
枣R2R3-MYB亚家族基因鉴定及其在果实发育中的表达分析
2022-05-16 08:44李世佳吕紫敬赵锦
中国农业科学 2022年6期
李世佳,吕紫敬,赵锦
枣R2R3-MYB亚家族基因鉴定及其在果实发育中的表达分析
李世佳,吕紫敬,赵锦*
河北农业大学生命科学学院,河北保定 071000
【目的】MYB作为植物中最大的基因家族之一,在植物生长、果实发育等方面具有重要作用。本研究拟鉴定枣R2R3-MYB亚家族成员,并进行生物信息学和表达模式分析,为深入探究该类基因在枣生长发育尤其是果实发育中的功能提供参考。【方法】以拟南芥R2R3-MYB亚家族成员为探针,利用BLAST和HMMER方法,在枣全基因组数据库中筛选R2R3-MYB亚家族成员;利用植物转录因子、UniProt、Gcorn plant等数据库,采用MEGA-x、ExPASy、TBtools、PheatMap等软件进行生物信息学分析;利用转录组和实时荧光定量PCR技术分析该类基因的表达,对重点基因进行互作蛋白的预测及验证。【结果】在枣全基因组范围内,鉴定出118个R2R3-MYB亚家族成员,均具有典型的MYB超家族结构域,分为18个亚组,依次命名为—。理化性质分析表明,该家族基因编码的氨基酸数目介于200—400;95个成员定位到12条染色体上,其中4号染色体上分布最多;谱系分析表明该类基因进化过程中发生了直系和旁系同源事件。通过转录组和实时荧光定量PCR分析,发现一些R2R3-MYB基因参与枣果实的发育过程,并进一步筛选出了可能与果实膨大和色泽发育相关的候选基因、。并利用蛋白互作预测和酵母双杂试验证明ZjMYB104和ZjMPK3存在互作。【结论】鉴定出118个枣R2R3-MYB亚家族成员,结构高度保守,分布在12条染色体上,在进化中发生了直系和旁系同源事件,并筛选出了与果实发育密切相关的候选基因,为进一步的功能分析提供了线索和参考。
枣;R2R3-MYB亚家族基因;生物信息学分析;表达分析;果实发育;蛋白互作
0 引言
【研究意义】v-myb禽成髓细胞瘤病毒致癌基因同源物(v-myb avian myeloblastosis viral oncogene homolog,MYB)转录因子家族作为植物中最大的基因家族之一[1-2],在植物生长发育、次生代谢及抗逆胁迫等方面中具有重要作用[3]。R2R3-MYB作为MYB家族中成员最多的一类[4-6],在植物生长发育过程中扮演着重要角色[7],鉴定并研究其参与的调控代谢有重要意义。枣(Mill.)属于鼠李科枣属植物,原产于中国,产量居干果第一位[8],鉴定枣R2R3-MYB基因可为研究该类基因在枣生长发育中的作用提供基础数据,分析该类基因在果实发育中的表达模式将为深入探究其调控机制提供线索。【前人研究进展】目前,在拟南芥[9]、茄科植物[10]、梨[11]、苹果[12]、水稻[13]和杨树[14]等基因组中均鉴定出了MYB家族成员,均以R2R3-MYB亚家族成员数目最多。研究发现R2R3-MYB亚家族不仅参与植物激素信号传导、抗逆胁迫等生理生化活动,而且与果实膨大、着色等果实生长发育过程密切相关[15-16]。葡萄和调控下游、和的转录活性,影响葡萄果实的生长发育[17]。梨的启动子甲基化抑制的表达和花青苷的生物合成,从而影响梨果皮着色过程[18]。草莓中MYB基因随果实的生长发育,表达量逐渐增加[19];同时,的变异等位基因,作为一个提前的终止密码子,编码蛋白的C-末端,导致草莓果实白化[20]。苹果中的启动子中微卫星的插入导致苹果果肉发育为红色[21]。2019年,Ji等[22]已鉴定出171个枣MYB基因家族成员,其中,99个枣R2R3-MYB亚家族成员。【本研究切入点】虽然枣MYB基因家族已有报道,但还未见针对R2R3-MYB亚家族的研究,且该亚家族成员与果实发育的关系尚不明确。【拟解决的关键问题】鉴定枣R2R3-MYB亚家族成员,对其理化性质、染色体定位、系统进化树及保守基序进行分析,通过转录组和实时荧光定量PCR分析该类基因在枣果发育中的表达,并利用酵母双杂技术验证互作蛋白,为开展该类基因的功能分析提供依据。
1 材料与方法
试验于2020年在河北农业大学进行。
1.1 枣R2R3-MYB亚家族基因的生物信息学分析
1.1.1 枣R2R3-MYB亚家族成员的鉴定 利用冬枣全基因组数据信息(JREP00000000.1),并以拟南芥R2R3-MYB亚家族成员为探针,基于局部比对算法的搜索工具(basic local alignment search tool,Blast)及HMMER(https://www.ebi.ac.uk/Tools/hmmer/search/ hmmsearch)网站中马尔科夫概率模型的方法,分别利用蛋白序列层次的相似性和蛋白结构域层次的同源性,在枣全基因组信息中搜索。将两个来源的序列结果用植物转录因子数据库(Plant Transcription Factor Database,http://planttfdb.gao-lab.org/index.php)和UniProt库(UniProt Knowledgebase,UniProtKB,https:// www.uniprot.org/)信息进行矫正,并删除重复及预测错误序列,最终得到该亚家族成员蛋白序列,根据其染色体位置信息重新命名。同时,使用ExPASy在线分析软件(https://web.expasy.org/compute_pi/)预测鉴定得到的蛋白等电点和分子量。
1.1.2 枣R2R3-MYB亚家族基因染色体定位及进化树构建 根据枣R2R3-MYB亚家族的染色体定位信息,利用TBtools工具[23]中的Gene Location Visualize功能绘制染色体定位图。利用MEGA-X软件,采用最大似然法(maximum likelihood,ML)构建进化树,并设置校验参数Bootstrap重复1 000次,位点覆盖范围设置为80%。
1.1.3 枣R2R3-MYB蛋白的同源域比对、保守基序及基因结构分析 使用NCBI中CD-Search功能对枣R2R3-MYB蛋白的保守结构域进行比对分析。用在线软件MEME(http://meme-suite.org/)鉴定蛋白序列的保守基序,最大基序数量设置为10,其他参数保留默认值。枣R2R3-MYB亚家族成员的基因结构信息来自其基因结构注释文件。
1.1.4 枣R2R3-MYB重点基因的谱系及进化分析 使用Gcorn植物(Gcorn plant)数据库(http://www.plant.osakafu-u.ac.jp/~kagiana/gcorn/p/),对枣R2R3-MYB重点基因进行谱系分析,得到相应基因的同源性信息以及进化信息。
1.2 枣R2R3-MYB基因在果实发育过程中的表达模式分析
选取两个主栽品种‘金丝小枣’和‘金魁王’为试材,采样地点为河北沧县枣树资源圃。采样时期将枣果从幼果到全红分为9个时期:S1—S9,即幼果期1、幼果期2、硬核前期、硬核期、白熟期、白熟后期、1/4着色期、半红期、全红期,每个时期设3次生物学重复,采用TaKaRa Mini BEST Plant RNA Extraction Kit试剂盒提取各时期样本的RNA,微型分光光度计和琼脂糖凝胶电泳对RNA进行质量检测,质量合格的RNA备用。在Illumina HiSeq 2000高通量测序平台进行转录组测序,使用PheatMap绘制表达热图。
选择‘金丝小枣’5个果实发育时期,幼果期、白熟前、白熟期、半红期以及全红期进行代表基因和的表达验证取0.5 µg RNA,采用TIANGEN Fastk gDNA Dispelling RT Super Mix试剂盒合成cDNA。使用实时荧光定量PCR仪和SuperReal PreMix Plus(SYBR Green)试剂盒以cDNA为模板进行表达分析。内参基因及引物序列见表1。20 µL反应体系:1 µL稀释cDNA模板,上、下游引物各0.4 µL,Power SYBR ® Green PCR Master Mix(2×)10 µL,ddH2O 8.2 µL。PCR反应程序:95℃预变性15min;40个循环(95℃ 10s, 60℃ 20 s)。每样品设4个生物学重复,分析荧光值变化曲线及熔解曲线,采用2-∆∆CT计算表达水平。
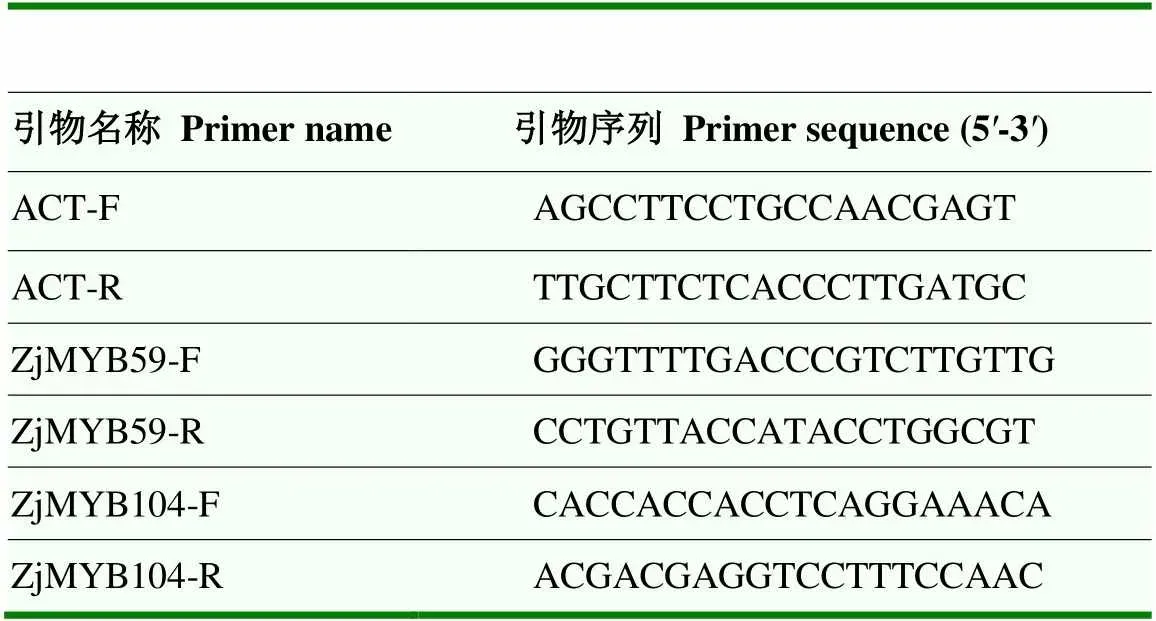
表1 实时荧光定量PCR引物名称及序列
1.3 互作蛋白的预测及验证
利用STRING(https://string-db.org/)网站,以拟南芥相应的同源蛋白为参考,对候选蛋白ZjMYB104进行互作蛋白预测分析。同时,设计酵母双杂交引物(表2),扩增和全长,连接pMD18-T克隆载体,转化大肠杆菌感受态细胞DH5α,筛选得到阳性克隆。经测序比对后,提取重组质粒,进行酶切、连接,构建AD+ZjMYB104::BD、AD+ZjMPK3::BD、ZjMYB104::AD+ZjMPK3::BD、ZjMYB104:: BD+ ZjMPK3:: AD的重组载体。利用LiAc法将重组载体质粒同时转化酵母感受态细胞,按1、10-1、10-2浓度梯度培养在三缺选择性培养基(-Leu-Trp-His)中,观察结果。

表2 酵母双杂交引物名称及序列
2 结果
2.1 枣R2R3-MYB亚家族成员的鉴定
在枣基因组中共鉴定出118个R2R3-MYB亚家族成员,参考Ralf等的分类方法[3],将该亚家族进一步分为18个亚组,参考染色体定位结果,分别命名为—(表3)。理化性质分析表明,该亚家族成员编码蛋白的大小和氨基酸数目差异很大,最小的蛋白ZjMYB25由160个氨基酸组成;最大的ZjMYB113由671个氨基酸组成,氨基酸数目处于200—400 aa的蛋白共89个,占比75.42%。该亚家族含有51个酸性蛋白(pH<6.5),20个中性蛋白(pH 6.5—7.5),47个碱性蛋白(pH>7.5),表明该家族蛋白功能具有多样性。
与Ji等[22]前期研究结果相比,本研究新鉴定出21个R2R3-MYB亚家族成员,并发现之前鉴定的(LOC107415212)、(LOC107420595)两个基因不具有典型的R2R3-MYB结构域,因而被剔除。
2.2 枣R2R3-MYB亚家族成员的染色体分布
根据枣全基因组注释信息,利用TBtools软件绘制枣R2R3-MYB基因的染色体定位图。将95个枣R2R3-MYB基因定位到12条染色体上(图1),其中,4号染色体上分布最多,有14个成员(—),占基因总数的14.74%;而9号染色体上成员分布最少,仅含有4个基因(),占4.21%。同时,该家族基因在Chr1、Chr3、Chr4和Chr8上呈簇状分布,可能与该亚家族基因的扩张以及功能分化相关。
2.3 枣R2R3-MYB亚家族成员进化树构建
利用枣R2R3-MYB亚家族成员构建系统进化树(图2),根据蛋白序列相似性,分为18个亚组(I—XVIII)。根据基因组进化分析,桃与枣具有较近的亲缘关系[8],本研究进一步构建了枣与桃R2R3-MYB亚家族成员的系统进化树,也分为18个亚组(图3),与图2分组结果基本一致,说明该亚家族成员在不同物种间具有很好的保守性。

图1 枣R2R3-MYB亚家族成员的染色体定位

表3 枣R2R3-MYB亚家族成员信息

续表3 Continued table 3
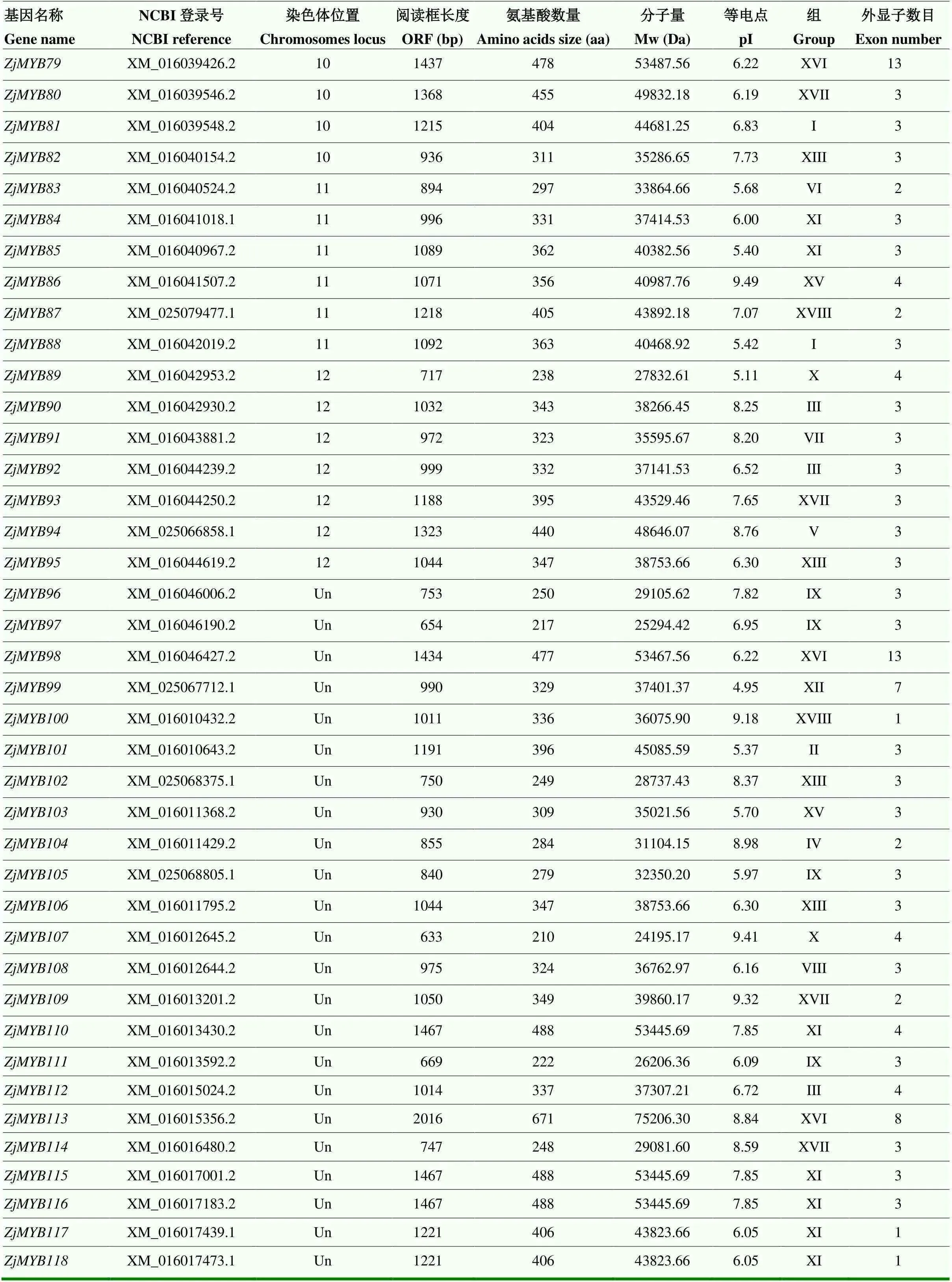
续表3 Continued table 3

图2 枣R2R3-MYB亚家族系统进化树的构建
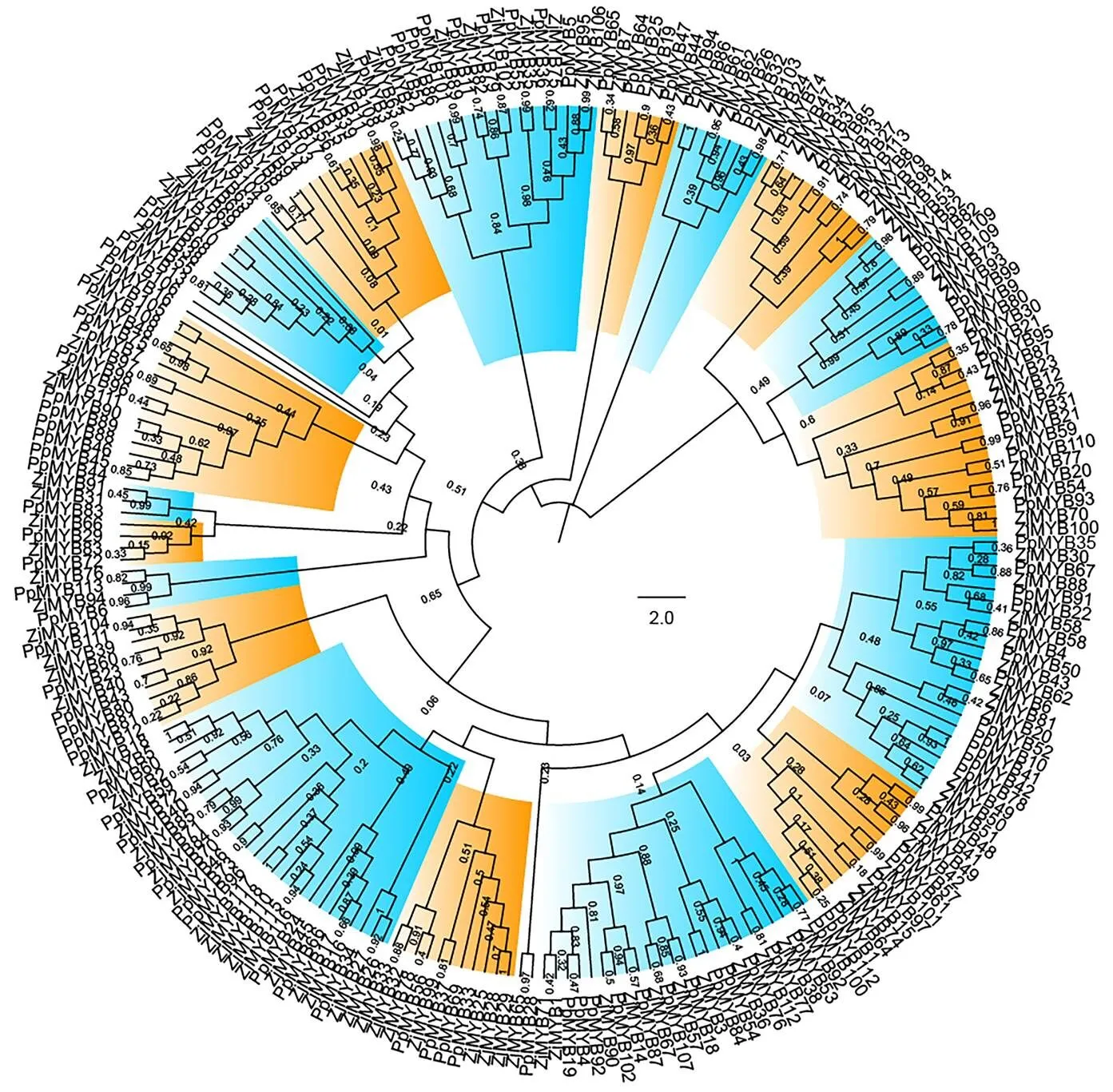
图3 枣与桃R2R3-MYB亚家族成员的系统进化树构建
2.4 枣R2R3-MYB亚家族成员的基因结构分析
利用MEME在线分析软件对枣R2R3-MYB编码
蛋白进行基序分析,设置最大发现基序数目为10,分析结果进一步验证了枣R2R3-MYB亚家族间的进化关系(图4)。其中,motif 1和2为枣R2R3-MYB亚家族最保守的基序,除ZjMYB28ZjMYB117和ZjMYB118外,其他115个成员均含有这两段保守基序。其次保守的基序为motif 3,除ZjMYB13、ZjMYB28、ZjMYB68、ZjMYB69、ZjMYB117和ZjMYB118等6个成员外,其他112个成员均含有该基序。值得注意的是,ZjMYB117和ZjMYB118均不具有以上保守的motif,但分析其同源结构域和基因结构,发现它们均有典型的R2R3-MYB亚家族特征,因此最终归为该亚家族。在该家族进化中,motif 1、2和3结构最为原始,可能具有最保守的功能,推测在此基础上逐步进化出其他基序,并形成了功能上的多样化。

图4 枣R2R3-MYB亚家族基因结构、蛋白保守基序及同源结构域分析
蛋白同源结构域序列特征分析发现,全部成员均具有一个十分保守的MYB超家族蛋白结构域(PLN03091、PLN03212、REB1 superfamily或Myb_DNA-binding),这可能与该亚家族成员的生物学功能密切相关。118个亚家族成员中,9个成员具有1个外显子(7.6%),16个成员具有2个外显子(13.6%),75个成员具有3个外显子(63.6%),12个成员具有4个外显子(10.2%),6个成员具有更多的外显子数目。多数成员具有3个外显子,可能与该亚家族成员功能上的保守性相关。
综上,无论是在保守基序、同源结构域,还是基因结构层面,枣R2R3-MYB亚家族成员间的特征都相对保守。但也有个别成员的基因结构特征与其他成员存在较大差异,如和有13个外显子,具备特殊的Alpha-man_mid superfamily结构域以及更长的氨基酸序列等,更多的外显子数目为这两个基因提供了更多的剪接可能,也说明这两个蛋白在功能上的多样性和特殊性。
2.5 枣R2R3-MYB亚家族重点基因的谱系分析
选取上述6个代表性的枣R2R3-MYB成员进行了谱系分析(图5)。其中,棕色线表示随进化时间的变化具有不同HIs阈值的基因序列数量,蓝色和绿色线分别表示包含这些基因的物种和科的数量。根据谱系折线图中同源事件发生规律,将这6个成员分为两组,第一组包括、、和,该组HI值在0.454—0.540,基因数和物种数都下降,说明发生了直系同源事件;同时和HI值分别为0.498和0.482时,基因数量减少而物种数量没有变化,表明发生了旁系同源事件。第二组包括和,该组在HI值为0.615和0.574时发生了直系同源事件。综合看,该家族成员在进化中,发生了直系和旁系同源事件。
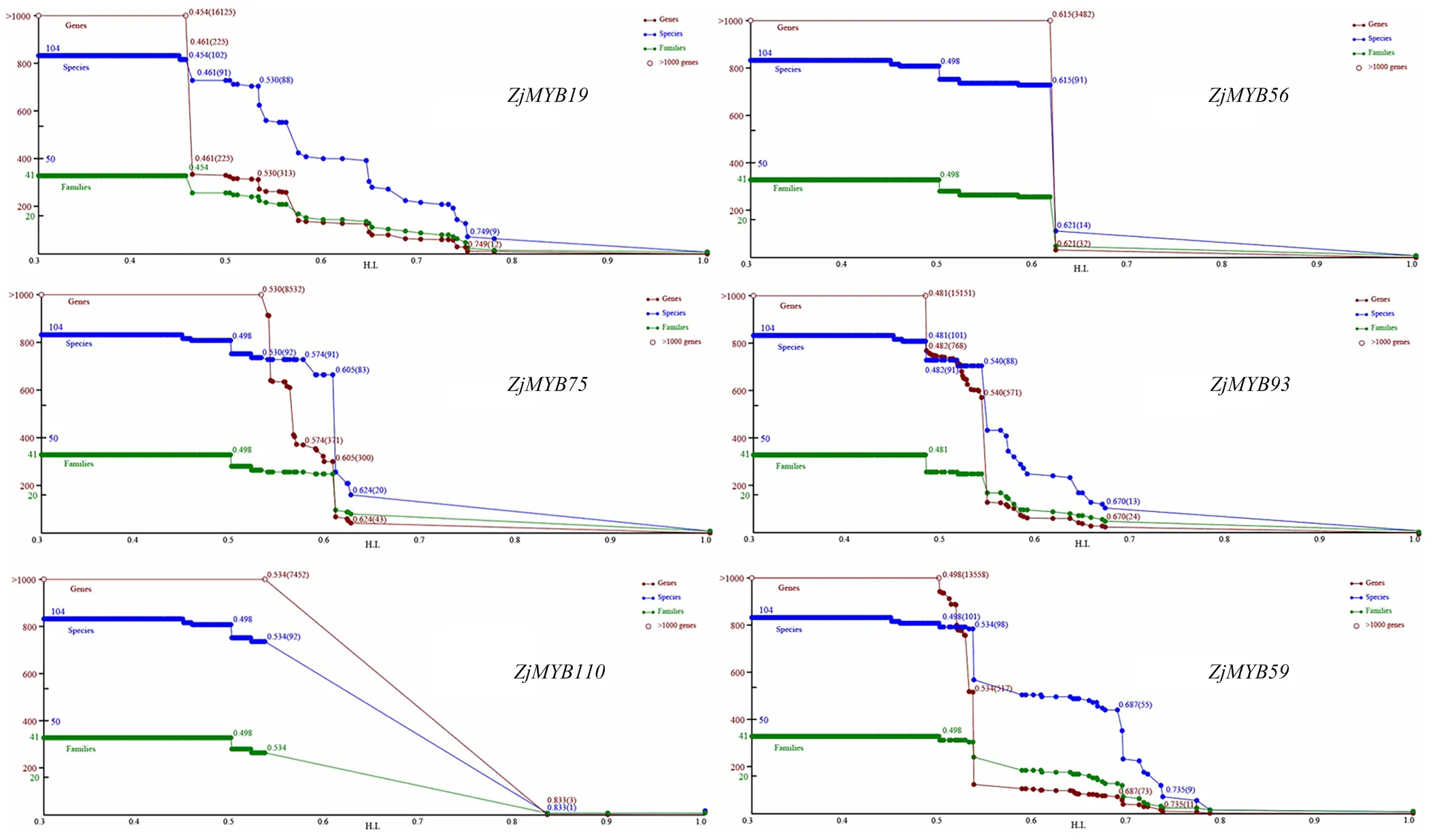
图5 6个枣R2R3-MYB基因谱系分析
2.6 枣R2R3-MYB基因在枣果实发育过程中的表达模式分析
以‘金丝小枣’和‘金魁王’两个品种不同发育时期的枣果(图6-A)为材料,转录组分析筛选出有明显差异表达的14个R2R3-MYB亚家族基因(图6-B),这些基因在两个品种的果实发育中呈现基本一致的表达趋势。
比较基因表达模式发现,总体上可以分为两类,第一类基因表达模式为前期(S1—S4)上调表达,后期(S5—S9)表达明显下降,如、等,表明此类基因在果实发育前期发挥作用,可能参与果实膨大过程;第二类基因的表达模式则相反,在果实发育前期表达量很低,后期逐渐上调,如、等,而后期是果实成熟过程,说明这些基因可能与果实成熟有关。
进一步的实时荧光定量PCR结果显示,在‘金丝小枣’果实发育前期表达量相对较高(图6-C);而表达模式恰好相反(图6-D),在果实发育后期表达量较高,与转录组数据结果一致,也说明可能参与调控枣果后期成熟或色泽发育。
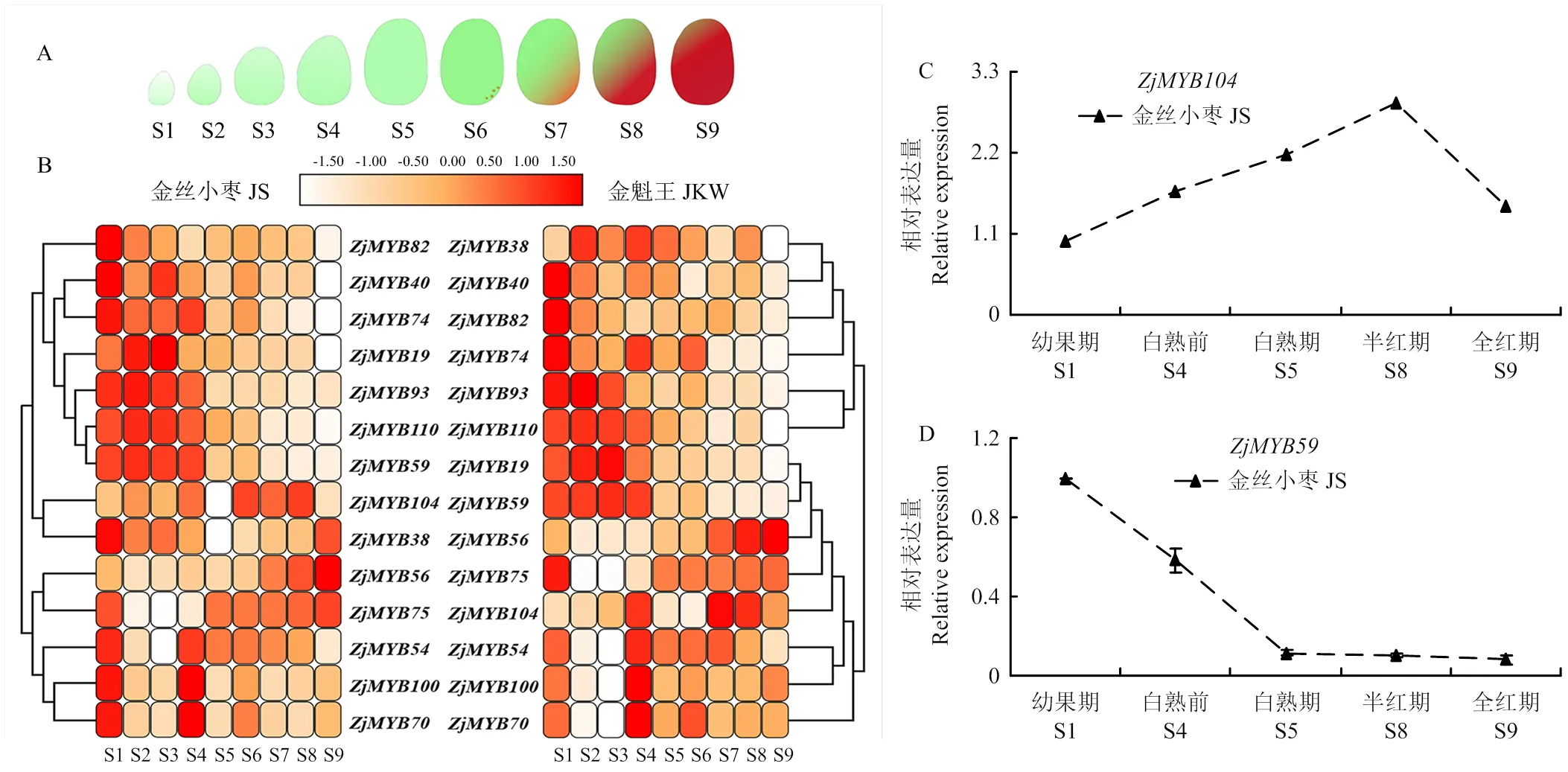
A:枣果不同发育时期;S1:幼果期1;S2:幼果期2;S3:硬核前期;S4:硬核期;S5:白熟期;S6:白熟后期;S7:1/4着色期;S8:半红期;S9:全红期。B:枣果不同发育时期中差异表达的R2R3-MYB基因;C:ZjMYB104在‘金丝小枣’果实不同发育时期的表达;D:ZjMYB59在‘金丝小枣’果实不同发育时期的表达
2.7 ZjMYB104互作蛋白的预测及验证
选取ZjMYB104进行蛋白互作预测(图7-A),结果显示,ZjMYB104与AtMYB44具有较高的同源性,AtMYB44和AtMPK3存在互作关系,推测ZjMYB104和ZjMPK3之间也能发生互作。酵母双杂交试验(图7-B)发现,重组载体AD+ZjMYB104::BD和AD+ ZjMPK3::BD在三缺培养基上不生长,而ZjMYB104:: AD+ZjMPK3::BD、ZjMYB104::BD+ZjMPK3::AD在三缺培养基上能生长,说明ZjMYB104和ZjMPK3确实能发生相互作用。
3 讨论
3.1 R2R3-MYB亚家族基因在不同物种之间具有保守性
在枣基因组中,Ji等[22]鉴定到了99个R2R3-MYB亚家族成员,而本研究鉴定到118个该亚家族成员。近年来,阳桃中鉴定到100个R2R3-MYB亚家族基因[24];梨中发现185个R2R3-MYB亚家族基因[11];梅花基因组中发现106个R2R3-MYB基因[25]。可以看出,枣、阳桃、梅花中该亚家族基因数目相近,但梨中数目较多,猜测可能与梨物种经历两次全基因组复制事件有关。本研究发现枣R2R3-MYB亚家族基因结构相对保守,这与桃、梨、梅花中的研究结果一致,说明该亚家族基因具有较高的系统发育相似性。
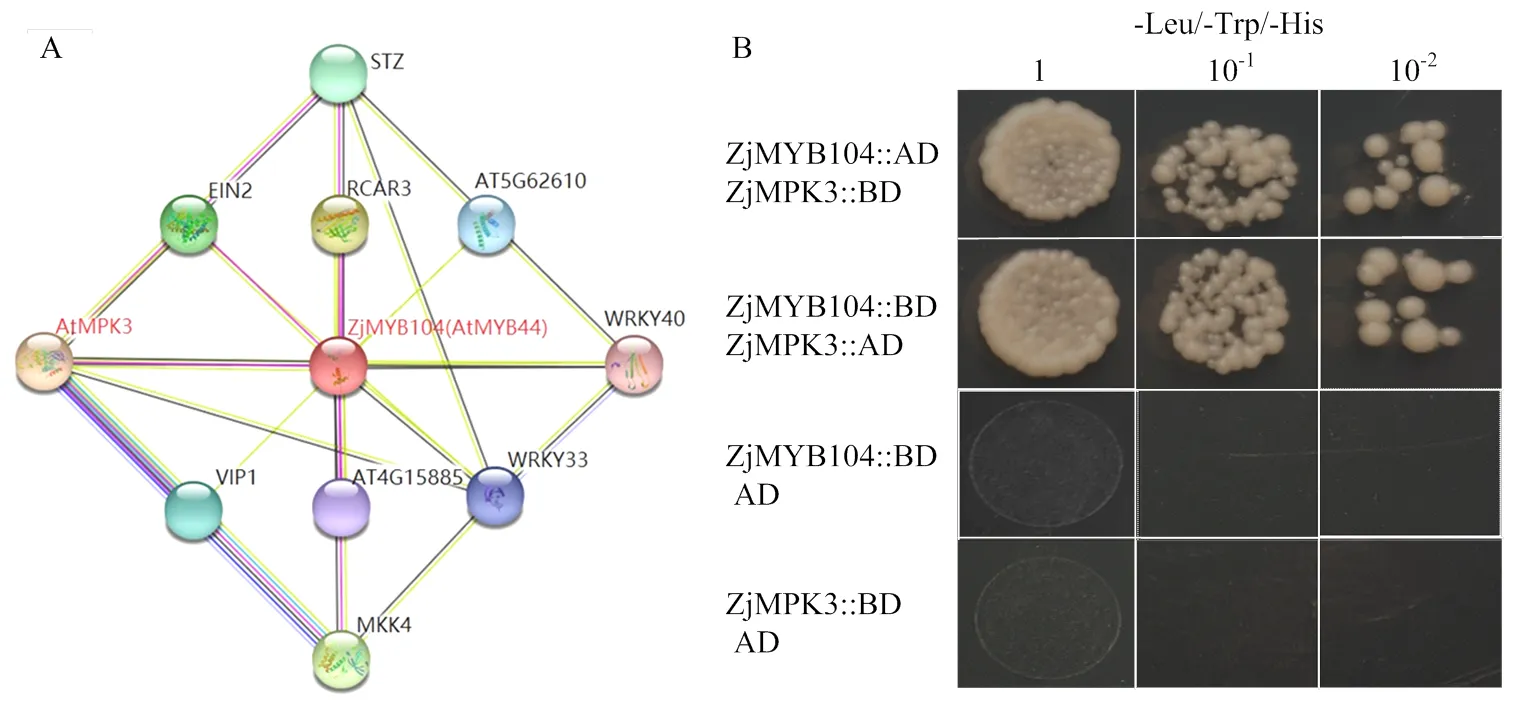
A:ZjMYB104互作蛋白预测;B:ZjMYB104和ZjMPK3酵母双杂试验
3.2 ZjMYB基因可能参与枣果实着色过程
石倩倩[26]研究发现参与调控枣果皮花青苷的合成调控;张琼[27]的研究也表明2个MYB基因正向调控枣果皮着色过程。本研究结果表明,、、等基因在枣果实发育前期(即膨大期)高表达,而、、等基因在果实发育后期表达上升,推测这些基因可能参与枣果实的发育过程,这与之前的研究结果一致。有研究表明,果实发育前期即膨大过程主要涉及一系列黄酮类化合物的产生[28],梨控制查尔酮合成酶基因、黄酮醇合成酶基因的表达,正调控黄酮醇的含量[29];桃能激活黄酮类化合物合成途径,促进黄酮类化合物积累[30];将桃与枣R2R3-MYB基因进行序列比对,发现与序列相似性达87%,而在枣果实发育前期高表达推测部分在枣果发育前期高表达的R2R3-MYB基因可能参与黄酮类化合物的产生、调控果实膨大过程。后续可进一步研究R2R3-MYB基因与果实膨大的关系。
在草莓研究中发现,FaMPK3作为信号传递蛋白激酶,与FaMYB10互作调控下游果实色素等代谢基因的表达情况以及花青素含量等生理指标,超量表达加速草莓果实着色过程[31]。在番茄中,敲除使果实变红提前,过表达发现果实变红延后[32]。而本研究中,在果实发育后期表达上升,且能与ZjMPK3发生互作,推测可能也通过一定方式参与果实着色过程,有待进一步研究。
4 结论
鉴定出118个枣R2R3-MYB亚家族成员,其结构高度保守,筛选出了与果实发育相关的基因、,且可能通过调控花青苷的合成来调节果实着色。ZjMYB104和ZjMPK3存在互作。研究结果为进一步研究R2R3-MYB亚家族基因在枣果实发育中的功能提供了重要的参考和线索。
[1] GRAF T.Myb: A transcriptional activator linking proliferation and differentiation in hematopoietic cells.Centaurus; International Magazine of the History of Science and Medicine, 1992, 2(2): 249-255.doi: 10.1016/s0959-437x(05)80281-3.
[2] DU H, ZHANG L, LIU L, TANG X F, YANG W J, WU Y M, HUANG Y B, TANG Y X.Biochemical and molecular characterization of plant MYB transcription factor family.Biochemistry Biokhimiia, 2009, 74(1): 1-11.doi: 10.1134/s0006297909010015.
[3] WESTON K.Myb proteins in life, death and differentiation.Current Opinion in Genetics & Development, 1998, 8(1): 76-81.doi: 10.1016/ s0959-437x(98)80065-8.
[4] STRACKE R, WERBER M, WEISSHAAR B.The R2R3-MYB gene family in.Current Opinion in Plant Biology, 2001, 4(5): 447-456.doi: 10.1016/s1369-5266(00)00199-0.
[5] MATUS J T, AQUEA F, ARCE-JOHNSON P.Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization acrossandgenomes.BMC Plant Biology, 2008, 8: 83.doi: 10.1186/1471-2229-8- 83.
[6] WILKINS O, NAHAL H, FOONG J, PROVART N J, CAMPBELL M M.Expansion and diversification of theR2R3-MYB family of transcription factors.Plant Physiology, 2008, 149(2): 981-993.doi: 10.1104/pp.108.132795.
[7] 陈哲, 胡福初, 阮城城, 范鸿雁, 郭利军, 张治礼.菠萝R2R3-MYB基因家族鉴定与表达分析.热带作物学报, 2019, 40(10): 1958-1971.
CHEN Z, HU F C, RUAN C C, FAN H Y, GUO L J, ZHANG Z L.Bioinformatics and gene expression analysis of pineapple R2R3-MYB gene family.Chinese Journal of Tropical Crops, 2019, 40(10): 1958-1971.(in Chinese)
[8] 刘孟军, 王玖瑞.新中国果树科学研究70年—枣.果树学报, 2019, 36(10): 1369-1381.
LIU M J, WANG J R.Fruit scientific research in New China in the past 70 years: Chinese jujube.Journal of Fruit Science, 2019, 36(10): 1369-1381.(in Chinese)
[9] DUBOS C, STRACKE R, GROTEWOLD E, WEISSHAAR B, MARTIN C, LEPINIEC L.MYB transcription factors in.Trends in Plant Science, 2010, 15(10): 573-581.doi: 10.1016/j.tplants.2010.06.005.
[10] 赵恩鹏, 成玉富, 杨旭.茄科植物转录因子MYB基因家族的研究进展.分子植物育种, 2020, 19(5): 1522-1530.
ZHAO E P, CHEN Y F, YANG X.Research progress of MYB transcription factors in solanaceae plants.Molecular Plant Breeding, 2020, 19(5): 1522-1530.(in Chinese)
[11] LI X L, XUE C, LI J M, QIAO X, LI L T, YU L A, HUANG Y H, WU J.Genome-wide identification, evolution and functional divergence of MYB transcription factors in Chinese white pear ().Plant and Cell Physiology, 2016, 57(4): 824-847.doi: 10.1093/pcp/ pcw029.
[12] AN J P, LI R, QU F J, YOU C X, WANG X F, HAO Y J.R2R3-MYB transcription factor MdMYB23 is involved in the cold tolerance and proanthocyanidin accumulation in apple.The Plant Journal, 2018, 96(3): 562-577.doi: 10.1111/tpj.14050.
[13] CHEN Y H, YANG X Y, HE K, LIU M H, LI J G, GAO Z F, LIN Z Q, ZHANG Y F, WANG X X, QIU X M, SHEN Y P, ZHANG L, DENG X H, LUO J C, DENG X W, CHEN Z L, GU H Y, QU L J.The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family.Plant Molecular Biology, 2006, 60(1): 107-124.doi: 10.1007/s11103-005- 2910-y.
[14] MA D, REICHELT M, YOSHIDA K, GERSHENZON J, CONSTABEL C P.Two R2R3-MYB proteins are broad repressors of flavonoid and phenylpropanoid metabolism in poplar.The Plant Journal, 2018, 96(5): 949-965.doi: 10.1111/tpj.14081.
[15] 朱婷婷, 梁东, 夏惠.R2R3-MYB调控果实花色苷合成的研究进展.基因组学与应用生物学, 2016, 35(4): 985-991.
ZHU T T, LIANG D, XIA H.Progress in the study of the regulation of anthocyanin synthesis by R2R3-MYB in fruits.Genomics and Applied Biology, 2016, 35(4): 985-991.(in Chinese)
[16] 靳亚忠, 齐娟, 何淑平, 吴兴彪, 张天一, 王雪娇, 胡文曲, 卢朝晖, 张鹏.R2R3-MYB转录因子调控果蔬品质形成的研究进展.农业生物技术学报, 2021, 29(2): 364-374.
JIN Y Z, QI J, HE S P, WU X B, ZHANG T Y, WANG X J, HU W Q, LU Z H, ZHANG P.Recent advance of R2R3-MYB transcription factors regulating the formation of fruit and vegetable qualities.Journal of Agricultural Biotechnology, 2021, 29(2): 364-374.(in Chinese)
[17] 李小溪.葡萄果实莽草酸途径和类黄酮代谢协同调节机制的研究[D].北京: 中国农业大学, 2016.
LI X X.Collaborative expression mechanism between shikimate pathway and flavonoid metabolism [D].Beijing: China Agricultural University, 2016.(in Chinese)
[18] WANG Z G, MENG D, WANG A D, LI T L, JIANG S L, CONG P H, LI T Z.The methylation of the PcMYB10 promoter is associated with green-skinned sport in max red Bartlett pear.Plant Physiology, 2013, 162(2): 885-896.doi: 10.1104/pp.113.214700.
[19] 周厚成, 李刚, 赵霞, 王子成, 郭蔼光.草莓果实不同发育阶段抑制差减文库(SSH)的构建及相关基因的表达分析.农业生物技术学报, 2013, 21(6): 641-649.
ZHOU H C, LI G, ZHAO X, WANG Z C, GUO A G.Construction of suppression subtractive hybridization(SSH) cDNA library from two developmental stages of×fruit and expression analysis of related genes.Journal of Agricultural Biotechnology, 2013, 21(6): 641-649.(in Chinese)
[20] WANG H, ZHANG H, YANG Y, LI M F, ZHANG Y T, LIU J S, DONG J, LI J, BUTELLI E, XUE Z, WANG A M, WANG G X, MARTIN C, JIN W M.The control of red colour by a family of MYB transcription factors in octoploid strawberry () fruits.Plant Biotechnology Journal, 2020, 18(5): 1169-1184.
[21] ESPLEY R V, BRENDOLISE C, CHAGNÉ D, KUTTY-AMMA S, GREEN S, VOLZ R, PUTTERILL J, SCHOUTEN H J, GARDINER S E, HELLENS R P, ALLAN A C.Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples.The Plant Cell, 2009, 21(1): 168-183.doi: 10.1105/tpc.108.059329.
[22] JI Q, WANG D W, ZHOU J, XU Y L, SHEN B Q, ZHOU F.Genome-wide characterization and expression analyses of the MYB superfamily genes during developmental stages in Chinese jujube.PeerJ, 2019, 7(1): 6353.
[23] CHEN C J, CHEN H, ZHANG Y, THOMAS H R, FRANK M H, HE Y H, XIA R.TBtools: an integrative toolkit developed for interactive analyses of big biological data.Molecular Plant, 2020, 13(8): 1194-1202.doi: 10.1016/j.molp.2020.06.009.
[24] 赵亚梅, 陈蕾, 吴春梅, 秦思, 翟俊文, 任惠, 吴沙沙.阳桃R2R3-MYB家族成员鉴定及其在木质素合成过程中的表达.植物遗传资源学报, 2021(6): 1-19.
ZHAO Y M, CHEN L, WU C M, QIN S, ZHAI J W, REN H, WU S S.Identification of the R2R3-MYB family members inand their expression involved in lignin biosynthesis.Journal of Plant Genetic Resources, 2021(6): 1-19.(in Chinese)
[25] 周婵, 陈超, 左静, 晏晓兰, 张俊卫, 包满珠.梅R2R3型MYB转录因子的鉴定及低温胁迫下的表达分析.北京林业大学学报, 2015, 37(S1): 61-65.
ZHOU C, CHEN C, ZUO J, YAN X L, ZHANG J W, BAO M Z.Identification and expression analysis of R2R3-MYB transcription factors inunder cold treatment.Journal of Beijing Forestry University, 2015, 37(S1): 61-65.(in Chinese)
[26] 石倩倩.枣果实色泽性状形成的分子机制研究[D].杨凌: 西北农林科技大学, 2019.
SHI Q Q.Molecuar mechanism of the formation of fruit pigment in jujube fruits [D].Yangling: North West Agriculture and Forestry University, 2019.(in Chinese)
[27] 张琼.枣着色过程中果皮结构及色素积累相关组分研究[D].保定: 河北农业大学, 2020.
ZHANG Q.Analysis of structure and components related to pigment accumlation during jujube coloring [D].Baoding: Hebei Agricultural University, 2020.(in Chinese)
[28] 陈昕, 陈蕊红, 黄建.枣果实不同发育阶段蛋白质组动态研究.西北林学院学报, 2019, 34(4): 82-90.doi: 10.3969/j.issn.1001-7461.2019.04.12.
CHEN X, CHEN R H, HUANG J.Comparative proteomic analysis on Chinese jujube fruit at different developmental and ripening stages.Journal of Northwest Forestry University, 2019, 34(4): 82-90.doi: 10.3969/j.issn.1001-7461.2019.04.12.(in Chinese)
[29] ZHAI R, ZHAO Y X, WU M, YANG J, LI X Y, LIU H T, WU T, LIANG F F, YANG C Q, WANG Z G, MA F W, XU L F.The MYB transcription factor PbMYB12b positively regulates flavonol biosynthesis in pear fruit.BMC Plant Biology, 2019, 19(1): 85.doi: 10.1186/s12870-019-1687-0.
[30] RAHIM M A, RESENTINI F, DALLA VECCHIA F, TRAINOTTI L.Effects on plant growth and reproduction of a peach R2R3-MYB transcription factor overexpressed in tobacco.Frontiers in Plant Science, 2019, 10: 1143.doi:10.3389/fpls.2019.01143.
[31] 韩瑜.信号系统在草莓果实发育和成熟中的作用及机理分析[D].北京: 中国农业大学, 2015.
HAN Y.The function and mechanism analysis ofsignaling system during fruit development and ripening in[D].Beijing: China Agricultural University, 2015.(in Chinese)
[32] 叶婷.番茄通过与SlSnRK1互作调控果实成熟机理研究[D].武汉: 华中农业大学, 2020.
YE T.Mechanisms ofregulate fruit ripening via interacting with SlSnRK1 in tomato [D].Wuhan: Huazhong Agricultural University, 2020.(in Chinese)
Identification of R2R3-MYB Subfamily in Chinese Jujube and Their Expression Pattern During the Fruit Development
LI ShiJia, LÜ ZiJing, ZHAO Jin*
College of Life Sciences, Hebei Agricultural University, Baoding 071000, Hebei
【Objective】As one of the largest gene families in plants, MYB family plays an important role in plant growth and fruit development.This study would identify the members of the R2R3-MYB subfamily of Chinese jujube and analyze their bioinformatics and expression patterns, aiming to provide the reference for further exploring their function in jujube growth, especially in fruit development.【Method】The R2R3-MYB genes ofwere used as probes, and this subfamily members in the jujube genome were screened out by BLAST and HMMER.Based on the Plant Transcription Factors Database, Uniprot, Gcorn Plant and other databases, R2R3-MYB genes were analyzed by MEGA-X, ExPASy, TBtools, and PheatMap.Transcriptome data and real-time quantitative PCR (qRT-PCR) were used to evaluate the expression of these genes, and the interaction proteins of key candidate proteins were predicted and verified.【Result】In this study, 118 R2R3-MYB subfamily members were identified in jujube genome, all of which had a typical MYB domain.They were divided into 18 subgroups, named-.The number of amino acids encoded by the genes ranged from 200 to 400.Among them, 95 genes were located on 12 chromosomes, and which on Chr.4 was the most widely distributed.The phylogenetic analysis showed that orthologous and paralogous events occurred during the evolution of these genes.By transcriptomic and qRT-PCR analysis, some R2R3-MYB genes were involved in the fruit development. In particular,andmight participate in fruit enlargement and color development.Protein interaction prediction and yeast two-hybrid test confirmed that ZjMYB104 and ZjMPK3 were interacted each other.【Conclusion】118 members of the R2R3-MYB subfamily members were identified and distributed on 12 chromosomes.The structure of these subfamily members was highly conservative.The orthologous and paralogous events occurred in jujube genome evolution and the candidate genes closely related to fruit development were screened out.This study provided clues and references for further functional analysis of R2R3-MYB subfamily in jujube.
Chinese jujube; R2R3-MYB subfamily; bioinformatics analysis; expression analysis; fruit development; proteins interaction
2021-06-13;
2021-08-10
国家重点研发计划(2018YFD1000607)、河北农业大学大学生创新创业训练计划资助项目(s202010086002)
李世佳,E-mail:1193326557@qq.com。通信作者赵锦,E-mail:zhaojinbd@126.com
(责任编辑 赵伶俐)
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
家庭医学·下半月(2019年6期)2019-08-16
电影(2019年3期)2019-04-04
阅读(低年级)(2018年11期)2018-05-14
少儿科学周刊·少年版(2017年3期)2017-06-29
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
奥秘(2015年4期)2015-09-10
中学生物学(2008年12期)2008-12-27
