancement of industrialization, the pollution of arable land by heavy metals, especially lead, has become a worldwide problem.Wheat is an important food crop, and its safe cultivation is critical to maintaining food security.Screening wheat varieties with strong tolerance to lead, low lead accumulation and mining relevant regulatory genes or QTL regions would lay foundation for further elucidating the genetic mechanism of lead tolerance in wheat.【Method】The tolerance to lead of 102 wheat varieties (advanced lines) were evaluated with a 140 mg·kg-1lead nitrate solution at the seedling stage, by the weighted membership function value (D Value) of the lead tolerance coefficients of maximum root length, root biomass and growth rate under three replicates, combining with the 335 438 high-quality SNPs(single nucleotide polymorphism)markers by wheat 660K SNP chip, a genome-wide association study (GWAS) for lead tolerance in wheat was conducted, to mine the candidate genes for lead tolerance in wheat.【Result】The lead among between wheat varieties (advanced lines) under different replicates showed rich variation, with the coefficient of variation of 44.8%-46.2%, and the correlation coefficient was between 0.87-0.97 (<0.001).It was found that varieties with strong lead tolerance showed low lead accumulation characteristics.The genotyping results showed that the SNP polymorphic information content (PIC) was 0.28-0.32, the population structure analysis showed that these wheat materials could be divided into 7 subgroups; a total of 20 SNPs and 8 key chromosomal segments that were significantly associated with lead tolerance in wheat (≤0.001) were detected by two GWAS methods respectively distributed on chromosomes 1B, 2A, 2D, 3A, 3B, 5A, and 7A.A single locus explains 15.33%-19.75% of the phenotypic variation, and 10 and 8 key chromosomal segments were detected in two and more environments.Analysis of candidate genes for significant and stable association sites and intervals revealed that the functions of the candidate genes were mainly related to transmembrane transport, protein modification and oxidative stress response, including 7 genes related to transporters, including,,etc.Two genes involved in ubiquitination and deubiquitination(and), three genes encoding transmembrane proteins (,and), one gene related to peroxidase ().【Conclusion】Seven wheat materials with strong lead tolerance were screened, 20 SNPs in 8 candidate regions significantly related to lead tolerance in wheat were detected, and 13 candidate genes related to lead tolerance in wheat were finally screened.
wheat; lead tolerance; genome-wide association analysis; candidate gene analysis
2021-09-18;
2021-11-16
国家自然科学基金(31671695)、陕西省重点研发计划(2020NY-045)
职蕾,E-mail:zl9711110@163.com。通信作者陈亮,E-mail:chenliang9117@qq.com
(责任编辑 李莉)
猜你喜欢
分子催化(2022年1期)2022-11-02
分析测试学报(2022年9期)2022-09-21
中国农业科学(2022年16期)2022-09-19
军事文摘(2022年16期)2022-08-24
今日农业(2022年4期)2022-06-01
中国典型病例大全(2022年7期)2022-04-22
电脑知识与技术(2018年19期)2018-11-01
现代农业科技(2016年20期)2016-12-20
现代养生·下半月(2015年6期)2015-09-07
现代养生·下半月(2015年2期)2015-06-15
