
基于SDMs-toolbox预测气候变暖趋势下辽宁省三裂叶豚草潜在入侵区域
2022-04-15 07:13王迪曲波周斌张依然刘智
生态科学 2022年2期
王迪, 曲波, 周斌, 张依然, 刘智
基于SDMs-toolbox预测气候变暖趋势下辽宁省三裂叶豚草潜在入侵区域
王迪1,2, 曲波3,*, 周斌4, 张依然1, 刘智1
1. 沈阳环境科学研究院,辽宁省城市生态重点实验室, 沈阳 110161 2. 中国科学院生态环境研究中心, 北京 100085 3. 沈阳农业大学生物科学技术学院, 辽宁省全球变化与生物入侵重点实验室, 沈阳 110866 4. 辽宁省生态气象和卫星遥感中心, 沈阳 110866
为明确气候变暖对三裂叶豚草的潜在适生区域的影响并揭示三裂叶豚草潜在扩散路径及规律, 文章选取辽宁省作为研究区域, 模拟气候变暖条件下三裂叶豚草在2050年潜在分布范围。研究结果表明, 拟合温室气体排放量低的情况下(RCP4.5), 三裂叶豚草扩散面积相比2019年增加了10919.9 km2, 其中中度和高度适生区分别增加了3137.32 km2和7782.58 km2, 面积占比分别增加了1.24%和3.07%; 拟合温室气体排放量高的情况下(RCP8.5), 三裂叶豚草中高度适宜区面积相比2019年增加了6932.2 km2, 中度和高度适宜区面积分别增加了2554.81 km2和4377.11 km2, 面积占比增加了1.01%和1.73%, 说明高温室气体排放量并没有加速三裂叶豚草的快速扩张。高低温室气体排放量情况下, 扩张方向大致相同, 均表现为为西北和西南两个方向。其一为以中部平原地区为中心向西北方向的锦州、阜新扩张, 由原来的低度适生区转变为高度适生区, 西部沿大小凌河方向扩张; 其二为沿辽宁中部河流向西南方向扩张, 营口盘锦沿海地区扩散趋势明显。刀切法检验结果表明高程对三裂叶豚草生境适宜度的贡献率最高, 占47.4%, 且高程处于0-200m的范围内、坡度处于0度左右的区域三裂叶豚草适宜性指数较高, 说明三裂叶豚草适合分布在高程较低、坡度较缓的平原区域。该研究结果为三裂叶豚草在辽宁省乃至全国的入侵风险预警防治提供技术支撑。
入侵物种; 预测; SDMs-toolbox; MaxEnt模型; 风险评估
0 前言
三裂叶豚草()又名大破布草, 原产于北美洲, 一年生粗壮草本植物, 被列入中国第二批外来入侵物种名单。1930年, 在我国辽宁铁岭地区首次发现, 并以种子量大且较轻等生物学特征迅速在辽宁省各市扩散蔓延。辽宁省位于我国东部的季风区, 夏季以东南风为主、冬季以东北风为主, 以风媒传播为主的三裂叶豚草随着风向形成了吉林-黑龙江-内蒙古和河北-山西两条扩散路径, 成为三裂叶豚草在我国传播和扩散的点源区域。三裂叶豚草从北美洲入侵到我国辽宁等地, 逃避了原有生境天敌的制约, 又因其植株味苦且适口性差导致牲畜不食, 以上两点因素很大程度上助长了其生长和蔓延。三裂叶豚草生长繁茂, 耗肥吸水能力强, 易造成土壤干旱贫瘠, 严重影响作物的生长, 促使共生小麦、大麦、大豆等作物减产甚至绝收[1], 花粉也是引起人体过敏哮喘等症状的重要过敏源之一。因此, 三裂叶豚草的发现、防控和清除等工作至关重要。
全球变暖的趋势下, 可能会加速外来植物的侵入、定殖、建群和扩散, 促进外来入侵植物在侵入地定居[2], 进而影响植被群落生物多样性、物种分布和栖息地环境。国内研究表明, 外来入侵植物互花米草()目前在我国的分布区域较小, 但在气候变暖的条件下适生区域明显增加, 且适生面积在RCP4.5情景下大于RCP8.5[3]。唐瑶等研究结果表明, 外来入侵植物刺萼龙葵()、腺龙葵()、北美刺龙葵()和银毛龙葵()在气候变暖的情况下, 在我国的适生区域均明显增加, 但扩张方向不同[4]。国外研究表明, 外来入侵植物普通豚草()对气候变暖反应同样敏感, 且RCP8.5情景下普通豚草在美国东部地区的适生范围远大于RCP4.5情景下[5]。在英国, HadCM3 (A1B)气候变暖情景下, 普通豚草在英国的适生区域同样明显增加, 表现为向东部和北部扩张[6]。
气候变暖条件下, 不同区域的不同外来入侵植物扩张的驱动因子不同。研究表明, 外来入侵植物刺萼龙葵、腺龙葵、北美刺龙葵和银毛龙葵向高纬度、高海拔区域扩散, 纬度和海拔是其主要的驱动因子[4]。互花米草在RCP4.5和RCP8.5的情景下, 扩张方向均为高纬度和低海拔区域, 驱动因子同为纬度和海拔[3]。普通豚草在英国的南部的驱动因子为水分, 英国北部的驱动因子为日照时长[6]; 而在美国东部, 普通豚草的扩张受一月份最高气温影响最大[5]。
目前, 应用较广的物种预测模型包括BIOCLIM[7]、BIOMAPPER[8]、BRT[9]、CLIMEX[10]、DOMAIN[11]、GAM[12]、GRAP[13]、和MAXENT[14]。王运生等利用ROC曲线的AUC值检验各预测模型预测准确率结果表明, MAXENT模型的准确率最高, AUC值达到0.95[15]。MAXENT模型原理为默认入侵物种在扩散过程中生态位是保守的, 将物种已知分布点的气候环境变量特征提取出来, 并将这些特征应用到未来气候环境特征中, 从而预测气候变暖趋势下外来入侵物种的潜在分布区域。Brown等开发的SDMs-toolbox工具正是融合了MAXENT模型的运算方法, 利用了地理信息系统的可视化和栅格数据的空间分析功能, 提取主要环境变量进行预测, 并对各环境变量的解释率进行计算和描述, 简化了物种潜在分布区域预测步骤[16]。
本研究选取辽宁省为“气候变暖条件下三裂叶豚草潜在分布区域预测”研究区域, 其原因为辽宁省是三裂叶豚草最早入侵我国的省份, 入侵时长近90年, 已在省内定殖、建群和扩散, 为研究三裂叶豚草潜在入侵分布区域提供充足的样本量。本研究基于SDMs-toolbox工具的最大熵算法, 结合辽宁省内三裂叶豚草分布点位、World Clim-Global数据库提供的气象预测数据和地形地势数据, 模拟分析气候变暖条件下三裂叶豚草在辽宁省潜在适生区域。其结果有助于理解水热条件对三裂叶豚草侵入、分布与扩散的影响, 为其他省份研究气候变暖条件下三裂叶豚草的潜在适生区域与危害范围提供参考。另外, 预测外来入侵植物的潜在分布, 阻止其进入适合生存的地区, 是预防和控制外来入侵植物扩张的重要举措。同时, 模拟气候变暖对外来入侵种潜在分布区域的影响有助于外来入侵物种的风险预警, 为外来入侵植物的防治提供技术支撑[17-20]。
1 材料与方法
1.1 研究区域
辽宁省位于我国东北地区的南部(38.43°43′ 3.181″—43°26′18.984″N, 118°53′22.765″—125°46′ 11.759″E, 图 1), 地处欧亚大陆东岸, 南濒黄海和渤海。辽宁省境内气候特征为温带大陆性季风气候, 雨热同期, 四季分明, 月平均温度和降雨量分别为6.4 ℃和642 mm。辽宁省两侧(辽东辽西)地形地貌为山地和丘陵, 平均海拔为800 m和500 m; 中部地区为平原地貌, 平均海拔为200 m。辽宁省境内河网密布交错, 大小河流共计300余条, 主要河流包括辽河、浑河、大凌河、太子河、饶阳河和鸭绿江等。
1.2 研究方法
1.2.1 模型介绍
本研究选取的MaxEnt模型是一种基于最大熵原理来选择或构建最佳分类器的模型。通过已有的物种分布数据和物种所在区域的环境数据, 对指定区域内该物种的生境适宜度进行评估[21-22]。SDM toolbox 2.0是基于一系列python语言脚本开发的物种分布模型工具, 它集成了ArcGIS软件特有的数据预处理、环境变量空间自相关分析和空间数据主成分分析等功能, 可以剔除高度自相关的环境变量且简化操作流程, 提高MaxEnt模型的应用效率[16]。
1.2.2 数据来源与处理
(1)物种分布数据(Occurrence data)
物种分布点位数据来源于本团队2010年-2019年外来生物监测数据, 包含143个具有准确经纬度的三裂叶豚草()分布现状点位, 利用SDM工具箱中的Spatially Rarefy Occurrence Data剔除掉空间自相关的点位, 最终保留84个三裂叶豚草点位数据。

图1 研究区域
Figure 1 Location of study areas
(2)气象数据(Climate data)
气象数据来源于World Clim - Global数据库(www. worldclim.org)(version 1.6和version 2.1)提供的2000年和2050年两个时期的气象因子, 本研究选取年平均气温(bio-01)、年平均气温日较差(bio-02)、等温性(bio-03)、气温季节性变动系数(bio-04)、最热月份的最高温度(bio_05)、最冷月份的最低温度(bio-06)、年温年较差(bio-07)、最湿季度平均温度(bio-08)、最干季度平均温度(bio-09)、最热季度的平均温度(bio-10)、最冷地区的平均温度(bio-11)、年降水量(bio-12)、最湿月份的降雨量(bio-13)、最干月份的降雨量(bio-14)、降水量季节性变化(bio-15)、最湿季度的降雨量(bio-16)、最干季度的降雨量(bio-17)、最温暖季度的降雨量(bio-18)、最冷季度的降雨量(bio-19)共19个气象因子作为气象因子候选指标, 分辨率为30 s。其中, 2050年的气象数据包括温室气体高浓度(Representative Concentration Pathways, RCPs 8.5)和低浓度(Representative Concentration Pathways, RCPs 4.5)两种排放情景。
(3)地形数据(Terrain data)
地形数据指标包括数字高程数据(Digital Elevation Model, DEM)、坡度和坡向3个变量。数字高程数据来源于中国科学研究院地理空间数据云(http://www.gscloud.cn/), 分辨率为30 m。以DEM数据为基础, 利用Arcgis的表面分析工具生成坡度和坡向数据, 分辨率均为30 m。为了保证气象数据与地形数据的分辨率一致, 利用SDMs-toolbox中的重采样工具将地形数据分辨率处理至1 km。
1.2.3 建立模型
(1)环境变量筛选
数据准备就绪后, 开始进行模型的的建立。利用SDMs-toolbox工具箱中的Remove Highly Correlated Variables工具对各环境因子进行空间自相关筛选, 若2个变量间相关性的绝对值大于0. 8, 那么只有1个变量选入模型。
通过对22个环境变量进行Person相关得出各指标间的相关关系, 最终保留相关系数绝对值小于0.8的10个环境变量。分别为bio-1年平均气温(℃)、bio-3等温性、bio-5最热月份的最高温度(℃)、bio-7年温较差(℃)、bio-12年降雨量(mm)、bio-14最干月份的降雨量(mm)、bio-15降水量季节性变化(mm)、DEM、坡度和坡向。
(2)数据运行参数
模型中将筛选后保留的三裂叶豚草生长点的75%用于与环境变量建模, 其余25%用于验证。设置模型运行次数为10次, 取10次模型模拟结果平均值作为最终的模拟结果。选择对每个环境变量创建响应曲线, 评价每个环境变量对模型的贡献率, 采用Jackknife刀切法检验对环境因子的重要性进行分析, 并采用受试者工作特征曲线(Receiver Operating Characteristic Curve, ROC)下面积即AUC(area under curve)对模型的精度进行评价。
(3)遗漏率计算
模型随机选取10%的三裂叶豚草生长点位进行遗漏率的评估, 获取各模型下的遗漏率(Omission Rates, OR), 遗漏率越低, 表示模型模拟的越好。

表1 环境变量Person相关分析结果
注: bio-1年平均气温; bio-3等温性; bio-5最热月份的最高温度; bio_7年温较差; bio-12年降雨量; bio-14最干月份的降雨量; bio-15降水量季节性变化。
(4)AUC值计算
ROC曲线是以灵敏度为纵坐标、以特异度为横坐标来表现一系列不同的分类阈值的曲线。本研究中的灵敏度为能正确预测适宜三裂叶豚草分布点位个数与三裂叶豚草实际存在的所有点位个数的比值; 特异度为能正确预测非适宜三裂叶豚草分布的点位个数据与实际所有非适宜三裂叶豚草分布的点位个数的比值。
AUC值为ROC曲线所覆盖的区域面积, AUC值越大, 分类器分类效果越好。当0.9<AUC≤1, 表明预测结果为“好”; 当0.8<AUC≤0.9, 表明预测结果为“较好”; 当0.7<AUC≤0.8, 表明预测结果为“一般”; 当0.6<AUC≤0.7, 表明预测结果为“较差”; 当0.5<AUC≤0.6, 表明预测结果为“失败”[27]。
(5)模型验证
三裂叶豚草的生长点位与环境变量之间符合哪一种预测模型, 主要由OR值、AUC值和模型的复杂性来决定。最理想的模型应依次满足“低OR值”、“高AUC值”和特征类参数最简三个特性(1线性; 2线性/二次; 3铰链; 4线性, 二次, 铰链; 5线性, 二次, 铰链, 乘积, 阈值)[27]。
由表2可知, 在各类预测模型的AUC和OR检验结果中, 第五种组合(Line-quadratic-hinge- threshold-product)的Train-AUC值最高, OR值最低, 因此确定预测模型为阈值特征预测模型。由图2可知, 在阈值模型预测下, 三裂叶豚草分布预测模型的Training-AUC值为0.902, 模型预测结果表现为“好”。
2 结果与分析
2.1 环境变量对三裂叶豚草分布影响
刀切法检验各环境变量对模型的贡献率的结果表明(表3), 高程对模型的贡献率最高, 为47.4%, 其次为坡度13.3%, 等温性对模型的贡献率为11.4%, 降水量季节性变化为9.9%。以上四个指标对模型的累计贡献率为82%, 说明以上四个指标基本解释了三裂叶豚草生境适宜性。坡向、年平均气温、年温较差、最热月份的最高温度、降雨量和最干月份的降雨量对三裂叶豚草生境适宜性影响较小。各环境变量对模型的重要性与贡献率的排序有所不同, 但高程对模型的重要性仍然排在第一位, 重要性值为58%。
通过贡献率最高的四个环境变量的响应曲线可以看出(图3), 高程处于0—200 m的范围内、坡度处于0度左右的区域三裂叶豚草适宜性指数较高, 并且呈现递减趋势。该结果说明三裂叶豚草适合分布在高程较低、坡度较缓的平原区域。该结果与实地调查结果相符, 三裂叶豚草在辽宁省的分布现状表现为两条主线路, 其一为以沈阳市为中心向西南和东北方向扩散; 其二以丹东市为源头, 沿海岸线向东南方向扩散。以上两条扩散线路所在区域高程值均处于0—200 m范围, 且第一条路线所经区域为典型河流冲积平原地貌, 降雨量的季节性变化相对较低(图4), 扩散速度相对较快。

表2 预测模型AUC和OR检验结果

图2 三裂叶豚草分布预测模型AUC和点位遗漏率检验结果
Figure 2 ROC curves and OR value of the distribution prediction model for
2.2 三裂叶豚草分布的预测结果
预测结果表明, 三裂叶豚草生境适宜性指数值域范围为0—1。将预测结果导入Arcgis10.6.1, 利用重分类方法将0.131作为适生等级的下限(训练遗漏率、预测面积和阈值平衡的阈值), 适宜度指数小于该值被认为三裂叶豚草暂无分布; 从该值至0.290(敏感度和特异性的和最大的阈值)作为低度适生区; 从该值至0.402(敏感度和特异性相等的阈值)作为中度适生区; 大于0.402作为高度适生区。
由表4可知, 预测结果表明三裂叶豚草2050年在辽宁省的分布范围明显增加。在高温室气体排放量拟合气候变化的情景下, 三裂叶豚草的无分布地区和低度适生区面积明显减少, 特别是低度适生区, 分别减少了602.82 km2和6329.04 km2, 面积占比分别减少了0.24%和2.5%; 中度和高度适生区大幅度面积增加, 分别增加了2554.81 km2和4377.11km2, 面积占比增加了1.01%和1.73%。在低温室气体排放量拟合气候变化的情景下, 三裂叶豚草适宜扩散的面积较高温室气体排放量拟合气候变化的情景下大。其中无分布区域和低度适生区域分别减小了3886.7 km2和7033.2 km2, 面积占比降低了1.53%和2.78%; 中度和高度适生区分别增加了3137.32 km2和7782.58 km2, 面积占比增加了1.24%和3.07%。

表3 各环境变量对三裂叶豚草分布贡献率
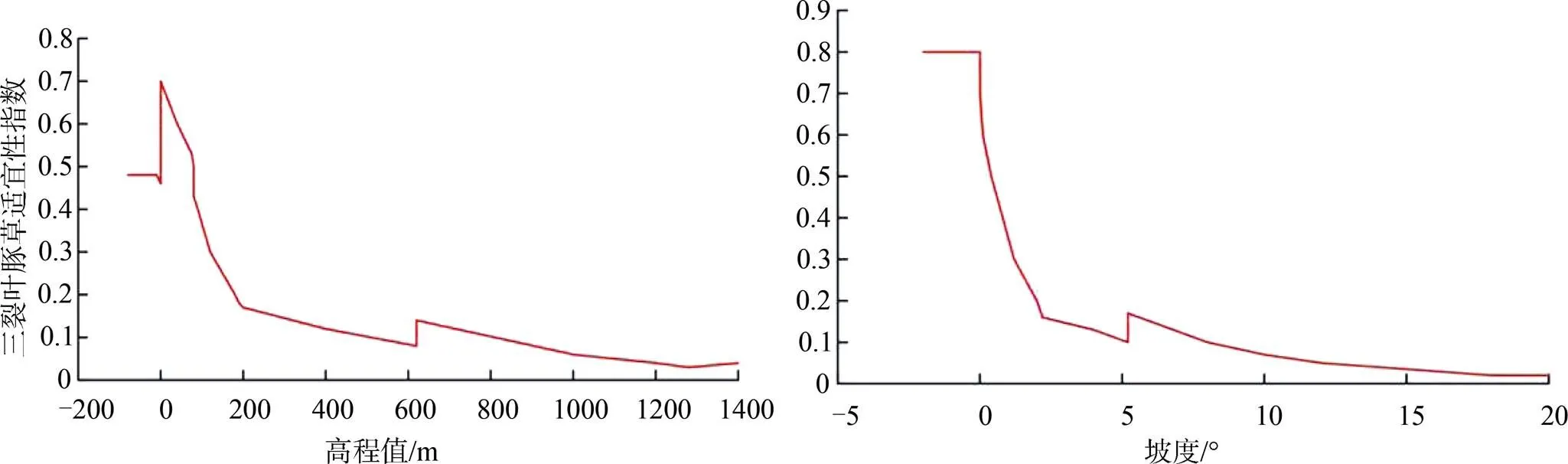
图3 三裂叶豚草适宜性对高程与坡度响应回归曲线
Figure 3 Suitability regression curve ofto elevation and slope
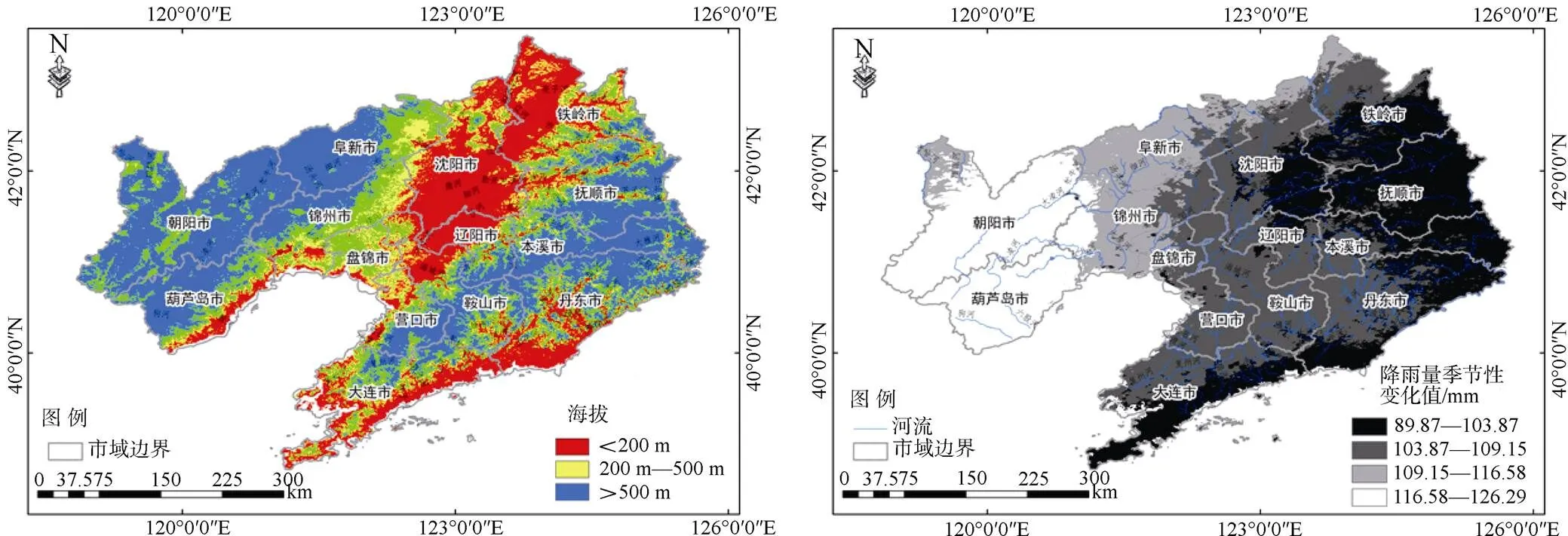
图4 DEM和降雨量季节性变化分布图
Figure 4 Distribution of DEM and seasonal variation in rainfall

表4 2000年-2050年三裂叶豚草生境适宜度面积变化
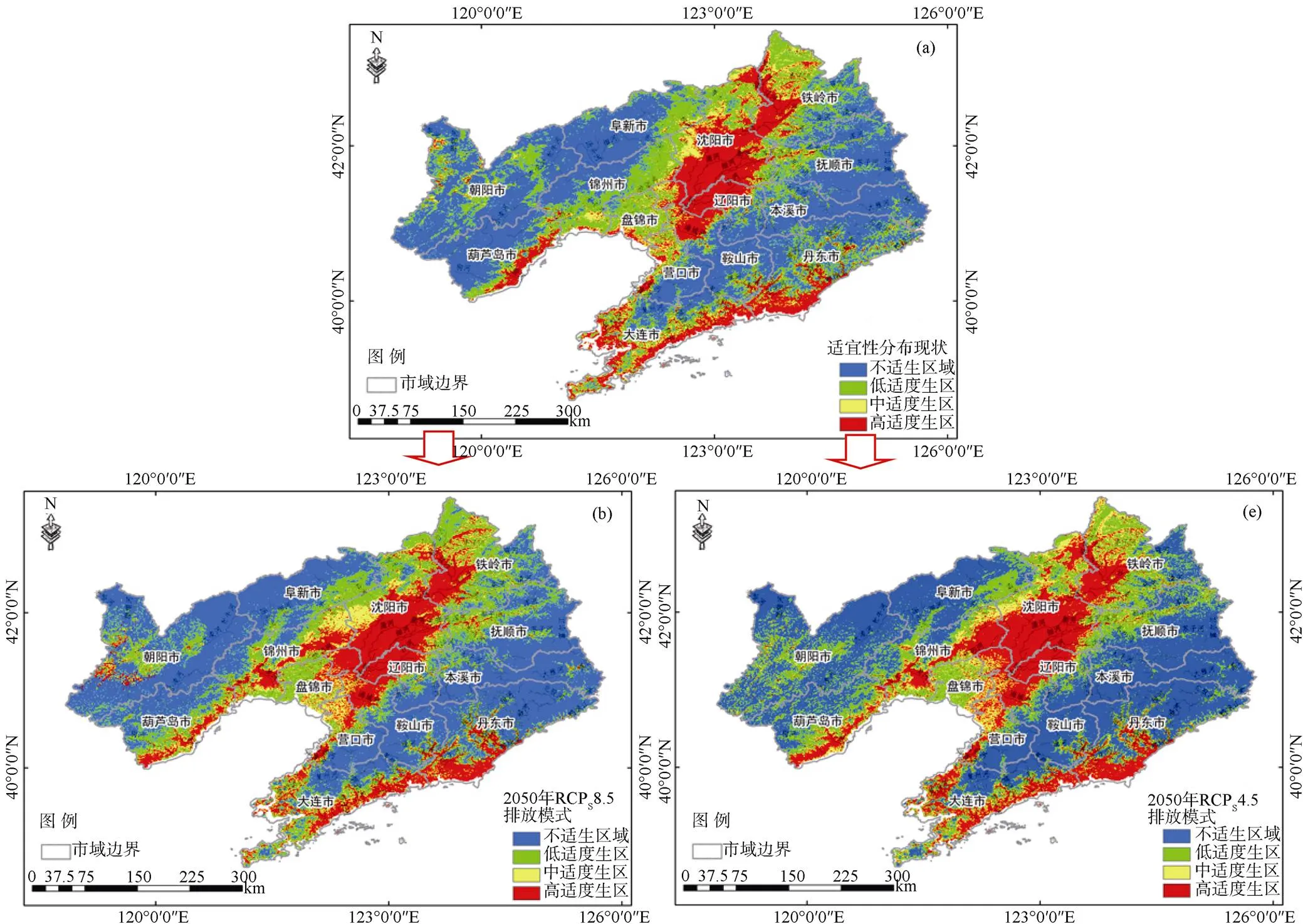
图5 2050年三裂叶豚草潜在分布预测结果
Figure 5 Potential distribution areas ofin 2050
3 讨论
3.1 三裂叶豚草潜在分布区域预测
本研究基于SDMs-toolbox工具MaxEnt模型, 预测了温室气体不同排放量影响的气候条件下三裂叶豚草在辽宁省潜在分布区域。三裂叶豚草在辽宁省的分布现状表明, 三裂叶豚草主要分布在辽宁中部地区河流沿岸、丹东和大连沿海沿岸, 辽西地区锦州和葫芦岛沿海地区分布明显, 其他区域零星分布, 与辽宁省三裂叶豚草调查点位基本相符[23-24]。三裂叶豚草预测结果表明, 无论是在温室气体高或低浓度排放量的情景下, 2050年三裂叶豚草在辽宁省内适生区范围不断扩散, 面积增加。温室气体低浓度排放量的情景下(Representative Concentration Pathways, RCP 4.5), 中度和高度适生区分别增加了3137.32 km2和7782.58 km2, 面积占比增加了1.24%和3.07%。温室气体高浓度排放量的条件下, 三裂叶豚草的中度和高度适生区面积大幅度增加, 分别增加了2554.81 km2和4377.11 km2, 面积占比增加了1.01%和1.73%。在美国东部地区, 已有气候变暖促进豚草属植物普通豚草的适生区面积增加的类似结论, 同时证明2050年RCP4.5和RCP8.5两种情景下普通豚草的适生区面积均增加[32], 与本研究结果区别在于Case等研究结果中RCP8.5的情境下普通豚草的扩散面积高于RCP4.5, 其原因可能为普通豚草为喜湿怕旱植物, 对降水的敏感性更强, 而降雨量与温室气体排放量呈正相关[5]。
入侵物种潜在分布范围预测结果可用于土地管理规划中[25-26]。本研究结果表明, 截止于2050年, 辽宁省内三裂叶豚草扩散主要表现为两个方向, 其一为以中部平原地区为中心向西北方向的锦州、阜新沿大凌河与小凌河方向扩张, 由原来的低度适生区转变为高度适生区, 验证了三裂叶豚草喜湿的生物学特征[35]; 其二为沿辽宁中部河流向西南方向扩张, 营口和大连沿海地区扩散严重。除辽宁省东北部铁岭、抚顺和本溪三个以山地为主的城市, 其他各市适生区均有明显增加。温室气体高浓度排放量和低浓度排放量情景下, 扩散方向大致相同, 但温室气体高浓度排放量情景下西北方向扩散面积明显小于低浓度排放量的情景, 其原因可能为高浓度排放量模拟条件下温度高于低浓度排放量模拟的情况, 而三裂叶豚草为喜湿、喜冷植物, 因此温度的快速增加并没有促进三裂叶豚草的扩散。
3.2 影响生境适宜性主要环境变量
Jackknife检验环境变量对生境适宜性预测的重要性分析结果表明, 高程对模型的贡献率最高, 为47.4%, 其次为坡度13.3%, 等温性对模型的贡献率为11.4%, 降水量季节性变化为9.9%。其中高程的贡献率起到主要作用, 并且高程和坡度单因素变量影响下的ROC 曲线结果表明, 高程处于0-200m的范围内、坡度处于0度左右的区域三裂叶豚草适宜性指数较高。三裂叶豚草适合分布在高程较低、坡度较缓的平原区域。分析其原因可能为辽宁省的中部平原地区为河流冲积平原, 河网密布, 外加北方多发多年一遇洪水, 河流两岸植被以草本植被群落和农田为主, 群落环境开放, 适合喜阳喜湿的三裂叶豚草生存该结论与王苏铭等和邵云玲等研究结果一致, 指出普通豚草在东三省内三江平原、松嫩平原、辽河平原和辽东半岛分布较多, 在大兴安岭、小兴安岭、长白山等高海拔区域分布较少[27-28]。柳晓燕等对我国普通豚草适生区预测结果显示, 最冷季度平均湿度和最湿季度湿度是影响普通豚草分布的主要环境变量[29], 与本研究得出的DEM为主要影响三裂叶豚草适生区面积变化的主要环境变量的结果不同, 其原因可能为研究的尺度不同, 与辽宁省尺度相比, 全国尺度上影响三裂叶豚草潜在适生区域的因素比较复。另外, 本研究中Jackknife检验结果显示年均降雨量与年平均气温对三裂叶豚草的生境适宜性影响较小, 其原因可能为辽宁省范围相对较小且纬度跨度小, 因此降雨量和平均气温差异较小, 故以上两个气象因子贡献率不高的原因。
4 结论
研究结果表明, 2050年高、低温室气体浓度排放量影响的气候条件下三裂叶豚草在辽宁省的分布范围均有所扩散, 扩散方向表现为以辽宁中部区域为中心, 沿大小凌河向西北方向扩散和沿辽河向西南方向扩散。因此, 对于辽宁省内河流生态系统的三裂叶豚草防治工作需要引起相关部门的重视, 做到早发现、早清除, 防止三裂叶豚草生长形成优势群落。
[1] 姜传明, 曲秀春, 刘祥君. 三裂叶豚草的分布、危害和传播特点[J]. 牡丹江师范学院学报(自然科学版), 1999, 2(1): 26–27.
[2] LI Xianping, LIU Xuan, KRAUS F, et al. Risk of biological invasions is concentrated in biodiversity hotspots[J]. Frontiers in Ecology and the Environment, 2016, 14(8): 411–417.
[3] 刘金雪. 气候变化对外来入侵植物互花米草潜在分布区的影响[D]. 南京: 南京师范大学, 2016.
[4] 唐瑶. 气候变化条件下四种入侵植物在我国潜在分布预测分析[D]. 安徽: 安徽农业大学, 2018.
[5] CASE M J, STINSON K A. Climate change impacts on the distribution of the allergenic plant, common ragweed () in the eastern United States[J]. PLoS ONE, 2018, 13(10): e0205677.
[6] Storkey J, Stratonovitch P, Chapman D S, et al. A Process-Based Approach to predicting the effect of climate change on the distribution of an invasive allergenic plant in Europe[J]. PLoS ONE, 2014, 9(2): e88156.
[7] Csuti B, Margules C R, Austin M P. Nature Conservation: cost effective biological surveys and data analysis[J]. Journal of Wildlife Management, 1991, 56(3):621.
[8] HIRZEL A H, HAUSSER J, CHESSEL D, et al. Ecological-niche factor analysis:how to compute habitat- suitability maps without absence data[J]. Ecology, 2002, 83(7): 2027–2036.
[9] FRIEDMAN J H, HASTIE T, TIBSHIRANI R. Additive logistic regression: a statistical view of boosting[J]. Annals of Statistics, 2000, 28(2): 337–407.
[10] SUTHERST R W, MAYWALD G F, SKARRATT D B. Predicting insect distributions in a changed climate. In: Insects in Changing Environment (eds Harrington R, Stork NE)[M]. London: Academic Press, 1995: 59–91.
[11] CARPENTER G, GILLISON A N. WINTER J. DOMAIN: a flexible modelling procedure for mapping potential distributions of plants and animals[J]. Biodiversity and Conservation, 1993, 2(6): 667–680.
[12] YEE T W, MITCHELL N D. Generalized additive models in plant ecology[J]. Journal of Vegetation Science, 1991, 2(5): 587–602.
[13] STOCKWELL D, PETERS D. The GARP modelling system: problems and solutions to automated spatial prediction[J]. International Journal of Geographical Information Systems, 1999, 13(2): 143–158.
[14] PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions[J]. Ecological Modelling, 2006, 190(3/4): 231–259.
[15] 王运生, 谢丙炎, 万方浩, 等. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4): 365–372.
[16] Brown J L. SDM toolbox: a python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses[J]. Methods in Ecology and Evolution, 2014, 5(7): 1–110.
[17] BYEON D H, JUNG S H, LEE W H. Review of CLIMEX and MaxEnt for studying species distribution in South Korea[J]. Journal of Asia-Pacific Biodiversity, 2018, 11(3): 325–333.
[18] CASE M J, STINSON K A. Climate change impacts on the distribution of the allergenic plant, common ragweed () in the eastern United States[J]. PLoS ONE, 2018, 13(10): e0205677.
[19] CHAPMAN D S, SCALONE R, STEFANIC E, et al. Mechanistic species distribution modeling reveals a niche shift during invasion[J]. Ecology, 2017, 98(6): 1671-1680.
[20] SKALOVA H, GUO Wenyong, WILD J, et al.in the Czech Republic: History of invasion, current distribution and prediction of future spread[J]. Preslia, 2017, 89(1): 1–16.
[21] LEI J C, XU H G. MaxEnt-based prediction of potential distribution ofin China[J]. Journal of Ecology and Rural Environment, 2010, 26 (2): 137–141.
[22] 麻亚鸿. 基于最大熵模型和地理信息系统预测藓类植物的地理分布范围——以广西花坪自然保护区为列[D]. 上海:上海师范大学, 2013.
[23] 刘旭昕, 方芳. 阜新外来入侵有害生物——杂草调查及防控建议[J].内蒙古林业调查设计, 2011, 34(1): 68–69+62.
[24] 曲波, 吕国忠, 杨红, 等. 辽宁省外来入侵有害生物—— 杂草调查[J]. 沈阳农业大学学报, 2006, 37(4): 587–592.
[25] SCHWARTZ M W. Using niche models with climate projections to inform conservation management decisions[J]. Biological Conservation, 2012, 155(4): 149–56
[26] IVERSON L, MCKENZIE D. Tree-species range shifts in a changing climate: detecting, modeling, assisting[J]. Landscape and Ecological Engineering, 2013, 28(5): 879–89
[27] 王苏铭, 张楠, 于琳倩, 等. 北京地区外来入侵植物分布特征及其影响因素[J]. 生态学报, 2012, 32(15): 4618–4629.
[28] 邵云玲, 曹伟. 外来入侵植物豚草在中国东北潜在分布区预测[J].干旱区资源与环境, 2017, 31(7): 172–176.
[29] 柳晓燕, 李俊生, 赵彩云, 等. 基于MAXENT模型和ArcGIS预测豚草在中国的潜在适生区[J].植物保护学报, 2016, 43(6): 1041–1048.
Prediction of potential invasive region forbased on SDMs-toolbox with climate warming in Liaoning Province
WANG Di1,2, QU Bo3,*, ZHOU Bin4, ZHANG Yiran1, LIU Zhi1
1. Shenyang Academy of Environmental Science,Liaoning Provincial Key Laboratory for Urban Ecology,Shenyang110161, China 2. Research Center for Eco-Environmental Sciences, Chinese Academy of Science, Beijing 100085, China 3. Biological science and Technology College of Shenyang Agricultural University,Liaoning Key Laboratory for Biological invasions and Global Changes,Shenyang110866,China 4. Liaoning Ecological Meteorological and Satellite Remote Sensing Center, Shenyang 110866, China
In order to clarify the effects of climate warming on the potential adaptive regionsand reveal the expansion direction of, Liaoning Province was selected as the sample region to simulate the potential distribution areas ofin 2050 under the condition of climate warming. Results showed that under the lower concentration emissions of greenhouse gas (RCP4.5), the area of 10919.9 km2increasecompared with 2019; the moderate and high adaptive areas were increased by 3137.32 km2(1.24%) and 7782.58 km2(3.07%), respectively. Under the higher concentration emissions of greenhouse gas (RCP8.5), the area was increased by 6932.2 km2compared with 2019; the moderate and high adaptive areas were increased by 2554.81 km2(1.01%) and 4377.11 km2(1.73%). The result showed that the higher concentration emissions of greenhouse gas did not accelerate the faster expansion of. Whatever the lower or higher concentration the greenhouse gas was, the spreading direction presented consistently, both southwest and northwest. The one of the spread direction was centered on central plain regions and spread to Jinzhou City and Fuxin City from northwestern, expanding along the Daling River and Xiaoling River; regional suitability changed from low-level to high-level. The other direction was along the Liaohe river to Yingkou City and Panjin City. The habitat suitability was most affected by elevation from the Jackknife test results, occupying 47.4%. ROC results showed suitability index ofwas higher in the area of 0-200 m above sea level with low gradient, which illustrated it was suitable for growing in plain regions. The results provide some supports for risk warning and prevention from invasive species ofin Liaoning Province and even China.
invasive species; prediction; SDMs-toolbox; MaxEnt model; risk assessment
10.14108/j.cnki.1008-8873.2022.02.008
Q14
A
1008-8873(2022)02-066-09
2020-05-24;
2020-06-22
国家重点研发计划(2017YFC1200100); 辽宁省科技厅农业攻关及产业化指导计划(2019JH8/10200017); 国家自然科学基金(31901209); 中国 博士后科学基金(2018M641492); 辽宁省自然科学基金(20180551024); 沈阳市科学事业费科技项目(sysy2018-041)
王迪(1990—), 女, 辽宁沈阳人, 博士, 主要从事生态环境修复与生物多样性研究, E-mail: ecology_wd@163.com
通信作者:曲波, 女, 博士后, 教授, 主要从事生物多样性与生物入侵机理研究, E-mail: syau_qb@163.com
王迪, 曲波, 周斌, 等. 基于SDMs-toolbox预测气候变暖趋势下辽宁省三裂叶豚草潜在入侵区域[J]. 生态科学, 2022, 41(2): 66–74.
WANG Di, QU Bo, ZHOU Bin, et al. Prediction of potential invasive region forbased on SDMs-toolbox with climate warming: the case study of Liaoning province[J]. Ecological Science, 2022, 41(2): 66–74.
猜你喜欢
西部林业科学(2022年6期)2023-01-03
中国野生植物资源(2022年3期)2022-04-19
治淮(2022年4期)2022-01-01
海河水利(2021年4期)2021-08-30
数学小灵通·3-4年级(2021年6期)2021-07-16
电脑爱好者(2020年10期)2020-07-28
植物资源与环境学报(2020年2期)2020-04-20
电脑爱好者(2019年16期)2019-10-30
竹子学报(2018年2期)2018-10-25
青岛科技大学学报(社会科学版)(2015年4期)2016-01-25
