
激活泛素-蛋白酶体系对脊髓小脑共济失调7型细胞模型中共济失调蛋白-7的作用研究*
2022-04-24 02:38苏莹珍贺启莲帅红艳XinYu
成都医学院学报 2022年2期
刘 冰,付 云,苏莹珍,贺启莲,帅红艳,Xin Yu △
1.大理大学 基础医学院(大理671000);2.大理大学 代谢性疾病转化医学研究院(大理671000);3.昆明学院 医学院(昆明 650214);4.大理大学 护理学院(大理671000)
脊髓小脑共济失调7型(spinocerebellar ataxia type 7,SCA7)是常染色体显性遗传的神经退行性疾病,临床表现为视网膜、小脑和脑干中的神经元退行性死亡[1]。该疾病表现为编码的失调共济蛋白-7(ataxin-7,ATXN7)蛋白中多聚谷氨酰胺(polyglutamine,PolyQ)结构域的扩增[2],这种扩增是由ATXN7基因中不稳定的CAG重复序列发生扩增突变导致。除SCA7外,另外8种神经退行性疾病的发病机制与SCA7相似,包括亨廷顿病、齿状核苍白球萎缩、脊髓延髓肌萎缩以及脊髓小脑共济失调1~3型、6型和17型,统称为PolyQ疾病[3]。PolyQ蛋白形成的单体、寡聚体和内含体(聚合体)等在细胞中以动态平衡形式存在[4],但各个突变蛋白结构引起细胞毒性的分子机制尚存争议。因此,研究错误折叠蛋白的有效清除方法对开发治疗PolyQ疾病的潜在方案具有重要意义。
泛素-蛋白酶体系(ubiquitin-proteosome system,UPS)是哺乳动物体内调控蛋白质降解的重要途径[5]。如何特异性激活UPS一直是本领域研究的技术瓶颈,目前已知UPS功能的激活机制包括:1)增强蛋白水解能力;2)刺激泛素化过程;3)抑制泛素分解酶。研究[5]发现,刚果红衍生物WSP774和WSP677能增强蛋白酶体对受感染细胞中朊病毒蛋白PrPSc的降解,减轻PrPSc对蛋白酶体的抑制作用。萝卜硫素能明显增强小鼠大脑、外周组织中UPS和自噬的活性,并促进泛素化蛋白在体内转化,减少PolyQ蛋白的聚集[6]。而1-[1-(4-氟苯基)-2,5-二甲基-1H-吡咯-3-基]-2-(1-吡咯烷基)乙酮{1-[1-(4-fluorophenyl)-2,5-dimethylpyrrol-3-yl]-2-pyrrolidin-1-yle-thanone,IU-1}是目前唯一通过抑制泛素特异性蛋白酶(ubiquitin specific protease 14,USP14)对多泛素化底物进行去泛素化,来增强蛋白酶体活性的小分子[7]。IU-1是泛素分解酶USP14的可逆性抑制剂[8],但目前IU-1对于UPS的激活作用是否能够影响PolyQ疾病尚无确切研究。
以往在UPS参与PolyQ突变蛋白降解作用的研究中,往往缺少激活UPS对促进PolyQ蛋白聚合体的影响[9]。本研究通过激活和抑制UPS活性,观察其对ATXN7累积与聚合体水平的变化及对细胞毒性的影响,探索UPS对PolyQ突变蛋白的清除机制。
1 材料与方法
1.1 细胞和质粒
大鼠肾上腺髓质嗜铬细胞瘤细胞系PC12和质粒pTRE-tight基因调控系统(tetracycline tet-off system,Tet-off)均购自美国Clontech公司。
1.2 实验材料
盐酸多西霉素(doxycline, Dox)、伯克改良伊格尔培养基(Dulbecco modified Eagle medium, DMEM)、IU-1、环氧甲酮四肽(epoxomycin,Epox)、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、二硫苏糖醇(dithiothreitol,DTT)、蛋白酶抑制剂混合物、4′,6-二脒基-2-苯基吲哚(4′, 6-diamidino-2-phenylindole,DAPI)、辣根过氧化物酶标记的抗体均购自美国Sigma公司;RIPA裂解液购自美国Millipore公司;脱氧核糖核酸酶(deoxyribonudease Ⅰ,DNaseI)(EN0521)购自美国Fermentas公司;Bradford蛋白浓度检测试剂盒购自美国Bio-Rad公司;2-(4-碘苯)-3-(4-硝基苯)-5-(2,4-二磺基苯)-2H-四氮唑钠盐[2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium sodium salt,WST-1]细胞活性检测试剂盒购自美国Clontech公司;ATXN7抗体、肌动蛋白(Actin)抗体(SC-1616)均购自美国SantaCruz公司;聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜购自英国Whatman公司。
1.3 实验方法
1.3.1 稳定表达ATXN7突变蛋白细胞系的培养稳定表达ATXN7蛋白的PC12细胞系及质粒构建方法参照本课题组前期方法[10]:分别将编码Q10和Q65的DNA片段插入到pTRE-tight off质粒的多克隆位点中,再将构建成功的质粒分别转染到PC12细胞中,经筛选获得稳定表达ATXN7蛋白的两种不同类型细胞,分别命名为ATXN7-Q10和ATXN7-Q65细胞,其中ATXN7-Q10细胞表达非突变蛋白ATXN7,ATXN7-Q65细胞表达突变蛋白ATXN7。PC12细胞系在含10%马血清、5%胎牛血清、100 mg/L G418、100 U/mL氨苄青霉素、100 mg/L链霉素、100 mg/L潮霉素、1 mg/L Dox的DMEM培养基中,置于37 °C,5% CO2的细胞培养箱中培养(图1)。
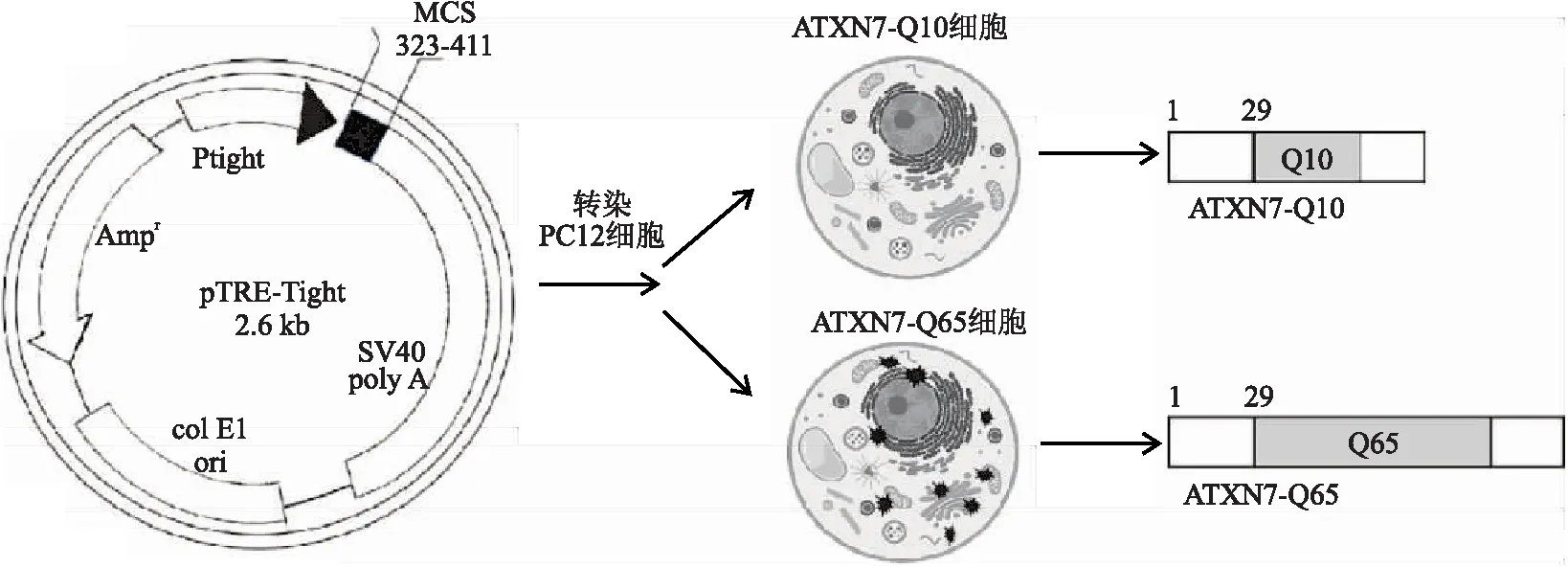
图1 稳定表达ATXN7-Q10和ATXN7-Q65的PC12细胞系构建
1.3.2 ATXN7蛋白的诱导表达 不添加Dox的Tet-off专用DMEM培养基培养PC12细胞,并在后续不同诱导时间(0、1、3、6、9、12 d)收集细胞并裂解,分别检测ATXN7-Q10和ATXN7-Q65细胞中的可溶性蛋白及其聚合体水平。在ATXN7蛋白降解的研究中,首先在已诱导表达目的蛋白12 d的PC12细胞培养基中添加Dox以抑制目的蛋白表达,继续培养细胞并检测24 h内不同时间点ATXN7可溶性蛋白及其聚合体表达水平的变化。
1.3.3 蛋白质印迹技术及斑点过滤杂交法检测可溶性ATXN7及其聚合体水平 ATXN7-Q10和ATXN7-Q65细胞经RIPA裂解后,使用含有蛋白酶抑制剂混合物和苯甲基磺酰氟的细胞裂解缓冲液裂解细胞并收集上清液,考马斯亮蓝法测蛋白浓度,样品经丙烯酰胺凝胶电泳后,分别与一抗ATXN7抗体(1∶700)、Actin(1∶500)、二抗辣根过氧化物酶标记的抗体(1∶2 000)进行特异性抗体标记,做蛋白质印迹技术分析。用斑点过滤杂交法法分析斑点印记强度,具体操作为将细胞裂解液沉淀部分经磷酸盐缓冲盐溶液洗涤2次,重悬于50 μL DNaseI反应缓冲液中,并加入4 U的DNaseI反应缓冲液,37 °C消化1 h,获得不溶性沉淀(内含体或多聚体)。向沉淀中加入2%的SDS和1×105μmol/L的DTT,95 °C变性处理5 min后,用0.22 μm孔径PVDF膜(Bio-Rad 微量过滤器)经真空过滤后进行特异性抗体标记。采用Super Signal West Pico试剂盒进行目的蛋白的显影,并用ChemiDoc XRS+ 成像系统(Bio-Rad)分析蛋白信号强度,最后用凝胶式成像系统软件(Bio-Rad,美国)进行定量分析。
1.3.4 WST-1细胞活性试验和DAPI染色法检测细胞凋亡 分别将ATXN7-Q10和ATXN7-Q65细胞以5×104个/孔浓度接种到96孔板中,并分别在0、1、3、6、9、12 d时,利用WST-1细胞活性检测试剂盒测定细胞存活率。同时重复上述方法,细胞接种于内置明胶包被盖玻片的6孔板内,分别诱导ATXN7-Q10和ATXN7-Q65细胞表达ATXN7蛋白0、6、12 d后,经4%多聚甲醛固定并进行DAPI染色(具体步骤按照产品说明操作)。在荧光显微镜(激发波长340~380 nm,发射波长435~485 nm)下随机选取10个视野观测细胞凋亡小体,计数凋亡细胞并计算细胞凋亡率。在UPS活性对ATXN7-Q65细胞表达ATXN7突变蛋白影响的研究中,向ATXN7-Q65细胞培养基中分别添加IU-1(5×104μmol/L)或Epox(0.2 μmol/L),以未添加试剂的ATXN7-Q65细胞为对照组,持续培养并在各时间点(0、1、3、6、9、12 d)用WST-1试剂盒检测细胞存活率。
1.4 统计学方法

2 结果
2.1 突变蛋白ATXN7的聚合性及细胞毒性
蛋白质印迹技术结果显示,在构建的两组神经细胞模型中,随着诱导表达时间的延长,在ATXN7-Q10细胞中,可溶性ATXN7蛋白的表达明显升高;在ATXN7-Q65细胞中,可溶性ATXN7蛋白的表达明显升高,并伴有ATXN7聚合体的增加(图2、表1)。
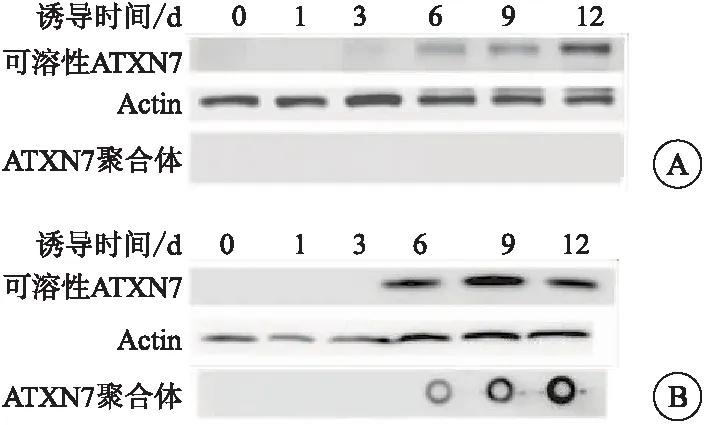
图2 诱导表达ATXN7蛋白对两组细胞可溶性ATXN7
表1 蛋白质印迹技术检测两组细胞可溶性ATXN7蛋白水平
WST-1细胞活性实验和DAPI染色显示,PC12细胞经诱导0~3 d后,细胞存活率未出现明显改变,经诱导6 d后,ATXN7-Q65细胞存活率明显降低,凋亡细胞的比率增加(P<0.05);经诱导12 d后,细胞存活率进一步降低,该比率与细胞凋亡率基本接近(表2~3)。在ATXN7-Q10细胞中,随着诱导时间延长,细胞存活无明显下降,细胞凋亡差异无统计学意义(P>0.05)。
表2 WST-1细胞增殖活性检测结果
表3 DAPI染色细胞凋亡检测结果
2.2 UPS活性变化对ATXN7蛋白水平的影响
在两组细胞加入Dox抑制剂结果显示,与ATXN7-Q10细胞相比,ATXN7-Q65细胞的可溶性ATXN7蛋白的降解相对缓慢,并于4、8 h后,ATXN7-Q65细胞的ATXN7蛋白水平明显高于ATXN7-Q10细胞(P<0.05)(图3、表4)。
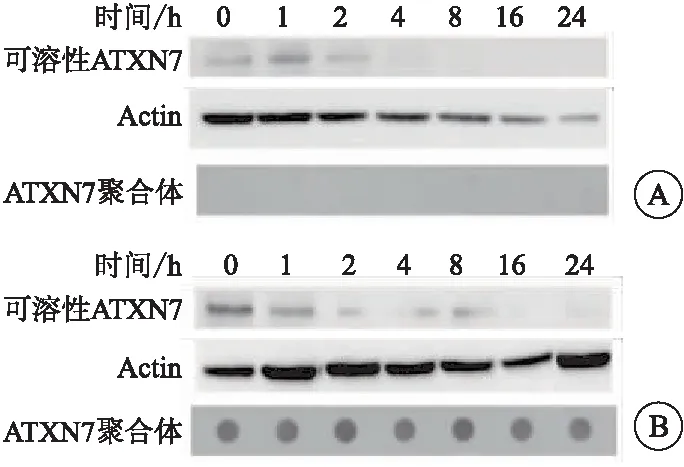
图3 加入Dox抑制剂后对两组细胞可溶性ATXN7
表4 蛋白质印迹技术检测两组细胞可溶性ATXN7蛋白水平
2.3 UPS活性变化对ATXN7-Q65细胞毒性的影响
经特异性UPS激活剂IU-1和抑制剂Epox持续处理ATXN7-Q65细胞后结果显示,与对照组相比,IU-1组激活后第12天ATXN7-Q65细胞存活率升高(P<0.05);相反,抑制UPS活性12 d后,Epox组ATXN7-Q65细胞死亡率明显增高(P<0.001)(表5)。
表5 WST-1检测UPS活性变化后ATXN7-Q65细胞的增殖活性
3 讨论
UPS是真核细胞中蛋白质降解的重要途径之一,主要通过降解泛素化的靶蛋白参与短半衰期蛋白或多肽的降解,而自噬主要参与蛋白聚合体等相对体积较大的底物降解[11]。研究[12]表明,突变的PolyQ蛋白对UPS的清除有抵抗力,甚至由于突变蛋白聚合体的形成会抑制UPS对底物的降解,进而破坏UPS的活性,因此降低ATXN7突变蛋白的聚合程度以及增加UPS的降解能力,均可改善由PolyQ蛋白异常扩增引起的细胞毒性。但是本研究发现,在PolyQ序列扩增形成多聚体后,UPS依然能降低PolyQ突变蛋白聚合体的水平。这可能是由于UPS途径的增强有效增加突变蛋白单体的降解,引起突变蛋白聚合体与可溶性单体之间的化学平衡发生变化,使平衡从聚合体向单体方向转变,最终引起突变蛋白聚合体水平降低[13];同时也可能是由于UPS活性的增强,降低突变蛋白对UPS蛋白降解功能的损伤,进而加速突变蛋白单体的清除效率[14]。
由此可见,如果能有效刺激UPS活性,延缓突变蛋白聚合的速度和程度,可能对解决由PolyQ蛋白突变及聚合所引起的神经退行性疾病具有重要意义。本研究在诱导表达ATXN7突变蛋白的PC12细胞模型中,利用IU-1和Epox处理细胞并检测UPS活性变化对突变蛋白降解和对细胞毒性的影响。与诱导表达ATXN7-Q10的PC12细胞相比,ATXN7-Q65细胞可引起蛋白聚合体的形成并产生细胞毒性。在ATXN7-Q65细胞降解实验中发现,扩增的PolyQ序列能明显增加可溶性突变蛋白及其聚合体的稳定性,抵抗细胞的降解作用。既往研究[11]结果显示,ATXN7突变蛋白的降解受UPS活性调控。IU-1激活UPS后,能促进可溶性突变蛋白ATXN7及其聚合体的清除,并有效缓解由PolyQ突变蛋白引起的细胞毒性;相反Epox抑制UPS的活性后,PC12细胞中PolyQ突变蛋白的水平增加,细胞存活率降低。
鉴于上述发现,有效促进UPS途径,可能是缓解PolyQ疾病的新途径。增强UPS的降解能力既可通过改变UPS活性,也可通过改善被降解底物的活性来提高降解效率。有研究[15]表明,利用人工设计的小分子多肽QBP1抑制PolyQ突变蛋白的聚合,能有效降低PolyQ突变蛋白在动物体内的毒性。目前开发的UPS激活剂其特异性及抑制效果一直存在争议,因为这类激活剂往往除了有效地激活UPS外,还会引起自噬功能的激活,进而无论是突变蛋白单体还是聚合体的降解都会得到增强[16]。然而不论是单独激活UPS还是附带对自噬功能的激活,其对于有毒性突变蛋白各种成分的清除作用都对细胞的存活表现出保护作用。但有效激活UPS活性相比对自噬功能的激活效果更优越。有研究[17]发现,PolyQ蛋白在形成聚合体之前会形成错误折叠的单体和寡聚体,其表现出比聚合体更强的细胞毒性。而UPS的激活能有效清除形成聚合体之前的多种蛋白结构形式,在形成聚合体之前降低突变蛋白水平。IU-1作为UPS的特异性小分子激活剂,能对蛋白酶体发挥可逆的选择性抑制作用,已被开始应用于细胞研究,但其不足之处表现为IU-1激活UPS活性的能力相对较弱[18],因此进一步优化其结构,增强其蛋白降解功能,对于防治突变蛋白的累积和聚集具有十分重要的意义。
综上所述,在PC12细胞系中,UPS途径在PolyQ突变蛋白的清除机制中起关键作用,UPS活性的增强对PolyQ突变蛋白单体及蛋白聚合体的清除具有促进作用;同时IU-1作为UPS的激活剂能促进可溶性ATXN7突变蛋白及其聚合体的降解,明显降低由PolyQ突变蛋白产生的细胞毒性。本研究为进一步探索利用UPS激活剂缓解和治疗由PolyQ蛋白突变引起的神经退行性疾病提供了新思路和理论依据。本研究不足之处在于未探讨在体内环境下,UPS对突变蛋白的清除作用,因此尚需进一步的体内实验来验证其作用效果及机制。
猜你喜欢
临床外科杂志(2022年8期)2022-11-23
南京中医药大学学报(社会科学版)(2022年1期)2022-11-23
四川蚕业(2022年2期)2022-11-19
中国药理学与毒理学杂志(2022年7期)2022-10-17
生物化学与生物物理进展(2022年8期)2022-08-20
科学导报(2022年44期)2022-07-25
当代水产(2022年6期)2022-06-29
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
江苏农业科学(2017年19期)2017-11-22
