
丹霞地貌石松类和蕨类植物区系比较研究
2022-04-15 06:54梁胜王梦楠韦宝婧胡希军陈存友
生态科学 2022年2期
梁胜, 王梦楠, 韦宝婧, 胡希军, 陈存友
丹霞地貌石松类和蕨类植物区系比较研究
梁胜, 王梦楠, 韦宝婧, 胡希军*, 陈存友
中南林业科技大学风景园林学院, 长沙 410004
在野外调查及资料收集的基础上, 对福建、广东两省中4个纬度相近区域的石松类和蕨类植物区系基本组成、地理成分、区系相似性等进行分析, 比较丹霞地貌与非丹霞地貌石松类和蕨类植物区系特征差异。结果显示:(1)研究区有石松类和蕨类植物38科80属206种, 其中丹霞山35科65属134种, 冠豸山31科51属103种, 观音岽31科57属103种, 南雄丹霞梧桐31科57属96种, 优势科明显但属种分化程度不高。(2)在区系丰富性上, 非丹霞地貌区丰富性较高于丹霞地貌区, 具体表现为丹霞山(0.74)>观音岽(无丹霞地貌)(0.14)>南雄丹霞梧桐(0.07)>冠豸山(0.02)。(3)在区系地理成分上, 科属水平均以热带成分为主(占84%以上), 与热带亲缘程度较高; 在种水平上, 热带成分占比均为59%以上, 呈热带向温带过渡特征。(4)在地理亲缘关系上, 南雄丹霞梧桐(丹霞地貌)与观音岽(非丹霞)亲缘关系明显, 科、属、种水平上相似性分别为0.88、0.97、0.8, 两者同质性明显。该结果为探讨丹霞地貌地带石松类和蕨类植物植物的起源和发育提供重要依据, 对丹霞地貌的植物资源合理开发利用具有参考意义。
丹霞地貌; 石松类和蕨类植物; 区系组成; 相关性分析
0 前言
丹霞地貌是指第三纪红色岩系发育的一种类似岩溶地貌的地质形态, 国内主要分布于广东北部和江西、福建、湖南等地区[1]。自20世纪30年代陈国达[2]提出“丹霞地貌”的概念以来, 丹霞地貌作为地貌学一个新的研究领域被不断扩展[3-4]。丹霞地貌有别于石灰岩地貌和花岗岩地貌的特殊地貌类型, 孕育出较为独特的植被类型和植物区系。因其独特地貌形态产生特殊的生态分异现象, 出现了一系列关于丹霞地貌特殊生境的研究报道[5-7], 随着当今生物多样性研究的不断深入, 特殊生境植物区系研究也逐渐成为当今植物学领域的热点[8]。
丹霞地貌中沟谷和小盆地往往被山块围合, 温度较高, 水分充足, 易形成湿热的小环境, “干”“湿”岛效应明显[9]。蕨类植物对自然条件具有高度的敏感性, 因环境因子的异质性, 特殊生境下植物区系的组成以及区系植物的适应性特征必然有别于普通生境[10]。石松类和蕨类植物是森林草本层的重要组成部分, 石松类还是最早出现的维管植物, 处于系统演化的过渡阶段, 是植物进化过程中的重要环节。目前中国现有蕨类植物63科221属2452种, 占世界总种数的1/4左右[11], 是东亚蕨类植物区系的主体部分, 因此石松类和蕨类植物在植物类群和区系构成中有着不可或缺的地位[12]。目前国内关于石松类和蕨类植物区系的研究还远远不如种子植物研究广泛和深入[13-14], 能收集到的关于蕨类植物区系研究报道有限[15], 对于特殊生境下石松类和蕨类植物区系的研究有待拓展。
当前对中国丹霞地貌植被的研究多以局域性的植物群落物种多样性、群落结构和物种空间分布为主[16-17], 对种子植物的分析研究较为全面, 缺少对石松类和蕨类植物的调查研究[18], 仅有的丹霞地貌蕨类研究均以蕨类植物资源调查和蕨类多样性分析为主[19]。调查研究丹霞地貌石松类和蕨类资源, 可进一步完善构建丹霞地貌区植物数据库, 是对全面探讨丹霞地貌植物区系地理特征的一个必要补充。为此, 研究选取纬度相近下3个极具代表性的丹霞地貌区域, 1个非丹霞地貌区作为对照组, 通过野外实地调查以及文献查阅, 结合科考报告整理出各个区域的植物数据库, 对石松类和蕨类植物区系特征进行比较, 分析区系地理成分特征差异, 以期丰富丹霞地貌地带区系方面的相关研究, 有助于了解丹霞地貌地带蕨类植物区系的起源及演化, 对合理开发利用丹霞地貌植物资源和保护整个生物多样性都具有十分重要的意义。
1 研究区概况
本研究选取4个纬度相近(北纬25°左右)的区域植物群落为研究对象, 见图1, 横跨福建广东两省, 均属于我国夏热冬暖地区, 有着良好的水热条件, 为石松类和蕨类植物的繁衍创造良好的生存环境。研究区基本概况如表1所示, 图表中1为广东韶关丹霞山国家级自然保护区(简称丹霞山, 有丹霞地貌), 2为南雄丹霞梧桐县级自然保护区(简称南雄丹霞梧桐, 有丹霞地貌), 3为南雄观音岽县级自然保护区(简称观音岽, 无丹霞地貌), 4为连城冠豸山国家森林公园(简称冠豸山, 有丹霞地貌)。
2 材料与方法
2.1 数据来源
随着分子系统学研究的发展, 蕨类植物的科属概念和分类系统近年发生了较大变化, 本研究通过野外调查, 采集标本调查南雄丹霞梧桐县级自然保护区、南雄观音岽县级自然保护区的石松类和蕨类植物, 同时通过收集其他另外三个研究区的科学考察报告, 参考前人的研究成果[20-21], 整理出研究区的植物名录, 并依照《中国植物志》、《Flora of China》[22-23]等进行鉴定分类, 消除异名产生的影响, 以秦仁昌1978分类系统为基础按照现代分子系统分类学对源数据中蕨类植物的类群归属进行整理, 得到各研究区最终石松类和蕨类植物名录。
2.2 统计分析
区系地理成分类型的划分参考吴征镒、陆树刚等[24-25]分布区类型的划分方法将中国石松类和蕨类植物划分为13种分布区类型3种亚型。同时计算研究区内热带区系成分与温带区系成分相应的比值R/T, R为各种热带分布类型(即2—7分布型)之和, T为各种温带分布类型(即8—13分布型)之和。

图1 研究区位置示意图
Figure 1 Location map of the study area
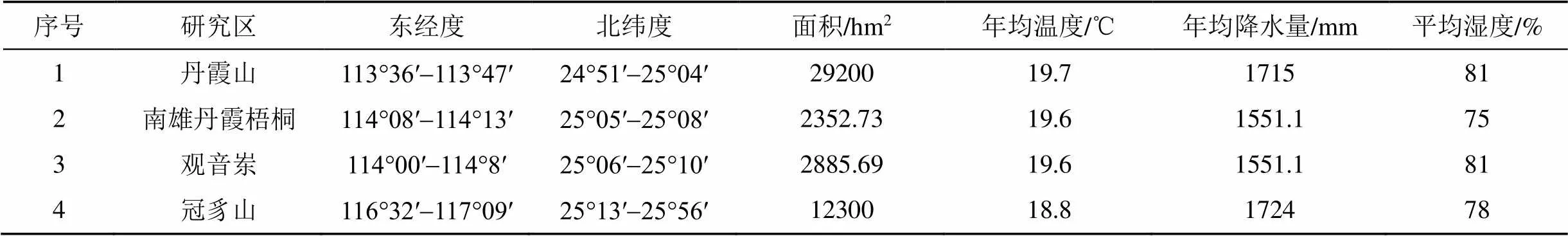
表1 研究区域基本特征
区系分化程度参照沈泽昊等[26]提出的方法利用种数与属数的比值以及种数与科数的比值分别代表属和科的区系分化强度, 对石松类和蕨类植物区系分化强度进行探讨。
区系相似性采用张镱锂[27]的方法计算对石松类和蕨类植物区系的相似程度进行计算, 区系相似性系数定义为:
Sc=2C/(A+B)×100%
式中Sc为相似性系数, C为两地除世界分布类型外共有属(或种)数, A和B分别为两地除世界分布类型外的总属(或种)数。
区系丰富性采用左家哺综合系数[28]对蕨类区系的地理成分丰富性进行分析, 定义为:

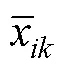
3 结果与分析
3.1 区系基本组成
研究区有石松类和蕨类植物38科80属206种(表2—表3), 分别占中国石松类和蕨类植物的科(40)、属(178)、种数(2147)的95.0%、44.9%、9.6%, 其中丹霞山共计134种, 隶属于35科65属; 冠豸山共计103种, 隶属于31科51属; 观音岽共计103种, 隶属于31科57属; 南雄丹霞梧桐共计96种, 隶属于31科57属。丹霞山优势科属明显, 物种丰富组成多样, 其中较大科有水龙骨科(Polypodiaceae, 16/8, 种数/属数, 下同)、凤尾蕨科(Vittariaceae, 15/3)、卷柏科(Selaginellsceae, 11/1)、铁角蕨科(Aspleniaceae, 10/2种)4科, 占自身总种数的46%; 较大属有凤尾蕨属(Pteris, 12种)、卷柏属(Selaginella, 11种)2属, 占自身总种数的12%。除丹霞山外, 其他研究区均无较大属出现。观音岽较大科有鳞毛蕨科(Dryopteris, 11/7)、水龙骨科(Polypodiaceae, 11/4)、凤尾蕨科(Pteris, 10/2)、金星蕨科(Pronephrium, 10/7)4科, 占自身总种数的41%。冠豸山较大科有鳞毛蕨科Dryopteris, 16/5)、水龙骨科(Polypodiaceae, 14/8)、蹄盖蕨科(Diplazium, 10/3)3科, 占自身总种数的39%。南雄丹霞梧桐仅水龙骨科(Polypodiaceae, 11/8)1科, 占自身总种数的11%。研究区除南雄丹霞梧桐外优势科明显, 除丹霞山外均缺乏较大属。
从历史成分看, 在研究区分布的石松类和蕨类植物中, 既有在系统位置上被认为是较原始的科, 如石杉科Huperziaceae、卷柏科Selaginellaceae、海金沙科Lygodiaceae、膜蕨科Hymenophyllaceae等类群, 也有被认为是较进化的科, 如水龙骨科Polypodiaceae、蘋科Marsileaceae、槐叶蘋科Salviniacae、满江红科Azollaceae等, 同时也有位于两者之间的蹄盖蕨科Anisocampium 、金星蕨科Thelypteridaceae、凤尾蕨科Pteridaceae、鳞始蕨科Lindsaeaceae、铁线蕨科Adiantaceae、蹄盖蕨科Athyriaceae等类群, 可见研究区石松类和蕨类植物在系统排序上分布着不同的发展阶段, 起源古老, 这表明研究区内石松类和蕨类植物在系统发育上或进化关系上存在较为连贯的谱系关系。
3.2 区系分化强度
通过对研究区石松类和蕨类植物科、属、种数量的分析(表4), 发现在科的水平上, 各研究区区系分化强度差异不明显, 其中丹霞山分化强度最强(3.83), 南雄丹霞梧桐最小(3.10); 在属的水平上, 区系分化强度大小关系为丹霞山(2.06)>冠豸山(2.02)>观音岽(1.81)>南雄丹霞梧桐(1.68)。整体而言, 研究区科的分化强度明显高于属, 科水平上表现出较强的区系分化特征, 属水平上分化特征不明显。表2、表3中研究区均分布有较大科、中等科、少种科、单种科, 但仅丹霞山存在较大属其他均只分布中等属、少种属、单种属, 也可以进一步说明研究区科属分化强度的差异。
3.3 区系丰富性
经计算(表5), 研究区石松类和蕨类区系综合丰富性指数的大小关系为:丹霞山(0.74)>观音岽(0.14)>南雄(0.07)>冠豸山(0.02), 其丰富性与经度变化关系不显著。在科丰富性指数上, 丹霞山(0.16)显著高于其他区域, 观音岽、南雄丹霞梧桐、冠豸山三者间无明显差异, 均在0.2—0.3; 在属丰富性指数上, 丹霞山最高(0.22), 观音岽、南雄丹霞梧桐次之(0.06), 最低为冠豸山(-0.05); 在种丰富性指数上, 丹霞山仍为最高(0.36), 观音岽次之(0.05), 南雄丹霞梧桐最低(-0.02)。

表2 石松类和蕨类植物区系科的组成
<
注:A/B, A. 该科属种的数目, B.占总科属种数的比例, 比例计算不包含世界分布。

表3 石松类和蕨类植物区系属的组成
注: A/B, A.该科属种的数目, B.占总科属种数的比例, 比例计算不包含世界分布。

表4 石松类和蕨类植物区系分化强度
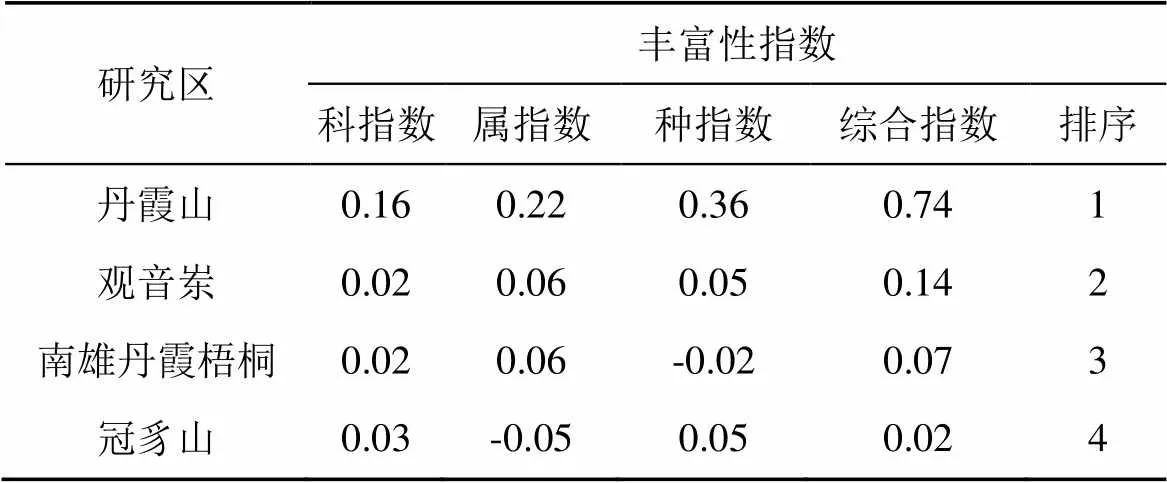
表5 石松类和蕨类植物区系丰富性
3.4 区系相似性
相似性分析表明(表6—表8):在科级水平上, 由于较近的地理关系, 区系成分易于交流, 各研究区相似性系数较大(均在0.8左右), 其中观音岽与冠豸山相似性最大(0.89); 在属级水平上, 南雄丹霞梧桐与观音岽相似性系数最大(0.97), 说明两地有着较强的历史联系, 除此之外各区域间的相似性系数无较大差异(均为0.7左右); 在种级水平上, 丹霞山与冠豸山的相似性最小(0.48), 南雄丹霞梧桐与观音岽的相似性系数最大(0.83), 明显高于其他研究区, 有着较强的独立性, 这显示出两者区系起源和演化关系密切, 这也与其近代地理位置关系较为吻合。
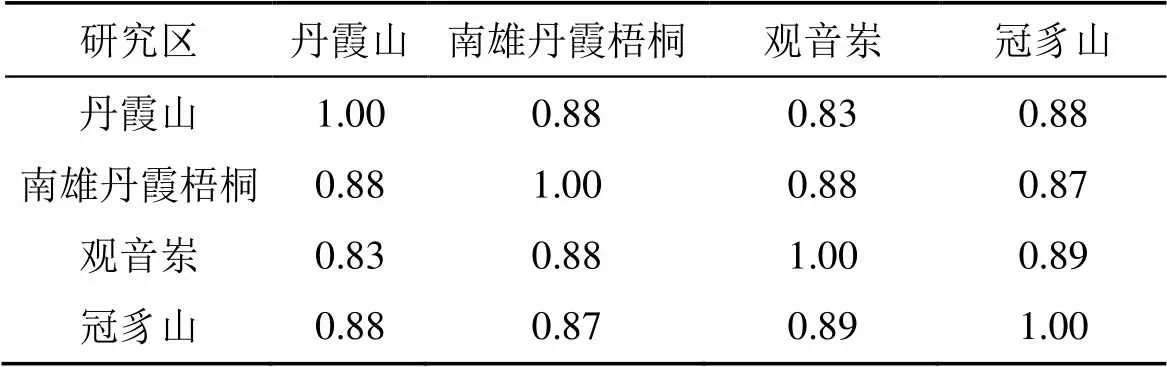
表6 科相似性系数

表7 属相似性系数

表8 种相似性系数
3.5 区系地理成分
按照研究区石松类和蕨类植物现代地理分布情况, 从表9可见, 在科水平上, 各研究区区系地理成分格局较为相似, 均以泛热带分布(占42%以上)、热带亚洲分布(占25%以上)为主, 均存在中国特有分布(占10%左右)。研究区蕨类植物的区系地理与热带分布类型的亲缘关系密切, 具有较强的热带属性, 热带区系成分占主体部分(占87%), 其余为温带成分(占13%), 这与其所处的地理位置、气候条件有很大关系。各研究区/值大小关系为:丹霞山(6.67)>南雄丹霞梧桐(5.67)> 观音岽(5.33)>冠豸山(5.67), 研究区有着很强的纬度地带性, 即热带性质明显, 这与他们起源于古生代湿热的、统一的大陆有关, 其中丹霞山热带属性最强; 在属水平上, 从表10可见, 区系成分以泛热带分布(占40%以上)、热带亚洲分布(占10%)以及旧大陆热带分布(占9%以上)为主, 与科相比整体分布格局相似但无中国特有分布, 区系特有性有所降低; 在种水平上, 从表11可见, 区系成分以热带亚洲分布(占43%以上)、中国特有分布(占35%以上)以及东亚分布(占27%以上)为主体, 东亚分布成分中主要右中国–日本分布构成, 各研究区/值大小关系为观音岽(1.86)>观音岽(1.85)> 丹霞山(1.65)>冠豸山(1.41)。与科属区系成分比较, 种区系热带成分明显降低, 种区系热带成分占59%以上, 温带成分占35%以上, 说明其区系成分呈热带向温带过渡特征, 且中国特有分布占比的提高也说明研究区特有性不断增强。
4 讨论
4.1 优势科属比较
在区系基本组成上, 研究区共分布有石松类和蕨类植物38科80属206种, 由于纬度地带性, 相似的大地理环境使得研究区科属组成较为相似, 且均出现有相似的优势科, 如水龙骨科Polypodiaceae、凤尾蕨科Pteris等。各研究区优势科明显(占总种数39%以上), 分化程度普遍高于属水平, 其中南雄丹霞梧桐最明显, 科分化强度为3.10, 但属分化强度仅为1.68。除丹霞山外, 各研究区属间以少种属占主要优势(占34%以上), 如凤尾蕨属Pteris、鳞毛蕨属Dryopteris、卷柏属Selaginella等, 这集中反映出生境的特殊很大程度上抑制了属种的分化。
4.2 区系丰富性比较
在区系丰富结果中, 观音岽(非丹霞地貌)面积虽不及冠豸山四分之一, 但其种丰富性指数(0.14)却高于有丹霞地貌的冠豸山(0.02), 与同面积水平的南雄丹霞梧桐(丹霞地貌)相比, 其种丰富性仍高于南雄丹霞梧桐(0.07)。蕨类生长在观音岽展现出较强的适宜性, 这表明丹霞地貌的特殊生境对石松类和蕨类植物的丰富度产生了较大影响, 出现了特异于非丹霞地貌区的区系组成特征, 这和丹霞地貌区独特的立地条件和所形成的孤岛效应有关[30], 使得丹霞地貌产生了一定程度的生境狭窄特有现象。

表9 石松类和蕨类植物区系科的分布型
<
注:A(B), A. 该分布区类型的数目, B. 占区系总数的比例, 比例计算不包含世界分布。
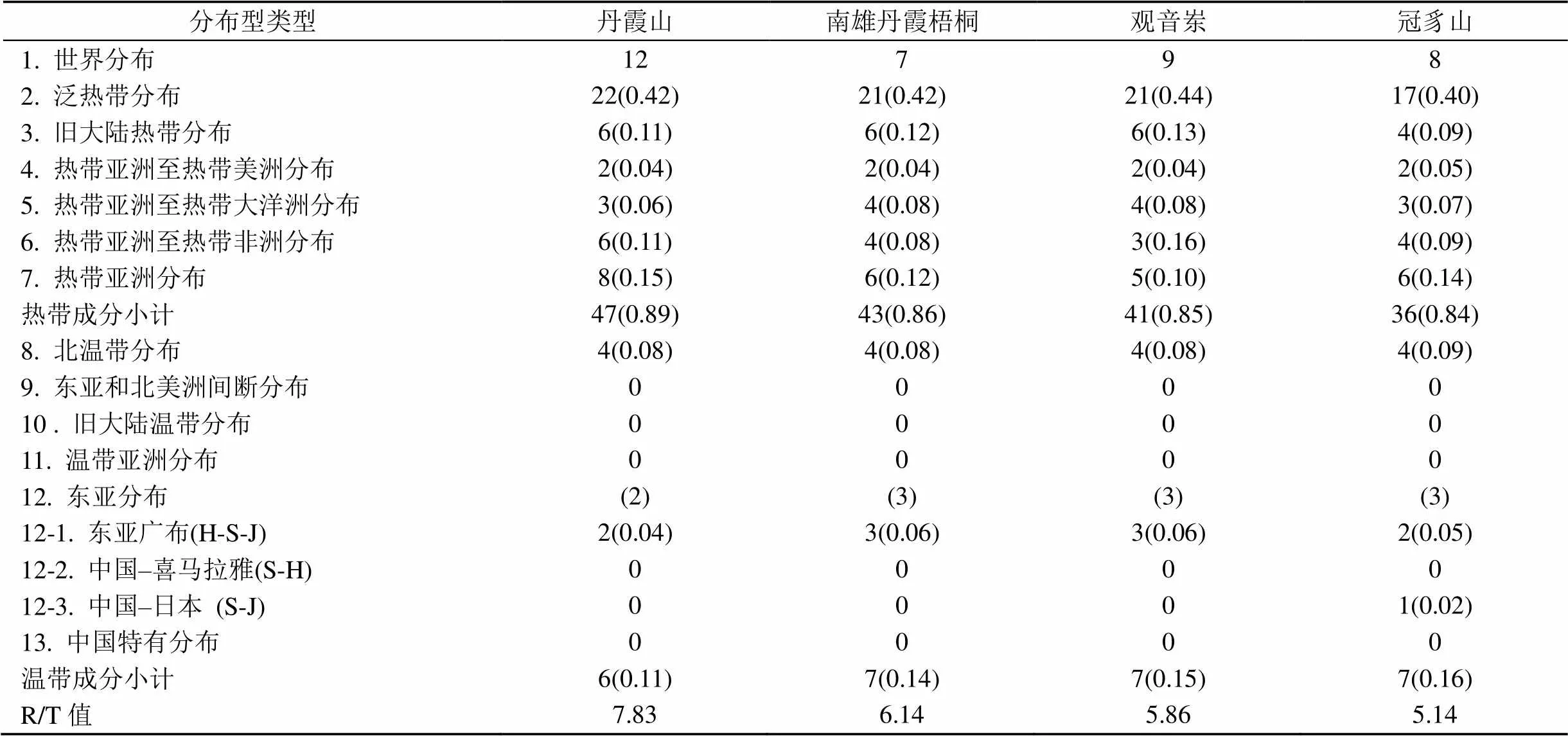
表10 石松类和蕨类植物区系属的分布型
<
注:A(B), A. 该分布区类型的数目, B. 占区系总数的比例, 比例计算不包含世界分布。

表11 石松类和蕨类植物区系种的分布型
<
注:A(B), A. 该分布区类型的数目, B. 占区系总数的比例, 比例计算不包含世界分布。
4.3 区系地理成分共性
从区系地理成分来看, 研究区石松类和蕨类植物与热带亲缘程度较高, 区系地理联系广泛, 大部分区域石松类和蕨类植物区系存在10个分布类型, 呈现出一定程度的纬度地带性特征。科属水平主要以泛热带分布、热带亚洲分布成分为主(在科上占68%以上, 在属上占54%以上), 温带成分仅占11%以上。种水平主要以热带亚洲分布(占43%以上)、东亚成分分布为主(占26%以上), 温带成分占比有所增加(占35%以上), 呈现明显的温带过渡与渗透性质。在种水平上中国–日本分布为东亚分布成分中的主体内容(占64%以上), 这也充分说明了研究区蕨类植物与日本蕨类有着密切的地理联系。
由于岭南山地地区是古热带植物区印度–马来亚区的一部分, 是热带过渡到亚热带的过渡带, 使得研究区蕨类植物热带属性明显。研究区在属水平上无中国特有分布, 特有现象不明显, 但在种水平上中国特有分布不断增多(占6%以上), 研究区大多位于中国东西走向的南岭山脉, 为中国自然地理和植物区系的主要分界线, 是华南地区生物多样性和物种的进化中心[31], 区系成分存在着丰富的交流, 出现福建观音座莲华南凤尾蕨抱石莲等中国特有分布, 说明丹霞地貌区特殊地貌与生境条件为特有成分的形成和保存创造了有利条件, 这对于丹霞地貌区蕨类植物区系组成具有重要意义。
4.4 区系相似性比较
从地理亲缘关系来看, 相似性分析表明研究区间的地理联系密切, 与现代自然条件相似的事实相吻合, 科相似性系数为0.8以上, 属相似性为0.7以上, 种相似性水平0.48以上, 可见研究区区系间的历史联系强于近代自然地理联系[32], 其中南雄丹霞梧桐与观音岽表现出较强的亲缘关系, 特别是在种水平(相似性水平为0.83)明显区别于一般区域, 这表明它们不仅具有共同的起源而且现代区系也有较高的同质性。受同纬度典型丹霞地貌区的限制, 比较研究样本选取有限, 但通过南雄丹霞梧桐(丹霞地貌)与观音岽(非丹霞地貌)的亲缘关系可以说明丹霞地貌对蕨类区系地理成分影响有限, 其区系成分与周边地区联系更为密切。
5 结论
(1)研究区分布有石松类和蕨类植物38科80属206种, 分布的石松类和蕨类植物起源古老, 进化关系连贯, 优势科明显但属种分化程度不高。
(2)非丹霞地貌区丰富性高于丹霞地貌区, 这与丹霞地貌独特的立地条件有关, 丹霞地貌产生了一定程度的生境狭窄特有现象。
(3)研究区石松类和蕨类植物的区系起源与热带地区的蕨类植物区系之间存在着较强亲缘关系, 区系成分以热带成分占主导地位, 从科属水平到种的水平, 温带成分占比从13%增加至35%以上, 呈现向温带过渡特征, 中国–日本成分占比从0%增加至16%以上, 研究区蕨类植物与日本联系密切。
(4)丹霞地貌对相近纬度石松类和蕨类植物区系的影响有限, 区系成分与周边邻近地区交流密切, 相似程度较高, 科、属、种相似性系数分别在0.8、0.7、0.48以上。
致谢:感谢南雄林业局钟平生主任、中南林业科技大学风景园林学院谢禄山副教授在野外植物调查中给予的帮助, 感谢中南林业科技大学风景园林学院硕士研究生周冬梅、王明佳在植物数据库资料整理时提供的帮助。
[1] 严岳鸿, 何祖霞, 马其侠, 周喜乐, 陆奇勇. 湖南丹霞地貌区蕨类植物多样性[J]. 生物多样性, 2012, 20(4): 517–521.
[2] Kuota C. On the Subdivisions of the Red Beds of South Eastern China[J]. Bulletin of the Geological Society of China, 1938, 18: 315–316.
[3] 欧阳杰, 朱诚, 彭华. 丹霞地貌的国内外研究对比[J]. 地理科学, 2011, 31(8): 996–1000.
[4] 彭华. 中国丹霞地貌研究进展[J]. 地理科学, 2000(3): 203–211.
[5] 欧阳杰, 彭华, 罗晓莹, 陈再雄, 张安新, 马育宣. 丹霞山国家珍稀濒危保护植物丹霞梧桐空间分布的微地貌环境特征研究[J]. 地理科学, 2017, 37(10): 1585–1592.
[6] 欧阳杰. 广东丹霞山丹霞地貌类型空间分布初探[J]. 城市地理, 2015, (1): 16–17.
[7] 王卫, 杨俊杰, 罗晓莹, 等. 基于Maxent模型的丹霞山国家级自然保护区极小种群植物丹霞梧桐的潜在生境评价[J]. 林业科学, 2019, 55(8): 19–27.
[8] 臧得奎. 中国蕨类植物区系的初步研究[J]. 西北植物学报, 1998(3): 148–154.
[9] 陈宝明, 李静, 彭少麟, 等. 中国南方丹霞地貌区植物群落与生态系统类型多样性初探[J]. 生态环境学报, 2008, 17(3): 1058–1062.
[10] 吴征镒, 孙航, 周浙昆, 等. 中国种子植物区系地理[J]. 生物多样性, 2011, 19(1): 148.
[11] 严岳鸿, 卫然, 舒江平, 张宪春. 通过现存蕨类植物多样性透视陆生植物的演化[J]. 生物多样性, 2019, 27(11): 1165–1171.
[12] 蒋仁菲, 徐成东, 冯建孟. 滇西北地区蕨类植物物种多样性和区系组成[J]. 生态科学, 2015, 34(5): 71–76.
[13] 严岳鸿, 张宪春, 马克平. 中国蕨类植物多样性与地理分布[M]北京: 科学出版社, 2013: 29–75.
[14] 张开梅, 沈羽, 周晓丽, 方炎明. 21世纪以来蕨类植物研究论文的发表情况: 基于Web of Science的数据统计[J]. 生物多样性, 2019, 27(11): 1245–1250.
[15] 姬红利, 詹选怀, 张丽, 等. 山脉石松类和蕨类植物多样性及生物地理学特征[J]. 生物多样性, 2019, 27(11): 1251–1259.
[16] LI C L, SHEN N H. Species Diversity of Rock Plant Community in Danxia Landform of Zhejiang Province[J]. Journal of Landscape Research, 2020, 12(2): 77–79.
[17] 彭华, 刘盼, 张桂花. 中国东南部丹霞地貌区小尺度植被分异结构研究[J]. 地理科学, 2018, 38(6): 944–953.
[18] 陈功锡, 杨斌, 邓涛, 等. 中国蕨类植物区系地理若干问题研究进展[J]. 西北植物学报, 2014, 34(10): 2130–2136.
[19] 薛高亮, 王韶敏, 苟光前, 郑开颜, 魏奇. 黔北丹霞地貌蕨类植物初步研究[J]. 山地农业生物学报, 2011, 30(2): 110–114.
[20] 彭少麟. 广东丹霞山动植物资源综合科学考察[M]. 北京: 科学出版社, 2011.
[21] 曾宪锋, 邱贺媛, 钱会莲, 庄雪影, 唐光大. 粤东蕨类植物区系初步研究[J]. 广西植物, 2009, 29(4): 466–470.
[22] 中国科学院中国植物志编辑委员会. 中国植物志(第一卷)[M]. 北京: 科学出版社, 2004: 78–92
[23] 福建植物志编写组. 福建植物志(第一卷. 修订本)[M]. 福州: 福建科学技术出版社, 1991: 5–264.
[24] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊Ⅳ): 1–139.
[25] 李承森. 植物科学进展 (第六卷)[M]. 北京: 高等教育出版社, 2004.
[26] 沈泽昊, 胡会峰, 周宇, 等. 神农架南坡植物群落多样性的海拔梯度格局[J]. 生物多样性, 2004, 12(1): 99–107.
[27] 张镱锂. 植物区系地理研究种的重要参数—相似性系数[J]. 地理研究, 1998, 17(4): 429–433.
[28] 左家哺. 植物区系基本特征的参数综合表达[J]. 武汉植物学研究, 1993(4): 300–306.
[29] 李登武, 陕北黄土高原维管植物区系地理研究[D]. 咸阳: 西北农林科技大学, 2008.
[30] 彭少麟, 李富荣, 周婷, 等. 丹霞地貌沟谷生态效应[J]. 生态学报, 2008, 28(7): 2947–2953.
[31] 田园, 龚粤宁, 邓素玲. 南岭华南物种进化中心[J]. 森林与人类, 2014(10): 18–24.
[32] 陈开森, 吴锦平. 福建梅花山自然保护区蕨类植物区系研究[J]. 西北植物学报, 2019, 39(6): 1121–1126.
Comparative study of lycophytes and ferns on Danxia landform
LIANG Sheng, WANG Mengnan, WEI Baojing, HU Xijun*, CHEN Cunyou
College of Landscape Architecture, Central South University of Forestry and Technology, Changsha 410004, China
Based on field work and collecting literature, we investigated the basic composition, floristic composition and correlation of lycophytes and ferns in similarly latitude Danxia Landform areas of Fujian and Guangdong Province, as well as compared the differences between Danxia landform and Non-Danxia landform. Our results are as follows. (1) The are totally 206 species, belonging to 38 families and 80 genera, with 134 species belonging to 35 genera and 65 families in Danxia Mountain, 103 species belonging to 31 genera and 51 families in Guanzhai Mountain, 103 species belonging to 31 genera and 57 families in Guanyin Dong, and 96 species belonging to 31 genera and 57 families in Nanxiong Danxia Wutong. The pteridophytes in the study area have ancient origins and continuous evolutionary relationships, and the predominant families are obvious but the genus is not highly differentiated. (2) On floristic richness, the Non-Danxia landform area has a higher richness than the Danxia Landform area, which is shown as Danxia Mountain (0.74) > Guanyin Dong (Non-Danxia landform) (0.14) > Nanxiong Danxia Wutong (0.07) > Guanzhai Mountain (0.02). (3) On floristic geographical components, the genus level is dominated by tropical components (accounting for more than 84%), with a high degree of kinship with the tropics. At the species level, the tropical components accounted for more than 59%, showing a transition from tropical to temperate characteristics. (4) On geographical relatedness, Nanxiong Danxia Wutong (withDanxia landform) and Guanyin Dong (withNon-Danxia landform) have obvious relatedness. The similarity of family, genus and species is 0.88, 0.97 and 0.8 respectively, indicating that Danxia landform has limited influence on the fern flora. The results provide an important basis for studying the origin and development of lycophytes and ferns in Danxia landform and provide a reference for the rational development and utilization of plant resources in Danxia landform.
Danxia landform; lycophytes and ferns; floristic composition; correlation analysis
10.14108/j.cnki.1008-8873.2022.02.020
S157.2
A
1008-8873(2022)02-166-08
2020-03-01;
2020-04-18
国家林业局野生植物保护项目(2016ZWZY06); 林业公益性行业科研专项(201404710); 湖南省高等学校“双一流”建设项目(湘教通〔2018〕469号)
梁胜(1994—), 硕士, 主要从事风景园林规划与设计, E-mail: liangsheng0904@foxmail.com
通信作者:胡希军(1964—), 博士, 教授, 博士生导师, 主要从事风景园林规划与设计、景观规划与生态修复方向研究, E-mail: 120795043@qq.com
梁胜, 王梦楠, 韦宝婧, 等. 丹霞地貌石松类和蕨类植物区系比较研究[J]. 生态科学, 2022, 41(2): 166–173.
LIANG Sheng, WANG Mengnan, WEI Baojng, et al. Comparative study of lycophytes and ferns on Danxia landform[J]. Ecological Science, 2022, 41(2): 166–173.
猜你喜欢
天天爱科学(2022年4期)2022-05-23
大众文艺(2021年21期)2021-11-25
炎黄地理(2021年8期)2021-10-22
源流(2021年1期)2021-07-28
岭南音乐(2019年6期)2019-12-31
人民之声(2019年5期)2019-12-23
艺术家(2019年4期)2019-04-20
神州民俗(2018年9期)2018-11-21
广东第二课堂·初中(2017年2期)2017-03-15
快乐作文·低年级(2016年12期)2017-01-03
