
Imbalance between nitrogen and potassium fertilization influences potassium deficiency symptoms in winter oilseed rape(Brassica napus L.)leaves
2022-03-30 08:52JingLiWenshiHuZhifengLuFanjinMengRihuanCongXiaokunLiTaoRenJianweiLu
The Crop Journal 2022年2期
Jing Li, Wenshi Hu, Zhifeng Lu, Fanjin Meng, Rihuan Cong, Xiaokun Li, Tao Ren,*,Jianwei Lu
a College of Resources and Environment, Huazhong Agricultural University, Wuhan 430070, Hubei, China
b Key Laboratory of Arable Land Conservation (Middle and Lower Reaches of Yangtze River), Ministry of Agriculture and Rural Affairs, Wuhan 430070, Hubei, China
Keywords:Potassium deficiency Nutrient equilibrium Dilution effect Critical K concentrations Photosynthetic characteristics
ABSTRACT Chlorosis at leaf margins is a typical symptom of potassium(K)deficiency,but inappropriate application of K with other nutrients often masks symptoms of K deficiency. A two-year field experiment was conducted to measure the interactive effects of N and K on leaf photosynthesis and dry matter accumulation and the resulting growth dilution effect on K concentration and leaf K deficiency symptoms.N application aggravated the imbalance of N and K nutrients and further exacerbated K deficiency symptoms under K limitation.Synergistic effects of N and K promoted plant growth,amplified the growth dilution effect,and reduced the critical K concentration in leaves. Using 90% of the maximum shoot biomass as a threshold,the critical K concentration was 0.72%at the recommended N(N180)fertilization level.The critical K concentration increased by 62.5% owing to the reduced biomass under insufficient N (N90) supply. In contrast, high N (N270) reduced the critical K concentration (0.64%), accelerating chlorophyll decomposition and exacerbating K deficiency symptoms. The basis of changing the critical K concentration by magnifying growth dilution effect was the functional synergistic effect of N and K on photosynthetic characteristics. Under insufficient N, the low maximum carboxylation rate (Vcmax) limited the net photosynthetic rate(An)and necessitated more K to maintain high CO2 transmission capacity,to improve the total conductance gtot /Vcmax ratio. High N supply increased gtot and Vcmax, possibly mitigating the effect of K reduction on photosynthesis. In conclusion, it is unwise to judge K status of plants only by K concentration without accounting for crop mass (or dilution effect), critical K concentration and deficiency symptoms are affected by N fertilization, and the synergistic effect of N and K on leaf photosynthesis is the foundation of maximal growth of plants under diverse critical K concentrations.
1. Introduction
Potassium(K),which is the most abundant cation in plant cells and accounts for up to 10% of plant dry weight [1], is essential for crop growth and yield.It is associated with many of the physiological processes supporting plant growth and development, such as interactions with water,photosynthesis,assimilate transport, protein synthesis, enzyme activation, and electrical charge balance[2,3]. K deficiency increases the generation of active oxygen in leaves and speeds up leaf senescence[4].Chlorosis at leaf margins is a typical symptom of K deficiency, spreading to the leaf center under prolonged K depletion, gradually developing into withering necrosis,and ultimately reducing crop growth and yield[5,6].Crop deficiency symptoms are indicators of crop nutrient status, which in turn informs strategies to ensure adequate fertilization. However,K deficiency symptoms may not be visible in the field because of rapid redistribution of K within the plant [7]. Moreover, unbalanced application of K relative to other nutrients(especially under low-N conditions) may mask the symptoms of K deficiency [8].Thus,appropriate K fertilization is frequently neglected in practice.
In addition to leaf K concentration, the severity of K deficiency symptoms is affected by leaf growth and the concentrations of other nutrients [9,10]. Nitrogen (N) is the main limiting nutrient of crop yield in current agricultural production [11], and inappropriate application of N fertilizer is common [12]. Imbalance between N and K is widespread [13]. Previous study [14] have shown that an excess of N and concomitant shortage of K disturbs canopy growth, limiting plant growth. In contrast, K deficiency under low N supply exerts only a minor negative effect on rice growth in comparison with excessive N treatment[15].The effects of K deficiency on plant growth under different N supply conditions may be closely linked to changes in leaf K concentration.Rui et al. [16] reported that K concentration was lower with than without N fertilizer treatment.Similarly,in rice,increased N fertilization increased leaf N concentration while reducing leaf K concentration [15,17]. The reduction of leaf K concentration achieved by application of N fertilizer may be due to a dilution effect that promotes plant growth[18].Increasing plant growth by N fertilization is also associated with improved photosynthesis, given that 90% of the carbohydrates that support plant growth are derived via fixation of photosynthetic carbon dioxide(CO2)[19].Photosynthesis depends not only on CO2conduction and assimilation, but also on the coordination between CO2conduction and assimilation,which are critical for maximizing photosynthetic capacity [20].Previous studies [15,21] have shown that N and K nutrition has synergistic and complementary effects on CO2conduction and assimilation. Consequently, understanding phenotypic differences and the physiological mechanisms of K deficiency under different N fertilization regimes will assist in determining plant K nutrition status and guide the application of N and K fertilizers.
Oilseed rape(Brassica napus L.)is an important source for edible oil and energy materials, which widely grown throughout the world [22]. Oilseed rape has substantial N and K requirements(200–300 kg N ha-1and 292–375 kg K2O ha-1,respectively),which are likewise higher than those of cereal crops. Any imbalance between N and K fertilization severely restricts rapeseed yield potential [23,24]. In production, farmers tend to pay more attention to the application of N than to that of K fertilizer [25]. Typically, a large amount of K is removed with the aboveground part of the crop,and underapplication of K fertilizer exacerbates K deficiency [26]. In field experiments, we observed that K deficiency symptoms in oilseed rape leaves are more severe under high than low N fertilizer application. Excessive application of N fertilizer exacerbates the symptoms of K deficiency, although insufficient application of N fertilizer may mask these symptoms in crops.The differences of K deficiency symptoms under different N rates were stable and repeated in positioning experiment from 2016.The unbalanced application of N and K interferes with diagnosis of K deficiency in leaves and fertilization. However, the reason for differences in symptoms of K deficiency and its effect on leaf photosynthetic characteristics and plant growth under different N fertilization levels remains unknown.
Symptoms of K deficiency in leaves are closely associated with the nutritional status of K. K concentration shows an allometric relationship with increase in crop biomass and is affected by the amount of N fertilizer. The aim of this study was to determine whether the aggravation of K deficiency after increasing N fertilization was due to the synergistic regulation of N and K nutrition on photosynthesis, promoting crop mass and increasing growth dilution effect while reducing K concentration.We tested the following hypotheses: (i) N fertilization amplifies the effects of K on plant growth, reduces K concentration due to growth dilution effect,and aggravates leaf K deficiency symptoms;(ii)plants can alleviate K limitation by reducing K concentration after N fertilization; (iii)the basis of K and N concentration in plants linked by allometric relationship with crop mass involves the coordinated regulation of leaf photosynthesis by N and K nutrition.
2. Materials and methods
2.1. Experimental site
The field experiment was conducted in Wuxue county, Hubei province, central China (30°06′N, 115°36′E), during the oilseed rape growing season from 2017 to 2019.The soil at the experimental sites is sandy loam,classified as an Ultisol[27].The soil with the following characteristics in the topsoil layer (0–20 cm): pH 5.76,organic matter 32.1 g kg-1, total N 1.75 g kg-1, exchangeable K 54.5 mg kg-1, non-exchangeable K 296.5 mg kg-1, Olsen-P 13.4 mg kg-1, hot-water soluble B 0.48 mg kg-1and the cation exchange capacity (CEC) 4.2 cmol kg-1. Low soil nonexchangeable K content and low CEC indicated that the soil had low K fertility status as well as low buffering ability and fertilizer retention capacity. Total precipitation and effective cumulative temperature during the winter oilseed rape growing seasons were 3164 and 3091 mm and 975 and 911 °C, respectively.
2.2. Experimental design and management
A two-factor, randomized complete block design with three replicates was adopted for the field experiment.The combinations of N and K fertilizer followed Ren’s [28] recommendation of optimal fertilization for high seed yield in winter oilseed rape production. There were nine experimental treatments, based on combinations of N fertilizer application rate (insufficient N: N90,90 kg N ha-1;moderate[recommended rate]N:N180,180 kg N ha-1;or high N:N270,270 kg N ha-1)and K fertilizer application rate(no K:K0,0 kg K2O ha-1;low K:K60,60 kg K2O ha-1;or moderate[recommended rate] K: K120, 120 kg K2O ha-1). Each plot was 20 m2(length 5 m and width 4 m).
To ensure that other nutrients did not limit plant growth,90 kg P2O5ha-1and 9 kg B ha-1was applied to all treatments.Fertilizers were urea(46%N),calcium superphosphate(12%P2O5),potassium chloride(60%K2O),and borax(10.8%B).Basal fertilizers,including 60% of the N fertilizer and all of the P, K, and B fertilizers, were incorporated 1 day before transplanting.The remaining N fertilizer was applied at 20%during the overwintering stage(BBCH stage 19)[29] and at 20% on initiation of the stem elongation stage (BBCH stage 30).
The winter oilseed rape cultivar used was Huayouza No. 9,which is grown widely in the Yangtze River Basin. Seeds were sown in seedbeds in mid-September. After 35 days, uniform seedlings with 4–5 leaves were transplanted into the field by hand, in double rows spaced approximately 0.3 m apart and with a spacing between plants of 0.25 m(112,500 plants per hectare).Field management,including herbicide application and pest and disease control,followed local practices.There were no obvious weed,pest,or disease problems during the two growing seasons.
2.3. Sampling and measurement
Owing to the strong demand for N and K nutrients during stem elongation of oilseed rape, insufficient K supply may cause symptoms of K deficiency in lower leaves [30]. Differences in K deficiency symptoms among different N fertilizer application rates in lower leaves were evaluated during stem elongation. During the stem elongation stage (BBCH stage 32–33) on March 12, 2018,and March 10, 2019, at least five uniform plants were sampled from each plot. The mean number of leaves was 13, with the number varying from 11 in the N90K0treatment to 15 in the N270K120treatment.Leaf growth status during the stem elongation stage in the 2018–2019 winter growing seasons are shown in Fig. S1.
2.3.1. Gas exchange and An-Cicurve measurement
Tagged leaves were subjected to simultaneous leaf gas exchange and CO2response curve measurements using a portable open-circuit infrared gas analysis system (Li-6400XT; Li-Cor, Inc.,Lincoln, NE, USA). Measurements were performed on at least five randomly selected leaves in each treatment on a sunny morning(09:00–15:00)under a light-saturating photosynthetic photon flux density (PPFD) of 1200 μmol m-2s-1(90% red light and 10% blue light). The CO2concentration in the leaf chamber (Ca) was set at 400 μmol mol-1air. The leaf temperature was maintained at 20 ± 0.2 ℃, the relative humidity was controlled at 50%–60%, and the flow rate was 500 μmol s-1. After acclimation to steady-state photosynthesis for 10 min, the net photosynthetic rate (An), stomatal conductance (gs), and intercellular CO2concentration (Ci) was recorded. The photosynthesis response to varying substantial CO2concentrations (An–Cicurves) was then measured in three leaves under the same environmental conditions. Cawas reduced stepwise from 400 to 300, 200, 150, 100, and 50 μmol CO2mol-1,and then increased from 50 to 400, 600, 800, 1000, 1200, and 1500 μmol CO2mol-1. The maximum carboxylation rate (Vcmax)and maximum electron transport rate (Jmax) were then calculated following Farquhar et al. [31]:
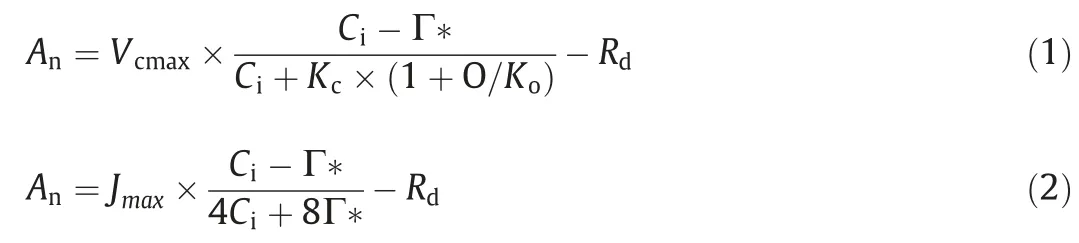
where Anand Ciare gas exchange measures and Γ* is the chloroplastic CO2compensation point in the absence of mitochondrial respiration: a typical value, 42 μmol mol-1, was used [32]. Kcand Ko,the Michaelis–Menten constants for carboxylation and oxygenation,were 406.7 μmol mol-1and 277.1 mmol mol-1,respectively[32].O is the partial pressure of CO2(210,000 μmol mol-1).Rd,the day respiration rate, was measured following Brooks and Farquhar [33].gtot, the total conductance to CO2diffusion from the leaf surface to carboxylation sites, was determined by Eq. (3):

where gscis the stomatal conductance to CO2calculated from the stomatal conductance to water, which is equal to gs/1.6 [34].
2.3.2. Estimation of gmby curve fitting
The gmestimation method followed Ethier and Livingston [35].The non-rectangular hyperbola version of Farquhar’s [31] biochemical model was used to fit the An-Cicurves.The model is based on the assumption that the curvature of the An-Cicurve decreases because of a finite value of gm. Values of the Michaelis–Menten constant for CO2(Kc) and oxygen (Ko) were obtained from Bernacchi et al.[36].Γ*was set at 42 μmol mol-1,a typical value,and Rdwas estimated as described previously.Because the gmobtained by the curve-fitting method was tightly coupled with the gmestimated from gas exchange and chlorophyll fluorescence measurements, the gmestimates from the An-Cicurves were used in the analysis.
2.3.3. Determination of leaf characteristics
After the photosynthetic parameters were measured, all leaves were removed and photographed with a digital camera, and the leaf area(S)was then estimated using Image-Pro Plus 6.0 software[37]. Using the color-recognition function of the software, the yellow areas(YS)of leaves were identified.The proportion of chlorotic area (PC) was then calculated as follows:

Leaf chlorophyll content was measured following Wintermans and DeMots[38]. Fresh leaf disks (0.25 g) were extracted by soaking for 48 h in 25 mL of 96%ethanol.Chlorophyll a and Chlorophyll b content were measured at 665 and 649 nm, respectively, with a spectrophotometer (UV2102 PCS, Unico, Shanghai China). The remaining leaves were dried at 105 °C for 30 min to deactivate enzymes, and then at 65 °C to constant weight for determination of plant biomass and N and K concentrations. The dried samples were ground into powder and digested with H2SO4–H2O2[39],and total N was measured with a flow injection auto-analyzer(AA3; Seal Analytical, Inc., Southampton, UK). Flame photometry(M-410; Cole-Parmer, Chicago, IL, USA) was adopted to determine the total K content.Shoot N and K uptake were determined by multiplying the dry matter weight by the respective concentrations in aboveground parts in the stem elongation stage.
2.4. Calculations and statistical analysis
2.4.1. Calculation of parameters
To construct nutrient vector diagrams,plant tissue N and K concentrations(%of dry mass)were multiplied by the mean dry mass to yield nutrient contents(g) for each treatment.Relative nutrient concentration,nutrient content,and unit dry mass were calculated as:

where R represents the value relative to the control treatment. For measuring the effect of N application on K dilution,the low-N(N90)treatments under different K application rates were defined as the control to calculate the relative R values. Data points (x, y, and z)were plotted in a relative nutrient content(x),relative nutrient concentration(y),and relative unit dry weight(z)space.Responses are depicted by vectors whose length represents magnitude of response and direction identifies specific nutritional responses. For example,if N fertilizer application increases the uptake of a specific nutrient(content), but reduces nutrient concentration, then dilution of that nutrient is indicated, given that nutrient concentration in plants is linked by an allometric relationship with growth. If a similar response occurs, but without changing the nutrient concentration,then nutrients are likely to be sufficiently provisioned. Responses indicating alleviation of nutrient limitation occur when increased nutrient concentration increases biomass growth, resulting in increased nutrient content. Further interpretations of vector nutrient diagrams can be found in Isaac and Kimaro [40].
With reference to the method for determining critical soil nutrient content based on yield responses[41],the critical K concentration influencing shoot biomass was determined from the relationship between leaf K concentration and shoot biomass in lower leaves, using the quadratic-platform model 90% of the maximum biomass was taken to indicate a strong effect on plant growth. Under all nitrogen rates, the optimal K concentration in leaves was defined as the minimum K concentration needed to obtain the maximum biomass.
2.4.2. Statistical analysis
Descriptive statistical analyses were used to obtain range and standard error (SE) statistics for the variables. All data were subjected to two-way analysis of variance performed using SPSS 18.0(SPSS Inc., Chicago, IL, USA) to identify statistically significant differences in the measured traits(Tables 1 and 2)among the various N and K treatments;N×K interaction effects were also estimated.Differences in traits among the K treatments under a single N treatment were tested using the least significant difference (LSD) post hoc test.Graphs were created,and regression analyses performed,with OriginPro 8.5 software (OriginLab, Inc., Northampton, MA,USA).

?

?
3. Results
3.1. Shoot biomass and leaf characteristics
The effects of N and K fertilization on shoot biomass, leaf area and the proportion of chlorotic area are shown in Fig.1.Compared with the N90treatment, shoot biomass increased respectively by 38.8% and 42.4% and by 40.5% and 48.1% on average in the N180and N270treatments during 2017–2018 and 2018–2019 winter oilseed rape growing seasons. Corresponding, the average leaf area were 36.7%and 49.6%higher in the N180and N270treatments than in the N90treatment during two consecutive years’experiments.K fertilizer application increased the positive effects of N fertilization on shoot biomass and leaf area.However,the effects of K fertilization differed with N supply. Under low N supply (N90), K fertilization showed minor effects on shoot biomass and leaf area,whereas it markedly increased shoot biomass and leaf area by respectively 52.9%–89.8% and 24.9%–62.6% under moderate (N180) and 92.2%–147.3%and 60.1%–111.9%under high N(N270)supply in two years.In contrast,under K limitation,the proportion of chlorotic area was increased with increased N fertilization. Compared with moderate N (N180) treatment, the proportion of chlorotic area decreased by 26.1% and 34.5% in the insufficient N (N90) treatment during 2017–2018 and 2018–2019. In contrast, the high N (N270) treatment increased the proportion of chlorotic area by as much as 62.4% and 95.2% relative to the N180treatment during 2017–2018 and 2018–2019 winter oilseed rape growing seasons.
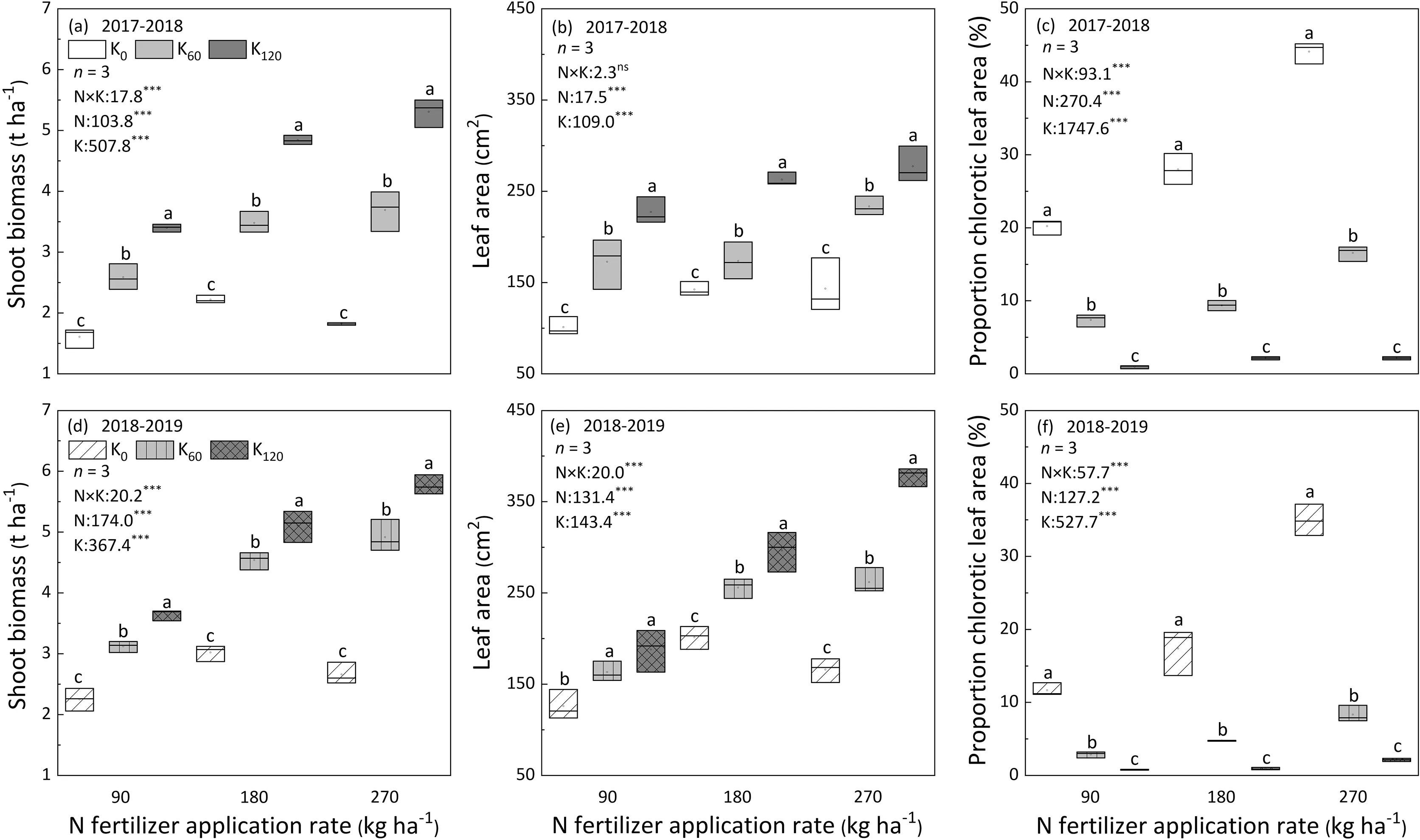
Fig.1. Shoot biomass,leaf area, and proportion of chlorotic area during stem-elongation stage under the combined application of N and K fertilization treatments in 2017–2018 and 2018–2019 winter oilseed rape growing seasons in Wuxue county,central China.Bars superscripted with different letters under the same N treatment in the same year were significantly different between different K fertilization rates by LSD test(P <0.05).N,Nitrogen;K,Potassium;N×K,Interaction between N and K; ***,P ≤0.001, ns means no significant effect.
Similar to shoot biomass, the shoot N accumulation of oilseed rape showed a significant response to K fertilization at the stem elongation stage (Fig. 2). In comparison with the K0treatment,shoot N accumulation was increased by a mean of 51.9% after K application.Similar responses of K accumulation to N supply were observed. In sufficient-N treatments (N180and N270), the mean K accumulation was 31.9% higher than in the N90treatment.
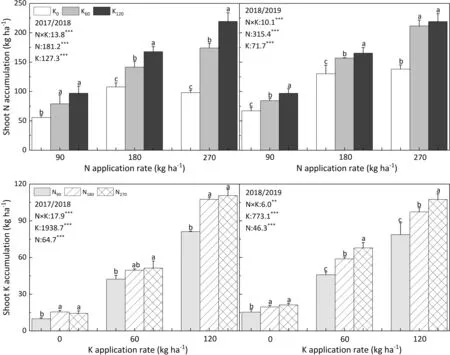
Fig. 2. Shoot N and K accumulation under different N and K fertilization treatments at stem-elongation stage during 2017–2018 and 2018–2019 growing season. Bars superscripted with different lowercase letters under the same N treatment in the same year are significantly different between different K fertilizer treatments by LSD test(P <0.05). N, Nitrogen; K, Potassium; N×K, Interaction between N and K; **, P <0.01; ***, P ≤0.001.
3.2. Leaf nutrients and chlorophyll content
Leaf N and K concentration were significantly influenced by the application of N and K fertilizer(Table 1).In the N90treatment,the mean N concentration was 17.5%lower than in the N180treatment,whereas the leaf K concentration was increased by a mean of 46.8%. In contrast, high N (N270) application increased leaf N concentration and promoted plant growth; however, it reduced leaf K concentration.In comparison with the N180treatment,N concentration was increased by 17.0%in the high N treatment,but the leaf K concentration was 11.7% lower. Leaf N to K ratio, used to measure the balance of leaf N and K nutrients, increased significantly when the application of N and K fertilizers was unbalanced(uncoordinated input of N and K fertilizers), and the ratio of N to K decreased rapidly after K fertilization. The total chlorophyll content (Chlorophyll a + Chlorophyll b) of leaves was also influenced by the combined application of N and K fertilizers. In comparison with the low N (N90) treatment, the chlorophyll content in the N180and N270treatment increased by only 9.3%and 12.5%on average. In sharp contrast, the chlorophyll content increased by as much as 44.5%after K fertilization, and with the increase of N fertilization, the effect of K on the chlorophyll content increased. In contrast to chlorophyll content, the ratio of chlorophyll a content to b content (Chlorophyll a/b) was only slightly affected by N and K fertilization. The value of Chlorophyll a/b increased by only 7.7% on average after K application (Table 1).
Fig. 3 shows the relationship between chlorophyll and leaf N and K concentration. There was a quadratic relationship between chlorophyll and leaf N and K concentration, which was negatively correlated with leaf N-to-K ratio. In contrast, under the condition of unbalanced N and K fertilization,the proportion of chlorotic area was a quadratic function of N concentration,but decreased rapidly with the increase in leaf K concentration. There was a significant positive correlation between the proportion of chlorotic area and leaf N-to-K ratio.
3.3. Plant nutritional responses to biomass growth
Leaf N concentration decreased with the increase of biomass under all N application rates after K fertilization (Fig. S2a). There was a significant negative correlation between N and K concentration (Fig. S2b). The interaction between N and K was further characterized by vector analysis (Fig. 4). K application promoted plant growth and diluted N concentration, the dilution intensity was amplified with increased K fertilizer application (Fig. 4a). Combined application of N promoted plant growth and reduced K concentration,while in the K0treatment,N fertilization aggravated the demand for K, resulting in more severe K limitation (Fig. 4b).
Based on the dilution effect of N fertilization on K concentration, differences in critical K concentration in leaves under different N application rates were measured (Fig. 5). Taking 90% of the maximum shoot biomass to be the threshold of winter oilseed rape under different N fertilization treatments at the stem elongation stage, the critical K concentrations in the N90, N180, and N270treatments were 1.17%, 0.77%, and 0.64%, respectively. Compared with the low N (N90) treatment, the critical K concentration of the moderate N (N180) and the high N treatments decreased by 34.2% and 45.3%, respectively. Like the critical K concentration of leaves, the optimum K concentration decreased with increased N fertilization.

Fig. 3. Curves fitting chlorophyll content, proportion of chlorotic area and leaf N, K concentration, and leaf N-to K-ratio in 2017–2018 and 2018–2019 winter oilseed rape growing seasons. ***, P ≤0.001.

Fig. 5. Leaf critical K concentration under different N treatments. The data were fitted by the corresponding optimum equation (P <0.01).
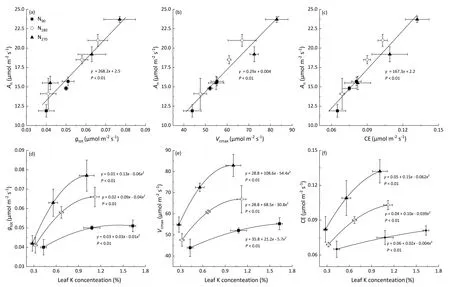
Fig. 6. Correlations of net photosynthetic rates (An) with (a) CO2 total conductance (gtot), (b) the maximum carboxylation rate (Vcmax), (c) carboxylation efficiency(CE), and the effects of leaf K concentration on (d) CO2 total conductance (gtot), (e) the maximum carboxylation rate (Vcmax), and (f) CE under different N fertilizer rates.
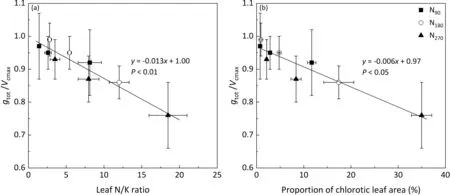
Fig. 7. Effects of leaf N/K ratio on (a) gtot /Vcmax, and the correlations of the proportion of chlorotic leaf area with gtot /Vcmax (b).
3.4. Photosynthetic characteristics
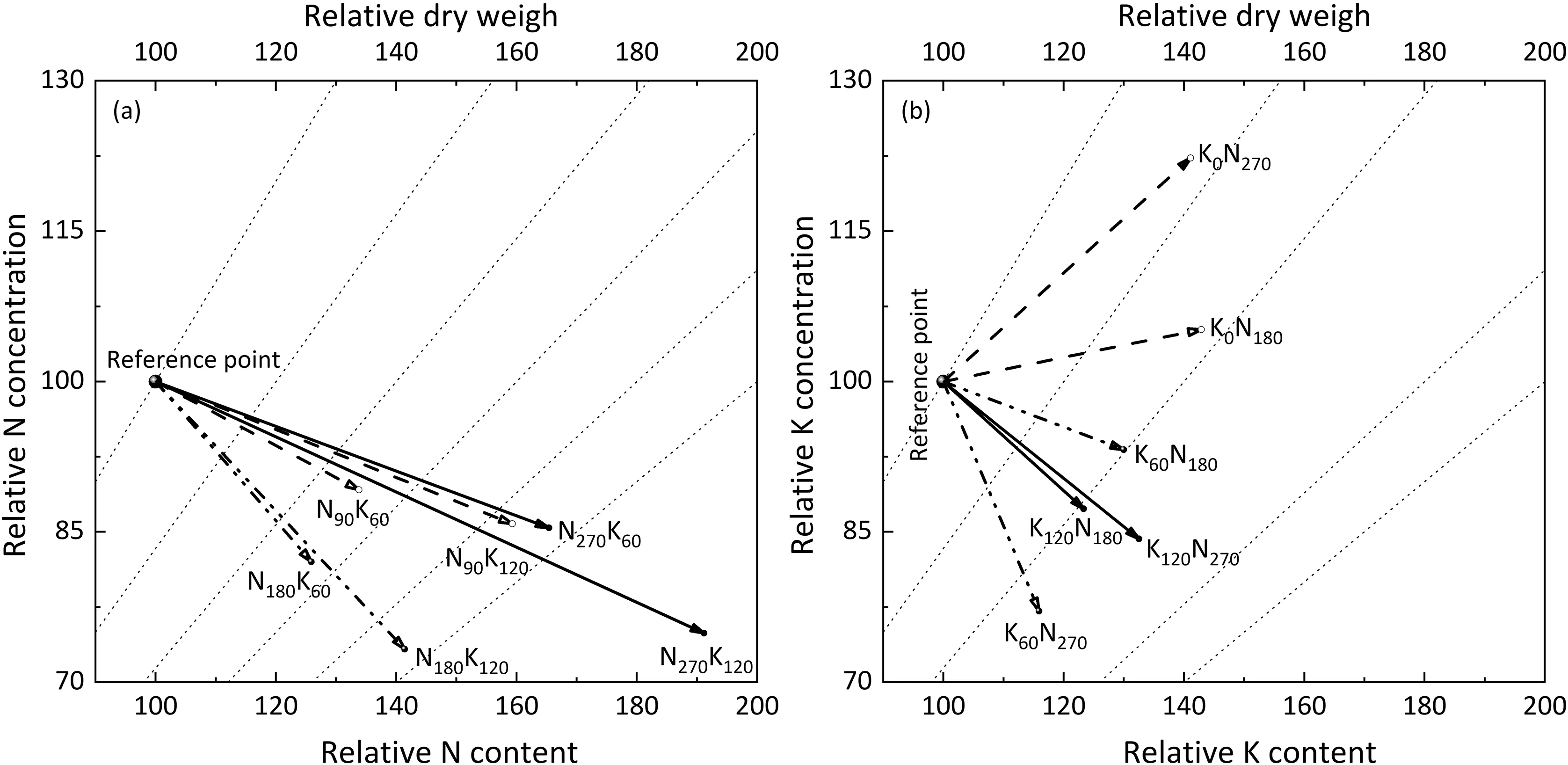
Fig.4. Vector diagrams of mutual effects of N and K under combined application of N and K fertilization.Changes in nutrient content(bottom horizontal x-axis)are expressed as nutrient concentration(vertical y-axis)multiplied by dry biomass(top horizontal z-axis extending to the second vertical y-axis).N nutrition responses to K under several N application rates are expressed relative to N accumulation(Na)without K application following:(Na with K/Na without K control)×100.Gray dashed lines indicate the z-axis responses by referencing to the zero-point, no-effect, intersection of the z- and y-axis. Dashed lines indicate biomass responses in intervals of ± 25% that of the controls.
N and K fertilization increased photosynthetic parameters(Table 2). In comparison with the N90treatment, Anvalues were increased by 26.3% and 37.6% in the N180and N270treatments.Application of K fertilizer increased the positive effect of N fertilization on An. Under low N supply, K fertilization showed minor effects on An, but markedly increased Anby 38.8% to 40.3% under moderate and high N supplies. Increase in Anwas closely related to CO2conductance capacity. Higher N supply increased CO2conductance. In sufficient N fertilization (N180and N270) treatments,the average gs, gm, and gtotvalues increased by respectively 5.1,29.6, and 20.6%, in comparison with the N90treatment. The influence of N supply on CO2conductance varied with K supply.In comparison with the K0treatment,the gs,gm,and gtotvalues under the K60and K120treatments were increased by respectively 42.8%,39.1%, and 38.9%, and 104.8%, 46.0%, and 60.6%.
Both Vcmaxand CE values increased with increased N and K supply (Table 2). In comparison with the insufficient N (N90) treatment, the average Vcmaxand CE values increased by respectively 15.9% and 17.6% for the N180and 39.0% and 45.1% for N270treatment. In the K60and K120treatments, the mean values of Vcmaxand CE were respectively 26.6% and 26.7%, and 40.1 and 45.8%,higher than in the K0treatment. The CO2assimilation-increasing effect of N increased gradually with increased K fertilizer application.Similar responses of Jmaxto increase N and K supply were also observed.Using the gtot/Vcmaxratio to estimate the degree of coordination between CO2conductance and carboxylation, under the same amount of K fertilization, the increase of Vcmaxwith N fertilizer application was greater than that of gtot; this reduced the degree of coordination between CO2conduction and carboxylation.In contrast, K fertilization substantially increased the degree of coordination between CO2conduction and carboxylation under the same N fertilizer application.
3.5.Effects of leaf K concentration on photosynthetic parameters under N fertilization treatments
Leaf photosynthetic capacity was affected mainly by CO2conductance and carboxylation. There were significant positive linear relationships of Anwith gtot,Vcmax,and CE.The effects of leaf K concentration on gtot, Vcmax, and CE under different N fertilization treatments are shown in Fig. 6. In the N180treatment, the rates of change in gtot, Vcmax, and CE with K concentration per unit leaf area were respectively 75.0%, 81.5%, and 89.7% higher than those in the N90treatment but 50.0%,76.6%,and 59.0%lower in comparison to the high N (N270) treatment. The gtot, Vcmax, and CE values were decreased by respectively 15.1%, 15.0%, and 17.5% in the insufficient N (N90) treatment compared with the recommended N (N180) treatment. In contrast, excessive N (N270) input increased gtot, Vcmax, and CE by respectively 12.8%, 22.2%, and 24.6% in comparison with the N180treatment. The degree of coordination between CO2conductance and carboxylation was affected by the balance between N and K in leaves, while the gtot/Vcmaxand N/K ratios showed a clear negative linear correlation (Fig. 7a). The gtot/Vcmaxratio decreased linearly with increasing proportion of leaf chlorotic area (Fig. 7b).
4. Discussion
4.1.Effects of chlorosis at leaf margins caused by K deficiency on plant growth
As the main organ of plant carbon assimilation, K deficiency accelerated chlorophyll degradation and caused leaf chlorotic,reduced leaf photosynthetic rate and limited the accumulation of plant dry matter (Figs. S3 and S4). Chlorophyll is responsible for light harvesting in photosynthesis, resulting in the excitation of electrons that are used to drive the NADPH and chemical energy in the form of ATP, for the reactions of the Calvin–Benson cycle[42]. Furthermore, K-deficient leaves accumulated soluble sugar and reduced the rate of assimilate export before photosynthesis was reduced [43]. Carbohydrate accumulation in chloroplast may destroy chloroplast structure, aggravating chlorophyll decomposition and leaf chlorotic. Synergistic improvement in leaf photosynthetic area and rate is essential for enhancing plant growth.Previous studies [44,45] shown that K deficiency accelerated leaf senescence, reduced leaf area, and delayed leaf appearance rate,which markedly reduced the interception of photosynthetically active radiation(PAR).In the present study,with increased N application rate, the limitation by K deficiency of leaf area expansion gradually increased, and chlorosis at leaf margins was increased,further reducing green leaf area (effective photosynthetic area).Consequently, the combined application of N and K was essential for increasing photosynthetic area and rate.
Responses of chlorosis at leaf margins to K insufficiency differed by N fertilization rate. N fertilization exacerbated chlorosis at leaf margins under K limitation, possibly in association with leaf N/K ratio. Large variations in N and K concentrations influence leaf physiological characteristics and plant growth; under a high N/K ratio, protein synthesis is blocked because low K concentrations result in the accumulation of free amino acids and break the charges balance on proteins[46].Low K also triggers the synthesis of putrescine and may accelerate chlorosis at leaf margins [47].These changes damage mesophyll cell homeostasis and reduce photosynthetic capacity[48,49]. A high leaf N/K ratio also reduces N metabolism and N-assimilation enzyme activity and influences leaf growth [50]. N fertilization increases leaf chlorophyll content and light energy capture [51]; however, the excess energy may induce more reactive oxygen species,destroy chloroplast structure,expedite chlorophyll decomposition, and cause the leaf to yellow in the absence of K[52].In the present study,there was a quadratic relationship between chlorophyll and leaf N, K concentration,which was negatively correlated with leaf N to K ratio.In addition,with the imbalance between N and K fertilization, the chlorosis at leaf margins deteriorated with increased leaf N concentration,while decreasing rapidly with increased K.
4.2. Growth dilution effect
The dilution of nutrient concentration resulting from the accumulation of dry matter is common [53,54]. This mechanism has frequently been suggested to account for reduced N concentrations in the process of plant growth [55]. However, changes in nutrient concentration are associated not only with plant growth stage and the availability of the nutrient, but with interactions among nutrient elements.According to Lemaire et al.[53],the availability of a given element to plants is in large part determined by the availability of other elements. In the present study, N and K fertilization showed mutually positive effects on plant growth and nutrient accumulation. This finding is consistent with the views in the recent review of Briat et al. [54], who pointed out that P and K uptake dynamics are determined not only by the quantity of aboveground biomass accumulated by the crop, but by the N nutrition status of the crop as influenced by N fertilizer supply.Under all N application rates, with increasing K fertilizer, leaf N concentration decreased with the increase of shoot biomass. Vector analysis also showed a mutual dilution effect between N and K, while without K application, the increase of N fertilization exacerbated the lack of K. Ziadi et al. [55] showed that high N fertilization on wheat or maize crops under a limiting P soil supply exacerbated P deficiency. Critical K curves depend highly on crop N supply, and the critical concentration of the major nutrients(N,P,and K)decreases as plant mass increases[54].Measurement of leaf critical K concentration under several N application rates showed that in comparison with the low N (N90) treatment, the critical K concentrations of the moderate N (N180) and high N(N270) treatments decreased by 34.2% and 45.3%, respectively. N fertilization promoted the growth dilution effect of plants, reducing the concentration of K in leaves [18]. This may be a strategy allowing plants to cope with nutrient imbalance,reduce the threat of limiting nutrients,and ensure maximum growth.Thus,it is dangerous to estimate K status of plants from K concentration alone without reference to crop mass (or dilution effect). And critical K concentration and deficiency symptoms are also affected by N fertilization.
4.3. Functional coordination of K and N
N and K may exert a beneficial synergistic effect on crop growth by regulating leaf photosynthetic carbon gain [15,17,21]. Both K and N are necessary for maintaining cell osmotic pressure and regulating the opening and closing of stomata [56]. Both N and K increase the proportion of mesophyll and chloroplast surface area exposed to intercellular air space per unit leaf area, promoting CO2mesophyll conductance[57,58].During CO2assimilation,high N supply increases the Rubisco enzyme content in leaves while also reducing its activity[59], and optimal K supply also increases the activity of Rubisco [48]. Our results indicate that more K is required to maintain a high photosynthetic rate under low-N conditions, owing to more N was distributed to photosynthetic systems [15,21]. In contrast, sufficient N supply directly increases the total amounts of photosynthetic components, such as Rubisco enzymes, while smaller molecular organic and inorganic N compounds can participate in osmotic pressure stabilisation and stomatal opening regulation, possibly counteracting the effect of a partial decline in K content on photosynthesis [56]. In the present study,synchronous increases in N and K supplementation ensured an appropriate leaf N/K ratio, simultaneously increasing gtotand Vcmaxand thereby optimizing the coordination between CO2conduction and carboxylation [21].
In addition to the coordinated effect of N and K on photosynthetic characteristics, the combined application of N and K may also affect the concentration of other nutrients, such as Mg and Na. As both N and Mg are structural elements of chlorophyll, they exert substantial effects on chlorophyll synthesis [7]. Chlorophyll,N, and Mg concentrations increased significantly with increasing N rates, but declined sharply with increasing K rates owing to the antagonistic relationships between K+and NH4+as well as Mg2+. It is also likely that the increase in K promoted the growth of leaves and diluted Mg concentrations. Previous study [60] have shown that Na can partially replace K in the vacuole, contributing to cell turgor and expansion. N application reduced the K concentration in leaves and may have increased the Na concentration to replace the effect of partial K concentration reduction on plant growth.
4.4. Prospects for application to actual production
In agricultural production, plants may be exposed to multiple simultaneous nutritional stresses (deficiency or imbalance) [52].As essential macronutrients, N and K strongly influence plant growth and yield [7,11]. However, large areas of agricultural land worldwide are deficient in K, including 75% of the paddy soil of China[7]. Additionally, a large amount of K is removed at harvest,accelerating its depletion in soil[61].Consequently,low K fertilizer application in the context of unbalanced fertilization may result in reduction of available soil K reserves,thus diminishing soil fertility[5]. Based on reports by farmers and our field experiments, excessive or insufficient N occurs in about 33% of farm fields in China[12]. In another study [62], excessive and insufficient N affected respectively 20% and 10% of the total sown area in China. Thus,in many areas,inappropriate N fertilization and K deficiency coexist.Insufficient application of N fertilizer masks the symptoms of K deficiency, explaining in part why K deficiency symptoms can be difficult to observe in field trials.In contrast,excessive application of N exacerbates the symptoms of K deficiency and limits plant growth, while an imbalance in the N/K ratio may be a driver of reduced N-use efficiency. Consequently, in production, knowledge of the effects of nutrient interactions,more accurate determination of K nutrition status, and timely supplementation with K fertilizer are necessary to mitigate the negative effects of climate change and abiotic and biotic stress on agriculture.
5. Conclusions
Chlorosis at leaf margins caused by K deficiency was closely associated with the nnutrient balance of N and K in leaves. Imbalance between N and K accelerated the decomposition of chlorophyll. High N fertilizer supply magnified the growth dilution effect, reduced the K concentration in leaves, and intensified chlorosis at leaf margins;The critical concentration of K decreased as plant mass increased after aapplication of N fertilizer.The coordination between N and K stimulates the maximization of net photosynthetic rate under the condition of reduced critical concentration of K resulting from the growth dilution effect. For this reason, K concentration, crop mass (or dilution effect) and the interaction effects of nutrients should be integrated in the evaluation of plant K nutrition status.In short,in crop production,optimizing the ratio of N and K fertilizer will ensure the appropriate ratio of N and K concentration in plants or leaves, reduce leaf chlorosis caused by K deficiency, and coordinate net photosynthetic rate and photosynthetic effective area, thereby increasing photosynthetic efficiency to promote crop growth and achieve higher crop yield.
CRediT authorship contribution statement
Jing Li, Xiaokun Li, Tao Ren, and Jianwei Luconceived and designed the experiments.Jing Li, Wenshi Hu, and Fanjin Mengperformed the experiments.Jing Li,Zhifeng Lu,and Tao Renanalyzed the data.Jing Li and Tao Rendrafted the manuscript.Zhifeng Lu, Rihuan Cong, Tao Ren, and Jianwei Luhelped in revising the manuscript. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (31872173), the National Key Research and Development Program of China (2018YFD0200900),China Agriculture Research System(CARS-12),and the Fundamental Research Funds for the Central Universities (2662018PY077).
Appendix A. Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2021.06.001.

- The Crop Journal的其它文章
- Origin, evolution, and molecular function of DELLA proteins in plants
- Far-red light: A regulator of plant morphology and photosynthetic capacity
- A rice XANTHINE DEHYDROGENASE gene regulates leaf senescence and response to abiotic stresses
- Dissection of heterotic loci for grain yield using interconnected chromosome segment substitution lines in rice
- A soybean NAC homolog contributes to resistance to Phytophthora sojae mediated by dirigent proteins
- The boron transporter SiBOR1 functions in cell wall integrity, cellular homeostasis, and panicle development in foxtail millet