
巴拉圭瓜多竹适生区分布研究
2018-10-25 02:02:12杨宇明杨德军谷中明张快富赵一鹤
竹子学报 2018年2期
陈 剑,杨宇明,王 娟,杨德军,谷中明,张快富,赵一鹤
(1.云南省林业科学院,云南 昆明 650201; 2.国家林业局云南珍稀濒特森林植物保护和繁育重点实验室,云南省森林植物培育与开发利用重点实验室,云南 昆明 650201; 3.云南大学,云南 昆明 650091;4.西南林业大学,云南 昆明 650224; 5.云南南亚竹藤研究中心,云南 昆明 650201)

图1 云南省景洪市引种的巴拉圭瓜多竹Fig.1 Guadua paraguayana in Jinhong City,Yunnan Province, China
巴拉圭瓜多竹(Guaduaparaguayana)为禾本科(Poaceae)竹亚科(Bambusoideae)箣竹属(Bambusa)瓜多竹亚属(Guadua)的竹种,天然分布于南美洲阿根廷北部、玻利维亚、巴西、巴拉圭和委内瑞拉等地,为中型丛生竹,秆实心,高8~10 m,2 m以上呈攀援状,外形优美(见图1),在原产地常用作农具、篱笆、标杆等用途,在竹地板加工、景观美化等方面有潜在的利用价值。2015年从巴西引进的巴拉圭瓜多竹实生幼苗在云南景洪市和普洱市进行试种,至今总体长势良好。巴拉圭瓜多竹的研究仅见基因克隆方面[1-2],还未见对该竹种进行引种试种等方面的报道。瓜多竹属其它竹种我国已有少量引种,已有研究报告的同属竹种是大型丛生竹Guaduaamplexifolia,已于2002年从南美洲引种至福建省华安县,并于近年引种至云南省普洱市和景洪市,均表现出良好的适应性,在种苗扩繁[3]、幼林结构调控[4]、光合特性[5]、低温胁迫[6]、Nacl胁迫[7]等方面已有相关研究。但目前尚未见瓜多属竹种在大尺度上的适生区分布研究,通过科学手段选择最适合竹种生长发育所需气候条件的适生区,可避免或减少将来产业化种植时面临的风险,为规模化育苗种植选址提供科学依据。
最大熵原理(the principle of maximum entropy)起源于信息论和统计力学, 是基于有限的已知信息对未知分布进行无偏推断的一种科学方法,可基于气候、海拔、植被等环境因子构建物种的地理分布[8]。MaxEnt模型即基于最大熵原理进行构建的模型,结合已知物种分布地的各种生态环境因子,对物种样本采集地以外的潜在分布区域进行预测,具有很高的预测准确度[9],尤其是在小样本数据条件下获得的结果比其它模型更为优越[10-11]。MaxEnt模型在国际上已经获得非常广泛的运用,如栗树根蚜虫潜在分布区预测[12]、造礁珊瑚玮度分布范围[13]、章鱼的分布区预测和实证[14]等,在我国,近年来基于MaxEnt模型对物种分布区分析和预测的研究极为流行,近年来基于MaxEnt模型的物种适生区分析就有黄顶菊[15]、非洲橘硬蓟马[16]、川贝母[17]等几十个物种的分布区预测。
巴拉圭瓜多竹天然分布南美洲,在缺乏其它地方引种研究的情况下,MaxEnt模型是研究巴拉圭瓜多竹在原产地外的适生区的有力工具。本研究根据巴拉圭瓜多竹在南美洲的分布样点和在中国引种情况,利用MaxEnt模型结合地理气候变量,对巴拉圭瓜多竹在世界范围内潜在适生区进行梯度划分和可视化分析,为巴拉圭瓜多竹的引种和利用提供前期的科学参考。
1 材料与方法
1.1 巴拉圭瓜多竹分布数据的来源与处理
本研究中巴拉圭瓜多竹分布点主要通过全球生物多样性信息数据库(http://www.gbif.org/)进行下载,除去重复记录的分布点,获得22个分布样点(如图2),并在中国云南景洪市和普洱市巴拉圭瓜多竹种植基地实地采集地理信息样点3个,共计采集分布样点25个(如图3),在Excel软件中按物种、经度、纬度的顺序制作表格转化为.CSV文件,西经、南纬的值为负,经纬度为十进制小数格式。
1.2 环境数据
环境数据变量从世界气候数据库(www.worldclim.org)上下载,空间分辨率为5 弧分,19个气候变量及其含义如表1所示。
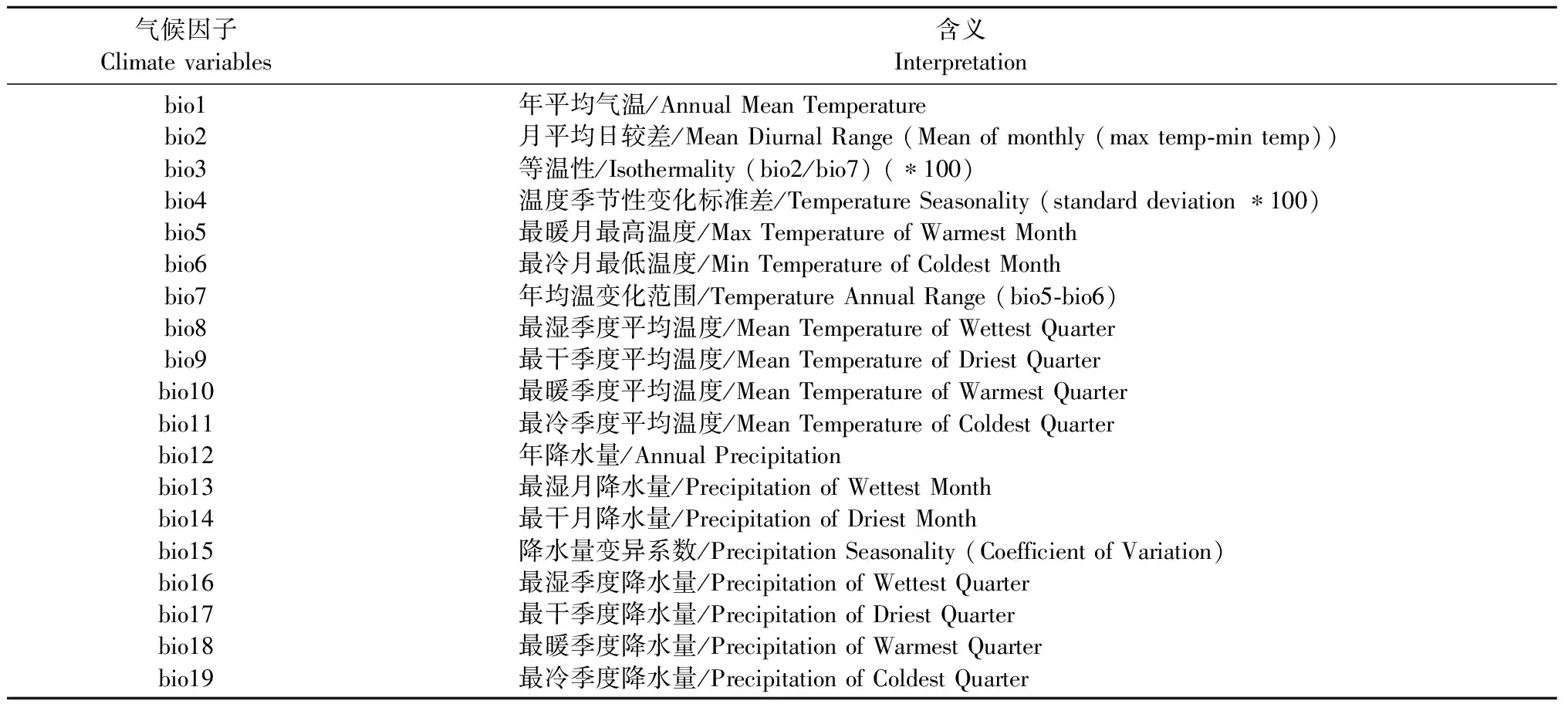
表1 19个环境因子及其含义Tab.1 19 climate variables and their interpretation
世界范围内的分析底图为世界地图和世界行政区划矢量图(http://nfgis.nsdi.gov.cn)。
1.3 软件及使用
本研究主要使用的建模与分析软件为MaxEnt V3.3.3(http://www.cs.princeton.edu/~schapire/maxent/)和arcigis10.2,MaxEnt V3.3.3是基于JAVA平台设计的物种适生区预测模型,运算前需要安装JAVA插件支持,通过物种分布点与环境变量建立适生区预测模型,并得到每个象元的适生区指数(概率),取值范围为0%~100%;模型运行时随机选取75%的分布点用于模型建立,25%的分布点用于模型验证;使用刀切法对环境因子权重进行分析,获得影响物种分布权重最大的环境变量;用受试者工作特征曲线 (ROC 曲线) 下面积AUC 值对模型模拟的准确性进行评估;分析结果用arcgis10.2进行自然分级进行图像处理,得到可视化的适生区梯度分级。
1.4 云南引种样点的利用
鉴于巴拉圭瓜多竹在云南景洪、普洱的3个分布样点5年来长势良好,故将该3个分布样点作为可能的适生区样点加入世界分布样点进行分析讨论。
2 结果与分析
2.1 巴拉圭瓜多竹的潜在适生区
基于巴拉圭瓜多竹在南美洲的22个分布样点和全球气候因子,通过MaxEnt建模分析统计得到的巴拉圭瓜多竹适生区分布,经arcgis 10.2进行分级处理后如图2所示:0.530 1~0.907 2为高适生区(黑色),0.256 2~0.530 1次适生区(深灰色),0.067 6~0.256 2为低适生区(浅灰色),0~0.067 6为非适生区(白色)。
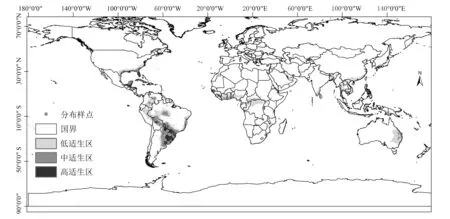
图2 巴拉圭瓜多竹南美洲采样点和适生区分布Fig.2 The sampling sites of Guadua paraguayana in South American and its potential distribution area
(1)巴拉圭瓜多竹适生区主要集中在南美洲。巴西东部的伊亚州(Bahia)东部、圣埃斯皮里图州(Espirito Santo)、里约热内卢州(Rio De Janeiro),南部的圣保罗州(Sao Paulo)南部、南马托格罗索州(MatoGrosso Do Sul)、巴拉那州(Parana)、圣卡塔琳娜州(Santa Catarina)、南里奥格兰德州(Rio Grande do Sul)是巴拉圭瓜多竹的高适生区;其次,巴西南部,中西部、东部也有大面积的中适生区和低适生区。巴拉圭除西部有部分非适生区外,其余国土均为适生区,自上巴拉圭省(Alto Paraguay)以东以南的大部分国土均为高适生区。阿根廷东北部自西向东为低中高适生区镶嵌格局,米西奥内斯省(Misiones)、科连特斯省(Corrientes)全境为高适生区;毗邻的恩特雷里奥斯省(Entre Rios)、圣塔菲省(Santa Fe)、查科省(Chaco)、福莫萨省(Formosa)各省中至东部均为高适生区,中到西部则为中到低适生区;布宜诺斯艾利斯省(Buenos Aires)几乎全境大部分均为中到低适生区、北部有少量高适生区。乌拉圭全境为适生区,其中中至西北部为高适生区,其余部分为中适生区。玻利维亚中北部为适生区,其中圣克鲁斯省(Santa Cruz)、科恰班巴省(Cochabamba)、埃尔贝尼省(El Beni)3省交界及附近区域均为高适生区。另,秘鲁,哥伦比亚、圭亚那等国也有大面积的中低适生区。
(2)其它地区的适生区分布情况。整个墨西哥湾沿岸地区均为从低到高各级适生区。美国毗邻墨西哥湾的州沿海区域均为低到高适生区,其中高适生区集中分布在佛罗里达州(Florida)和路易斯安娜州(Louisiana);墨西哥毗邻墨西哥湾的维拉克鲁斯州(Veracruz)、塔巴斯科州西侧和东侧(Tabasco)、坎佩切州西南部(Campeche)、瓦哈卡州东北部(Oaxaca)为高适生区。中美洲危地马拉(Guatemala)中到北部依次为低中高适生区格局;洪都拉斯(Honduras)大部分国土为中适生区,尼加拉瓜(Nicaragua)大部分国土为低适生区。西印度洋群岛均为中到高适生区,其中高适生区主要分布于巴哈马(Bahamas)和古巴(cuba)。非洲中部从利比里亚(Liberia)到乌干达(Uganda)东西方向上有带状的低到中适生区;东部从莫桑比克(Mozambique)的赞比西省(Zambezia)到南非的西开普敦(Western Cape)沿海地区也有呈带状分布的适生区,高适生区主要分布在赞比西亚中部和夸祖鲁-纳塔尔省(KwaZulu-Natal)和东开普敦(Eastern Cape)沿岸地区;马达加斯加东侧沿海地区为中低适生区。大洋洲澳大利亚适生区主要分布在昆士兰州(Queensland)和新南威尔士州(New South Wales),以低到中适生区为主,高适生区分布在沿海地区;新西兰北部有低适生区分布;巴布亚新几内亚南部为高中低适生区镶嵌;斐济、新喀里多尼亚、瓦努阿图等群岛为中到高适生区。亚洲适生区分布情况为:南亚印度南部有低到中适生区;斯里兰卡西部有高中低适生区镶嵌格局;东南亚诸群岛中,分布在印度尼西亚的主要是低适生区;菲律宾南部有低适生区,中部为中适生区,北部主要为中高适生区;泰国中部为中高适生区,北部以低适生区为主,在清迈省(Chiang Mai)与达府省(Tak)交界的区域为高适生区;柬埔寨中到南部全部为中低适生区;越南南北向上分布有中低适生区,北部基本为非适生区;中国海南省中到东部分别为低到中适生区,台湾省北部及东部沿海地区主要为中适生区、北部有零星高适生区,福建省、广东省、广西省有大面积连片的低适生区,云南省西双版纳州中到南部为低适生区。
2.2 将云南分布样点加入后巴拉圭瓜多竹适生区变化
如图3所示,加入巴拉圭瓜多竹在云南的3个分布样点后,MaxEnt模型得到的适生区在世界范围内的分布格局主要变化体现在保持原有分布格局的基础上,高中低3个级别的适生区都有所增加,在东南亚的变化最为明显。0.514 1~0.923 2为高适生区(黑色),0.231 7~0.514 1次适生区(深灰色),0.061 5~0.231 7为低适生区(浅灰色),0~0.061 5为非适生区(白色)。
中国范围内,云南省由原来的西双版纳州仅有的低适生区变为整个云南中至南部均为中低高变化的适生区,而西双版纳范围内大部分地区为高适生区;福建省、广东省、广西省的低适生区面积比原来扩大,且中适生区面积显著增加;海南省由原来的中到东部的中低适生区变为几乎全境均为高适生区;台湾西部增加了中低适生区。
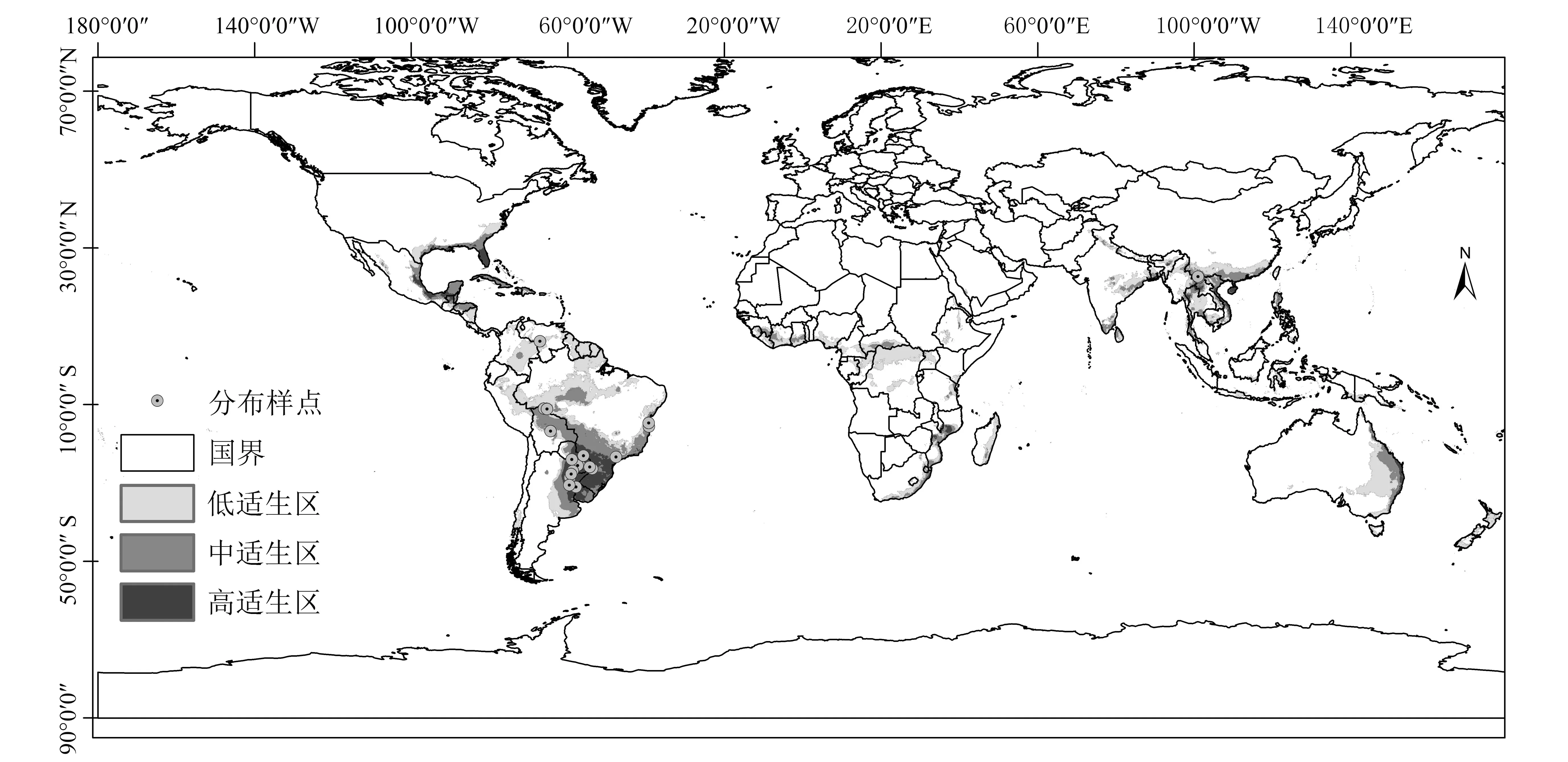
图3 加入云南分布样点后巴拉圭瓜多竹的适生区分布Fig.3 The potential distribution area of Guadua paraguayana after considering the sampling sites in Yunnan Province, China
2.3 影响巴拉圭瓜多竹适生区分布的气候变量
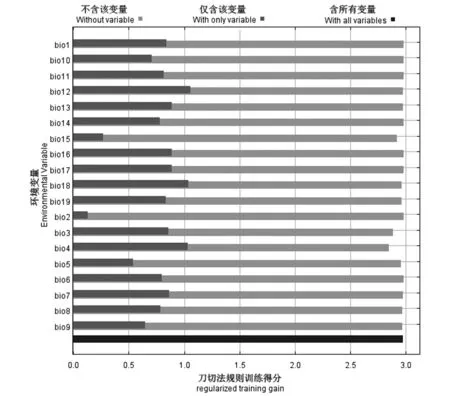
图4 刀切法环境变量重要值Fig.4 Importance analysis of environmental variables by jackknife test
图4是MaxEnt模型对19个环境变量进行刀切法分析的结果,深灰色条带表示单个环境变量对物种适生区分布贡献的权重,浅灰色条带表示除了该变量以外的其他所有变量综合起来对物种分布的贡献权重,黑色条带表示所有变量对物种分布贡献权重,19个环境变量中,刀切法环境变量重要值权重排在前3名的变量分别为bio12(年降水量)、bio18(最暖季度降水量)、和bio4(温度季节性变化标准差)。这3个变量值权重差别不大,均在1.000~1.105 3之间。根据物种分布概率对气候因子的响应曲线来看,当物种分布概率大于0.5时,年降水量在1 125~3 500 mm、最暖季降水量大于330.35 mm、温度季节变化标准差在1 354.17~4 375.00之间时,物种分布对这3个气候变更有最显著的响应。说明充足的降水、温度季节变化较小是影响瓜多瓜多竹分布的关键气候变量。
当bio15(降水量变异系数)、bio3(等温性)、bio4(温度季节性变化标准差)等3个气候变量缺失时,对物种的分布有较明显的影响,说明这3个气候变量对瓜多竹的分布有潜在的影响,可能体现在较小的季节性降水量变化和较小的气温波动,更适宜瓜多瓜多竹的分布。
bio2(月平均日较差)、bio5(最暖月最高温度)对瓜多竹分布所起的作用权重最低,且除去他们时对权重的影响不大,说明这2个气候变量对瓜多竹分布影响程度最低。
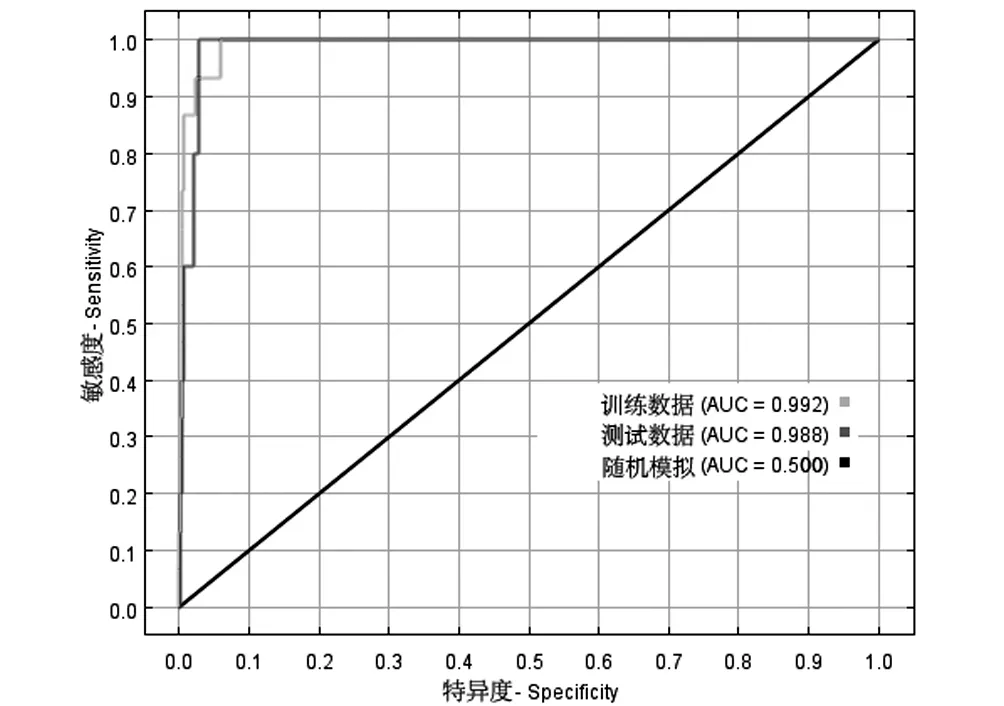
图5 ROC曲线Fig.5 ROC curve
2.4 模型精度分析
MaxEnt模型精度用受试者工作特性曲线即ROC曲线(receiver operating characteristic curve)进行评价,ROC曲线是根据一系列不同的二分类方式(分界值或决定阈),以真阳性率(灵敏度)为纵坐标,假阳性率(特异度)为横坐标绘制的曲线。
ROC 曲线以真阳性率(实际存在,且被预测为存在的比率)为纵坐标,以假阳性率(实际不存在,但被预测为存在的比率)为横坐标,AUC值指ROC曲线与横坐标围成的面积值,值域范围为0~1,AUC值越大表示与随机分布相距越远,环境变量与预测的物种地理分布之间的相关性越大,即模型预测效果越好,反之说明模型预测效果越差。 AUC 评价指标[18]:很差(≤0.060),较差(0.600~0.700),一般(0.700~0.800),好(0.800~0.900),极好(0.900~1.000)。
如图5所示,本研究中训练数据的AUC值为0.992,测试数据的AUC值为0.988,说明模型的预测准确度很好。
3 结论和讨论
3.1 巴拉圭瓜多竹适生区概况
本研究利用巴拉圭瓜多竹在南美洲和中国云南的分布样点、世界气候因子数据为基本数据,通过MaxEnt模型获得了巴拉圭瓜多竹在世界范围内的适生区。巴拉圭瓜多竹适生区主要集中分布在南美洲,在中美洲、非洲中部、大洋洲、东南亚的部分地区也有分布,影响巴拉圭瓜多竹分布的气候变量主要是年降水量、最暖季度降水量、温度季节性变化标准差。在中国范围内,仅用南美洲分布样点数据时,MaxEnt模型获得的结论是云南省、福建省、广东省、广西省的部分地区为低适生区,海南省、台湾省部分地区有低到中适生区;加入云南省引种分布样点后,以上各省的适生区范围均有明显的增长,其中,云南西双版纳和海南省是潜在的高适生区,适于进行产业化种植。
3.2 加入云南分布点后巴拉圭瓜多竹适生区增加的原因
生态位是衡量物种与环境变量的相互作用的一个重要指标,可表述为“一个物种生存和繁殖的特定环境变量的区间[19-20],基于MaxEnt模型的适生区研究通常也称作基于该物种的生态位研究[21-22],在物种分布信息有限的情况下,MaxEnt模型可有效地分析出与原分布点环境生态位最为匹配的潜在适生区。加入中国云南的分布样点后,巴拉圭瓜多竹的适生区分布范围有所增加,尤其在东南亚最为明显,原因可能是巴拉圭瓜多竹对气候因子的适应范围比原分布地更为宽广,在数据缺乏的情况下,仅用南美洲的分布样点不足以充分体现该物种能适应的环境因子变化范围,而加入更多的分布样点拓展了原来有限的生态位区间。5年时间还不足以判断巴拉圭瓜多竹是否能在云南已经引种成功,比如繁殖能力、生命周期等生理特征还需要进一步观察,因此加入云南分布点后新增加的适生区范围是一个可能的参考信息。
3.3 与适生区相关的风险
巴拉圭瓜多竹的适生区范围内,还应注意生态风险的控制,有研究认为它是具有很强入侵能力的植物[23-24],天然状态下通过开花结籽形成的实生苗数量大,长成以后如同一堵带刺的墙,人畜难以进入,给生态环境带来潜在的危害,需要在种植时尤其是产业化种植时采取防范措施。
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
廉政瞭望·下半月(2023年8期)2023-09-07 12:37:55
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
西北植物学报(2022年3期)2022-04-18 01:40:50
电脑爱好者(2020年10期)2020-07-28 17:10:30
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
电脑爱好者(2019年16期)2019-10-30 03:35:12
中国果业信息(2018年1期)2018-01-17 23:36:16
农药科学与管理(2018年5期)2018-01-17 12:26:04
青岛科技大学学报(社会科学版)(2015年4期)2016-01-25 09:57:38
