
银杏叶提取物通过Nrf2∕HO-1信号通路减轻老年性聋大鼠耳蜗氧化应激损伤的机制
2022-04-08 06:33王青玲张梦娴周颍东郭向东康浩然王庆林
实用医学杂志 2022年3期
王青玲 张梦娴 周颍东 郭向东 康浩然 王庆林
1河南中医药大学第一临床医学院(郑州450046);2河南中医药大学第一附属医院耳鼻咽喉科(郑州450000);3河南医学高等专科学校护理系(郑州451191)
老年性聋是由机体老化引起的双耳对称性、进行性感音神经性耳聋,听力损失从高频开始,逐步向中、低频发展[1]。我国第二次残疾人抽样调查发现老年性聋的人数高达949 万,且患病率逐年攀升[2]。目前,老年性聋的治疗手段主要包括人工耳蜗植入、佩戴助听器和药物治疗等[3],但人工耳蜗植入和佩戴助听器成本过高,而现有的治疗药物临床疗效欠佳[4-5],因此,从老年性聋的发病机制入手、寻求有效治疗的药物是目前临床上亟需解决的关键问题之一。老年性聋的发病机制可能与氧化应激和线粒体功能障碍引起的内耳毛细胞和螺旋神经节细胞退行性变性有关[6-7]。核因子红系相关因子2(Nrf2)是调节细胞氧化应激和线粒体稳态的重要转录因子,氧化应激状态下,Nrf2 移位到细胞核内,促进下游血红素加氧酶1(HO-1)等抗氧化因子表达,进而降低耳蜗中氧自由基(ROS)水平,从而保护内耳细胞免受氧化应激损伤[8]。银杏叶提取物(ginkgo biloba extract,GBE)是从银杏干燥的叶中提取的有效成分,含有银杏黄酮、槲皮素、银杏内酯、有机酸等多种抗氧化成分[9],其可通过降低ROS 累积,提高线粒体氧化呼吸链复合体Ⅰ、Ⅳ和Ⅴ的活性和氧化磷酸化水平,进而对多种内耳疾病、心脑血管疾病和神经退行性疾病发挥保护作用[10-11],但有关GBE 治疗老年性聋机制方面的研究相对较少,为此,本研究采用D-半乳糖制备老年性聋模型,应用多种实验方法探讨GBE 能否通过调节Nrf2∕HO-1 信号通路对老化耳蜗发挥保护作用。
1 材料与方法
1.1 实验动物 从河南华兴动物养殖中心购买4 周、体质量150 ~250 g、听阈正常的雄性SD 大鼠45 只,于SPF 级饲养环境中饲养,室内温度控制在(24 ± 3)℃,自由饮食和摄水,实验前适应性饲养7 d,实验动物许可证号:SYXK(豫)2020-0002。
1.2 试剂 GBE(北京华润有限公司,Z11021350);D-半乳糖(上海麦克林有限公司,C11484411);TSOD和MDA(江苏宝莱有限公司,MM-0731R1、MM-0385R1);2.5%戊二醛(北京索莱宝公司,SP0041);Nrf2、HO-1 抗体(美国genetex 公司,GTX103322、GTX30748);Myosin-Ⅶa 抗体(美国Protrus BioSciences 公司,25-6790);Alexa Flour 488 抗体(美国Jackson 公司,111-545-003);内参GAPDH 抗体(上海UBIYOBIBIO,UBI6010)。
1.3 方法
1.3.1 模型制备和给药方法 将SD 大鼠依次编为1-45 号,按照随机数字表分为对照组、模型组和GBE 低、中、高剂量组,每组9 只。模型组和GBE各组大鼠均在颈背部皮下注射D-半乳糖,剂量500 mg∕(kg·d),对照组在颈背部皮下注射等量生理盐水,共8 周[12]。造模成功后,GBE 低、中、高剂量组分别腹腔注射GBE[10、20、30 mg∕(kg·d)],对照组及模型组腹腔注射等量生理盐水,连续8 周。
1.3.2 听性脑干诱发电位(ABR) 10%的水合氯醛麻醉大鼠,记录电极置于大鼠的颅顶中点,参考电极置于检测耳耳后,扬声器置于距受刺激侧耳廓10 cm 的位置,地线接对侧耳耳后。TDT RZ6∕BioSigRZ 系统收集ABR,采用click 为刺激音,声刺激从90 dB 开始,10 dB 递减,接近阈值时5 dB 递减,直至能分辨出Ⅲ波的最低刺激强度确定反应阈值。
1.3.3 酶联免疫吸附实验 选取治疗前、后各组大鼠血清在冰上解冻,离心后取上清,按照相应试剂盒中提供的制造商说明测定T-SOD 和MDA 的含量。用黄嘌呤氧化酶法测定T-SOD,用硫代巴比妥酸法测定MDA。
1.3.4 透射电子显微镜观察螺旋神经节细胞超微结构 麻醉后,断头处死,用2.5%戊二醛(pH 7.4)在4 ℃灌注固定24 h,脱钙液脱钙3 周,用显微器械取下基底膜,1%的锇酸4 ℃固定2 h,PBS 漂洗3 次,梯度乙醇,丙酮脱水,将组织包埋在环氧树脂中,进行超薄切片,切片用醋酸铀酰和柠檬酸铅双染色(20 min),染色后,水洗,烘干,使用日本透射电子显微镜(JEM-1400)拍照。
1.3.5 免疫荧光染色基底膜铺片 耳蜗用4%多聚甲醛(PFA)固定过夜,脱钙3 周,在解剖显微镜下将基底膜分离为顶回、中回、底回三段,10%山羊血清封闭通透,4 ℃过夜。Myosin-Ⅶa(1∶500)孵育过夜。PBS 漂洗3 次∕10 min。二抗Alexa Flour 488(1∶500)避光孵育2 h。PBS 漂洗3 次∕10 min,DAPI(1∶800)室温孵育15 min,抗猝灭甘油封片,在荧光显微镜下从耳蜗顶回至底回开始逐个视野连续计数内、外毛细胞数目,并观察毛细胞形态。
1.3.6 免疫组织化学 石蜡切片在60 ℃烘干1 h,二甲苯脱蜡,然后梯度乙醇脱水;使用柠檬酸钠缓冲液在180 ℃下反应90 s;室温冷却后,用3%H2O2溶液封闭30 min;1XPBS 漂洗3 次∕5 min;10%山羊血清封闭30 min;Nrf2 与HO-1 抗体分别用抗体稀释液按照1∶100 比例稀释,在4 ℃下过夜;1XPBS漂洗3 次∕5 min,Bio-羊抗兔IgG 按照1∶100 比例配置工作液,4 ℃孵育1 h;链霉素亲和素-POD 工作液4 ℃孵育1 h;滴加DAB 显色液在显微镜下显色,待组织颜色变为棕褐色,用蒸馏水洗涤终止反应;苏木素轻度复染,脱水,透明,封片,光学显微镜下观察蛋白表达情况并拍照。
1.3.7 蛋白印迹法 称取20 μg 耳蜗组织、剪碎、捣碎、冰上裂解10 min;根据BCA 试剂盒测最终蛋白上样体积。然后制备SDS-PAGE 凝胶、电泳、转膜、1XTBST 洗膜3 次∕5 min、5%脱脂奶粉封闭1 h、洗膜;配制目的蛋白抗体:Nrf2(1∶1 000)、HO-1(1∶1 000)、GAPDH(1∶1 000),在4 ℃过夜,利用HRP 标记的二抗在室温孵育2 h;用ECL 发光液进行显影,Image Lab 成像系统对形成蛋白条带的斑点进行定量,采用GAPDH 作为内参对照,对数据进行统一处理。
1.4 统计学方法 使用SPSS 25.0 软件进行分析,所有数据均以均数±标准差表示,多组数据比较采用单因素方差分析和Dunnett 多重比较检验,所有数据均重复3 次以上,P<0.05 为差异有统计学意义。
2 结果
2.1 ABR 阈值 治疗前,其余各组与对照组相比ABR 阈值均显著提高(P<0.01),表明造模成功;治疗后,GBE 低剂量组与模型组比较ABR 阈值差异无统计学意义(P>0.05),中、高剂量组ABR 阈值均明显降低,差异有统计学意义(P<0.05),见图1。
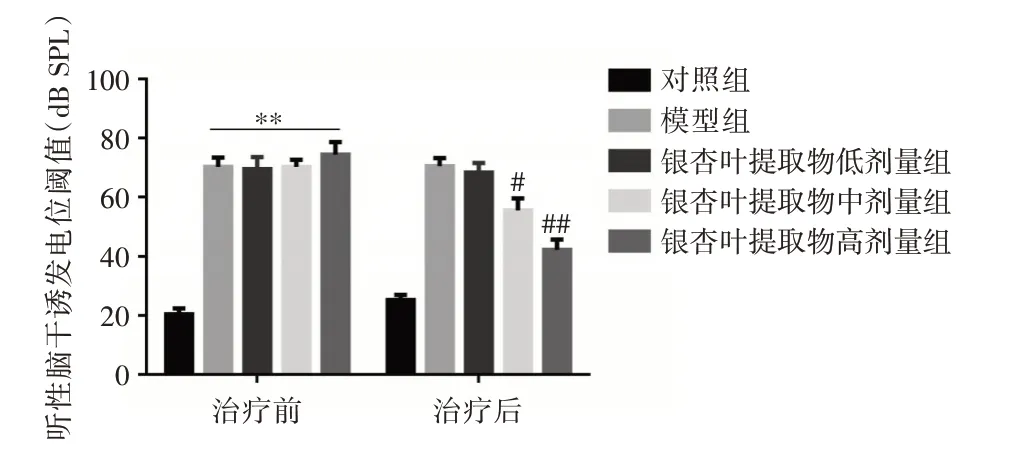
图1 银杏叶提取物治疗前后各组大鼠ABR 阈值变化Fig.1 Changes of ABR threshold in rats before and after Ginkgo biloba extract treatment
2.2 血清中T-SOD 和MDA 的含量 治疗前,与对照组比较,其余各组血清T-SOD 含量显著降低(P<0.01),而MDA 含量明显升高(P<0.01);治疗后,与模型组比较,GBE 各组T-SOD 含量逐渐提高,MDA 含量显著下调(P<0.05),尤以中、高剂量组较明显(P<0.01),见表1。
表1 治疗前后各组血清中T-SOD 和MDA 含量变化Tab.1 Changes of T-SOD and MDA in serum before and after treatment ±s

表1 治疗前后各组血清中T-SOD 和MDA 含量变化Tab.1 Changes of T-SOD and MDA in serum before and after treatment ±s
注:治疗前,与对照组比较,**P <0.01;治疗后,与模型组比较,#P <0.05,##P <0.01
组别对照组模型组银杏叶提取物低剂量组银杏叶提取物中剂量组银杏叶提取物高剂量组T-SOD 含量(pg∕mL)治疗前440.09±36.40 295.78±28.16**300.64±14.26**283.04±27.35**306.42±36.40**治疗后441.03±39.36 298.89±29.36**326.84±38.12#382.37±20.36##408.65±35.14##MDA 含量(nmol∕mL)治疗前5.60±0.44 11.33±0.42**10.06±0.23**11.42±0.50**10.96±0.54**治疗后5.67±0.35 10.92±0.53**8.95±0.42#6.93±0.35##6.18±0.55##
2.3 螺旋神经节细胞的超微结构 对照组螺旋神经节细胞超微结构正常,线粒体外膜和嵴清晰,髓鞘板层结构紧密,板层呈同心圆排列,边缘光滑;模型组螺旋神经节细胞出现明显衰老现象,线粒体数量减少,密度降低,大量线粒体空泡样变、双层膜破裂、嵴模糊不清,螺旋神节细胞胞浆内溶酶体聚集,髓鞘板层排列疏松、紊乱,甚至出现“脱髓鞘现象”。经不同剂量GBE 治疗后,螺旋神经节细胞呈现剂量依赖性改善,胞浆内空泡化的线粒体数量逐渐降低,线粒体外膜和嵴形态逐渐恢复,髓鞘板层排列规整紧密。见图2。

图2 各组耳蜗螺旋神经节细胞超微结构变化Fig.2 Ultrastructural changes of cochlear spiral ganglion cells in each group
2.4 基底膜内、外毛细胞的形态和数量 对照组内、外毛细胞形态清晰,呈圆形或矩形,排列整齐,未见明显病理损伤;模型组中,顶回内、外毛细胞基本正常,中回内、外毛细胞丢失不明显,仅出现外毛细胞形态大小不一,底回内、外毛细胞排列紊乱,形态极其不规整,内、外毛细胞均出现大量丢失,外毛细胞丢失更严重(P<0.05);与模型组相比,GBE 各组内、外毛细胞结构逐渐清晰,形态逐渐改善,底回内、外毛细胞丢失数量明显降低,高剂量组改善较明显(P<0.05)。见图3、表2。

图3 免疫荧光染色观察基底膜顶、中、底回毛细胞损伤情况(×400)Fig.3 Observation of hair cells damage in Apical turn,Middle turn,Basal turn of basement membrane by immunofluorescence staining(×400)
表2 基底膜内、外毛细胞的数量Tab.2 Number of inner and outer hair cells in basement membrane ±s

表2 基底膜内、外毛细胞的数量Tab.2 Number of inner and outer hair cells in basement membrane ±s
注:与对照组比较,*P <0.05,**P <0.01;与模型组比较,#P <0.05,##P <0.01
组别对照组模型组银杏叶提取物低剂量组银杏叶提取物中剂量组银杏叶提取物高剂量组内毛细胞顶回21.20±1.36 20.93±1.76 20.96±1.21 21.15±0.89 21.18±1.60中回20.32±0.96 19.94±1.26 19.99±0.96 20.26±0.53 20.29±1.92底回22.08±1.72 12.09±0.85*13.68±1.12#14.07±1.76#19.85±1.94##外毛细胞顶回74.30±4.73 71.58±6.25 71.96±5.97 72.87±7.03 72.98±2.96中回72.18±5.39 68.84±3.85 69.92±4.86 71.62±6.02 72.05±2.85底回78.18±3.82 48.09±8.89**54.96±6.32#59.03±5.80#73.26±6.37##
2.5 螺旋神经节细胞中Nrf2和HO-1的表达 Nrf2、HO-1 蛋白在对照组螺旋神经节细胞的胞浆中呈棕褐色表达。与对照组相比,模型组螺旋神经节细胞胞浆中Nrf2、HO-1 蛋白表达显著降低;与模型组比较,GBE 各组Nrf2、HO-1 蛋白表达明显上调,并表现为剂量依赖性。见图4。
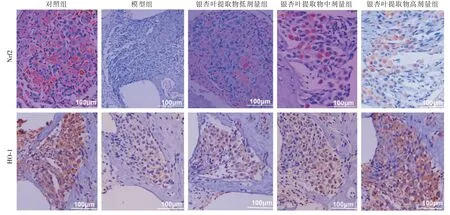
图4 各组螺旋神经节细胞中Nrf2、HO-1 蛋白表达情况(×200)Fig.4 The expression of Nrf2 and HO-1 protein in spiral ganglion cells in each group(×200)
2.6 耳蜗组织中Nrf2 和HO-1 蛋白的表达 与对照组相比,模型组Nrf2、HO-1 的表达水平均显著降低(P<0.01),与模型组对比,GBE 各组Nrf2、HO-1的表达水平剂量依赖性上调,差异具有统计学意义(P<0.05),见图5。
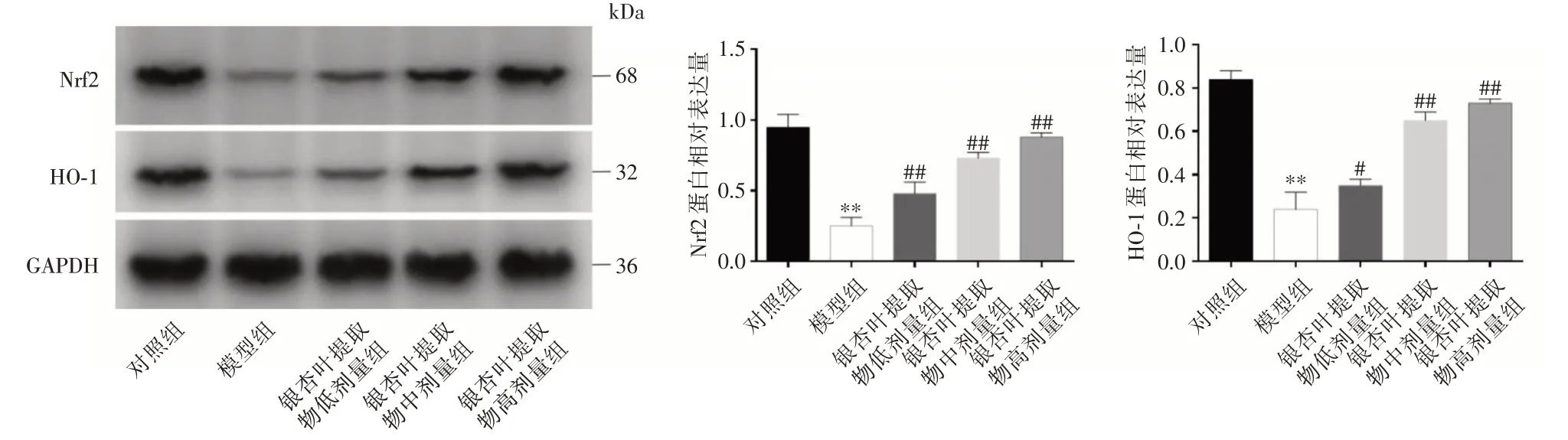
图5 各组大鼠耳蜗中组织Nrf2、HO-1 蛋白表达的情况Fig.5 Expression of Nrf2 and HO-1 protein in cochlea of rats in each group
3 讨论
老年性聋是由环境、衰老、遗传等多种因素共同作用的结果,氧化应激和线粒体功能障碍在耳蜗细胞衰老过程中发挥了重要作用[13]。随着年龄的增长,耳蜗线粒体中内源性抗氧化系统(T-SOD和谷胱甘肽)的活性降低,导致ROS和MDA过表达,进而损害线粒体DNA、双层膜和呼吸链等成分,造成毛细胞和螺旋神经节细胞的氧化损伤[14]。Nrf2可维持线粒体的正常结构和功能,调控线粒体膜电位和氧化磷酸化水平,并促进ATP 合成,进而抑制内耳细胞氧化损伤[15]。HO-1 是Nrf2 的下游产物,Nrf2 移位到细胞核,促进HO-1 转录,从而降低细胞内的ROS、MDA 水平,保护细胞活力[16-17]。LI 等[18]研究发现,Nrf2 和HO-1 蛋白在衰老大鼠听皮层中的表达明显下调,听功能显著减退,其原因可能是衰老阻断了Nrf2 的表达,抑制HO-1 转录,进而使抗氧化防御系统减弱,线粒体氧化损伤增强,从而导致线粒体功能障碍和ROS 过表达。由此可见,减轻机体氧化应激反应对延缓老年性聋至关重要。GBE 可通过清除ROS、提高线粒体功能对耳聋、心脑血管等疾病发挥保护作用[19]。CHEN 等[20]研究表明,GBE 可激活Nrf2∕HO-1 通路,抑制线粒体氧化损伤及凋亡,从而保护心肌细胞。然而,既往GBE 防治老年性聋的研究大多集中于临床观察,未曾对机制进行深入探讨,鉴于此,本研究通过构建老年性聋模型,来研究GBE 是否能通过Nrf2∕HO-1 信号通路减轻氧化应激对老年性聋发挥保护作用。
既往研究采用GBE 联合巴曲酶治疗老年性聋患者,治疗后老年性聋患者的纯音听阈阈值明显降低,血清中T-SOD 水平显著提高,NO 水平明显降低,耳蜗细胞氧化应激损伤减轻,听功能改善[21]。本研究与既往研究具有相似性,本研究的结果显示,模型组大鼠ABR 阈值明显升高,螺旋神经节细胞超微结构明显受损,内、外毛细胞形态紊乱,数量大量丢失,血清中T-SOD 含量降低、MDA含量升高,Nrf2 和HO-1 蛋白表达明显下调。而经不同剂量GBE 治疗后,ABR 阈值显著降低,螺旋神经节细胞的超微结构明显改善,内、外毛细胞形态清晰,数量呈剂量依赖性增多,血清中T-SOD 含量升高,MDA 含量降低,Nrf2 和HO-1 蛋白表达均显著上调。
本研究应用耳蜗基底膜铺片,Myosin-Ⅶa 标记观察毛细胞形态和数量变化,透射电镜观察螺旋神经节细胞超微结构,并用免疫组化与免疫印迹法同时检测出GBE 对各组大鼠Nrf2∕HO-1 信号通路影响,具有一定的创新性。虽然本研究对GBE治疗老年性聋进行了深入探讨,但导致老年性聋的病因和发病机制相对复杂,而本研究仅进行了体内实验,未进行体外实验,也未对相关靶点基因进行验证,今后还需要从全方位、多层次、多角度结合更加科学、前沿的技术来阐明GBE 防护老年性聋的机制,为临床上探索老年性聋的靶向治疗药物提供帮助与依据。
综上所述,GBE 可延缓老年性聋的进展,其机制可能是通过Nrf2∕HO-1 信号通路提高了线粒体的功能,减轻了氧化应激对耳蜗细胞的损伤。
猜你喜欢
介入放射学杂志(2022年8期)2022-11-26
中草药(2022年17期)2022-09-05
中国临床医学影像杂志(2022年5期)2022-07-26
中国听力语言康复科学杂志(2021年6期)2021-12-21
健康之家(2021年19期)2021-05-23
中华耳科学杂志(2021年1期)2021-02-01
听力学及言语疾病杂志(2020年4期)2020-12-20
复旦学报(医学版)(2020年2期)2020-04-17
发明与创新·中学生(2019年3期)2019-03-20
浙江大学学报(理学版)(2019年1期)2019-02-27
