
Origin, evolution, and molecular function of DELLA proteins in plants
2022-03-30 08:50HuidanXueXiangGaoPengHeGuanghuiXiao
The Crop Journal 2022年2期
Huidan Xue, Xiang Gao, Peng He, Guanghui Xiao
a College of Life Sciences, Shaanxi Normal University, Xi’an 710119, Shaanxi, China
b School of Food and Biological Engineering, Shaanxi University of Science and Technology, Xi’an 710021, Shaanxi, China
c School of Ecology and Environment, Northwestern Polytechnical University, Xi’an 710012, Shaanxi, China
Keywords:DELLA proteins GA signaling pathway Cross-talk Plant hormone Growth and development
ABSTRACT Gibberellic acid (GA), a ubiquitous phytohormone, has various effects on regulators of plant growth and development. GAs promote growth by overcoming growth restraint mediated by DELLA proteins(DELLAs). DELLAs, in the GRAS family of plant-specific nuclear proteins, are nuclear transcriptional regulators harboring a unique N-terminal GA perception region for binding the GA receptor GIBBERELLIN INSENSITIVE DWARF1 (GID1) and a C-terminal GRAS domain necessary for GA repression activity via interaction with multiple regulatory proteins. The N-terminal conserved region of DELLAs evolved to form a mode of GID1/DELLA-mediated GA signaling originating in bryophytes and ferns. Binding of GA to GID1 increases the affinity between DELLAs and a SCF E3 ubiquitin–ligase complex, thus promoting the eventual destruction of DELLAs by the 26S proteasome. DELLAs negatively regulate GA response by releasing transcription factors to directly activate downstream genes and indirectly regulate GA biosynthesis genes increasing GA responsiveness and feedback control by promoting GID1 transcription. GA communicates extensively with other plant hormones and uses crosstalk to regulate plant growth and development.In this review,we summarize current understanding of evolutionary DELLA-mediated gibberellin signaling and functional diversification of DELLA, focusing primarily on interactions of DELLAs with diverse phytohormones.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 288
2. Evolutionary DELLA-mediated gibberellin signaling and DELLA genes in plants. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 288
3. Conserved domains of DELLA proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 288
4. Dellas and the GA signal transduction pathway . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290
5. Functions of DELLA proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290
5.1. Crosstalk between DELLAs and other phytohormone signal transduction pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290
5.2. Role of DELLA protein in seed germination. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 291
5.3. Role of DELLA proteins in flowering. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 292
5.4. Role of DELLAs in hypocotyl elongation and apical hook development. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 292
5.5. Role of DELLAs in nitrogen use efficiency (NUE) and yield . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293
5.6. Role of DELLAs in stem elongation, trichome development, and root meristem . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293
5.7. Role of DELLAs in other aspects of plant development. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293
6. Transcriptional regulation and post-translational modifications of DELLA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294
6.1. Transcriptional regulation of DELLA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 294
6.2. Posttranslational modifications of DELLAs. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295
7. Conclusion and perspectives. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295
CRediT authorship contribution statement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296
Declaration of competing interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296
Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 296
1. Introduction
Light intensity, temperature, and humidity are the main environmental factors regulating plant development from seed germination through fruit ripening. Endogenous plant hormones are essential in plant growth and development. Gibberellic acid (GA)is a ubiquitous phytohormone that controls multiple processes of plant growth and development, including germination, stem and hypocotyl elongation, and flowering [1–3]. GAs, a large family of tetracylic diterpenoids discovered by scientists at Tokyo University in the 1950s, have aroused great attention globally owing to their uses in agriculture,nurseries,tissue culture,and other applications[1–3]. To date at least 136 structural variants of GA have been identified in a variety of plants[3,4],with GA1 and GA4 as the main active GAs.GAs stimulate plant growth by overcoming the growthrestraining effects of DELLA proteins (DELLAs), a group of nuclear growth repressors [5]. DELLAs, from the GRAS family of plantspecific nuclear proteins, are key negative regulators of GA signaling [6] and are widely distributed in crop plants including wheat(Triticum aestivum), rice (Oryza sativa), maize (Zea mays), grape(Vitis vinifera), barley (Hordeum vulgare), and soybean (Glycine max) [5,7–9].
Besides DELLAs, the GA receptors GIBBERELLIN INSENSITIVE DWARF1 (GID1) and GID2 and the F-box proteins SLEEY1 (SLY1)and SNEEZY have been shown[10–14]by mutation analysis to participate in GA signaling in both Arabidopsis and rice. GID1 is localized to both the cytoplasm and the nucleus.It binds bioactive GAs to promote formation of the GID1-DELLA complex by interacting with the N-terminal domain of DELLAs [15,16]. DELLAs are subsequently recognized and bound by SCFSLY1/GID2E3 ubiquitin ligase Fbox protein components (SLY1 in Arabidopsis and GID2 in rice),leading to ubiquitination and degradation of DELLAs via the 26S proteasome pathway [11,12,17]. DELLAs participate not only in the GA signal transduction pathway, but in crosstalk with other phytohormones including auxin, abscisic acid (ABA), ethylene,and jasmonate (JA) [18]. Various plant hormones coordinate with one another,affecting diverse aspects of plant growth and development and influenced by environmental factors such as temperature, light, oxygen, and moisture [19]. DELLAs interact with some key regulatory proteins involved in multiple environmental and hormonal signaling pathways [20]. In this review, we summarize previous studies of DELLA proteins including those investigating the origin and evolution of the DELLA gene family in plants and the interaction and signaling of DELLA proteins, in an attempt to understand the complex regulatory network of plant growth and development involving DELLAs.
2. Evolutionary DELLA-mediated gibberellin signaling and DELLA genes in plants
Since the late 1950s, GAs have been found in both seed- and non-seed-bearing plants, including unicellular and multicellular algae, mosses, and ferns [21,22]. The phytohormone gibberellin has long been known to regulate angiosperm growth via the GADELLA signaling mechanism [19,23–26]. Yasumura et al. [27]defined various distinct stages of the GA-DELLA regulatory mechanism over evolutionary time and concluded that it arose relatively early during land-plant evolution. In particular, the GA-DELLA mechanism was independently established by characterization of the GA-stimulated GID1-DELLA interaction and DELLA growthrepression functions[27,28].The original DELLAs lacked the ability to interact with GID1, although GID1 affinity for DELLAs was present. GID1 does not interact with DELLA in Physcomitrella patens(a bryophyte),but this interaction is found in Selaginella kraussiana(a lycophyte) [27], suggesting that the GID1-DELLA interaction originated in lycophytes (~430 MYA) (Fig. 1). DELLA-mediated growth repression in the control of seed germination and stress responses occurred after the divergence of the lycophytes and before that of the gymnosperms(~300 million years ago)[27].This finding implies that the biological function of DELLA proteins requires not only GID1, but also other proteins.
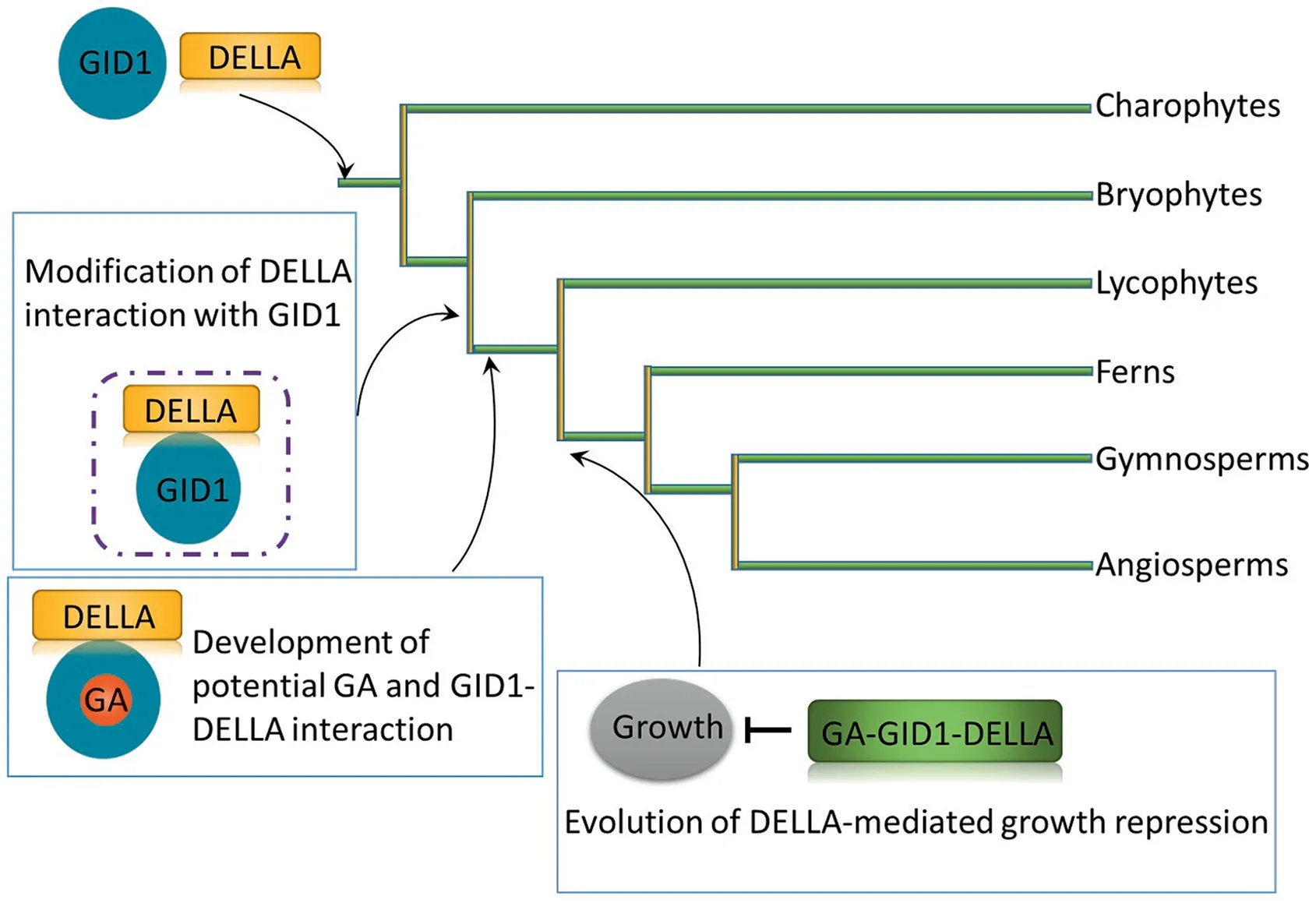
Fig. 1. Evolutionary DELLA-mediated gibberellin signaling. DELLA-GID1 interaction and development of the GA-GID1-DELLA interaction took place after bryophyte divergence (~430 MYA) but before lycophyte divergence (~400 MYA). Growth repression of GA-GID1-DELLA occurred before gymnosperm divergence (~300 MYA). It is unclear whether the growth repression mediated by the GA-GID1-DELLA mechanism occurred before fern divergence.
GID1-mediated GA perception by the GID1/DELLA system similar to the GA feedback regulation in flowering plants is found in Selaginella moellendorffii, but not in P. patens [29]. There are, however,some genes in P.patens encoding proteins homologous to rice GID1: SLENDER RICE 1 (SLR1), and GID2. The functional conservation of GID1/DELLA-mediated GA signaling in moss differs from that in vascular plants. S. moellendorffii contains the conserved domain of DELLAs,which is not present in P.patens,indicating that the conserved region of DELLAs began to evolve to form a mode of GID1/DELLA-mediated GA signaling in ferns [29,30]. DELLAs are split into four main evolutionary branches: algae, bryophytes,monocotyledons, and dicotyledons [31]. Domain and motif analysis shows that lower plants including lycophytes and bryophytes lack classic DELLA conserved regions. For example, SkDELLA contained divergent but conserved domains, while PpDELLAa and PpDELLAb contained more divergent conserved domains [27].
Numerous studies have focused on DELLA genes in vascular plants(Table 1).The DELLA family has expanded in vascular plants owing to gene duplication. DELLAs are present mainly in the form of polygenes,with a few in the form of single genes in rice,wheat,and cotton. Large differences in the numbers of gene families are due mainly to differences among species.Furthermore,expression levels of DELLA genes vary greatly among different tissues of the same species.For example,the DELLA gene family includes AtRGL1,

Table 1Known DELLA genes in plants.
AtGAI,AtRGA,AtRGL2,and AtGRL3 in Arabidopsis.AtGAI is expressed in all tissues, whereas AtRGL1, AtRGL2, and AtGRL3 are expressed only in flowers, fruits, and seeds [32]. In summary, although DELLA-mediated GA signaling occurs in six main evolutionary branches, studying the evolution of DELLA activity requires comparative analysis of the DELLA interactome in different types of plants. In addition, DELLA-mediated GA signaling could have affected adaptation on an evolutionary time scale. The ancestral role of DELLAs or GAs in transcriptional coordination awaits investigation.
3. Conserved domains of DELLA proteins
DELLAs belong to a subgroup of the GRAS family of plantspecific transcription factors (TFs) [33,34], which usually contain a variable N-terminal domain and a highly conserved C-terminal GRAS domain. The N-terminal region of DELLAs differs from those of the rest of the GRAS family [10]. All DELLAs consist of a divergent N-terminal GA-perception region necessary for binding to GID1 and a C-terminal GRAS domain that contributes mainly to repressing GA responses by interacting with multiple regulatory proteins(Fig.2).The N-terminal domain is characterized by DELLA and VHYNP domains,which are required for GID1 interaction,and polymeric Ser/Thr/Val (poly S/T/V) motifs. There were no interactions between PtGID1 and PtDELLA proteins in a 17-amino-acid deletion in the DELLA domain in Pinus tabuliformis[35],indicating that the DELLA domain is essential for the interaction between GID1 and DELLA protein.
A second highly conserved motif (called VHYNP), composed of the amino acid residues TVhynPxxLxxWxxxM, is crucial for interaction between GID1 and DELLA protein. SLR1, a sole DELLA-type protein, suppresses GA signals and overexpression of SLR1 results in a GA-insensitive dwarf phenotype in rice [36,37]. Both DELLA and VHYNP domains at the N-terminal end in SLR1 are essential and sufficient for the GA-dependent GID1-SLR1 interaction [38].A single amino acid insertion in the VHYNP domain of the D8 gene,a typical DELLA gene in maize, leads to the appearance of a dominant dwarf mutation [39]. The poly S/T/V motif is likely not a key component for DELLAs, but possibly participates in phosphorylation or glycosylation of DELLA proteins [40].
Six unique motifs occur in the C-terminal GRAS domain of DELLA proteins: two leucine heptad repeats (LHRs), possibly involved in protein–protein interactions, a putative nuclear localization signal (NLS) [41,42] and three conserved motifs: VHIID,SH2-like, and SAW [43]. The LHRs are critical for protein homodimerization and are likely required for DELLA activity and GAdependent DELLA degradation [44]. In particular, the LHR1 motif is essential for the interaction between DELLAs and TFs such as PHYTOCHROME INTERACTING FACTORs(PIFs),FLOWERING LOCUS C, and BRASSINAZOLE-RESISTANT1 (BZR1) [45]. Loss-of-function mutation of the C-terminal VHIID,PFYRE and SAW motifs in DELLA protein results in a slender phenotype [43], suggesting that these motifs are also necessary for DELLA proteins.
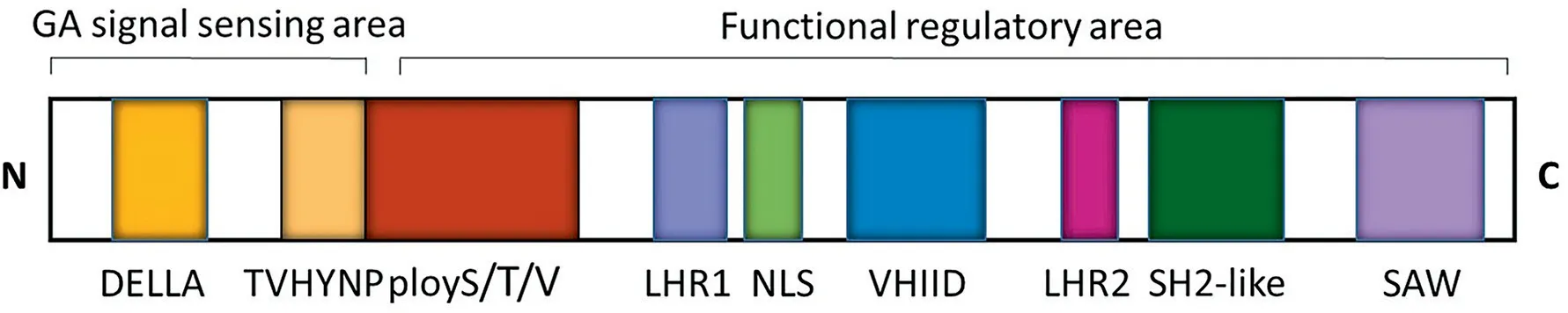
Fig.2. The conserved domains of DELLA proteins.DELLAs consist of a divergent N-terminal GA perception region and a C-terminal GRAS domain.The N-terminal domain is characterized by DELLA and TVHYNP domains,which are required for GID1 interactions,and polymeric Ser/Thr/Val(poly S/T/V)motifs.The C-terminal GRAS domain contains a leucine heptad repeat1 (LHR1) and LHR2, a putative nuclear localization signal (NLS), and three conserved motifs: VHIID, Src–homology 2 like (SH2-like) and SAW.
4. Dellas and the GA signal transduction pathway
GAs are cyclic diterpenoid molecules that modulate almost all aspects of plant growth and development via the GA signal transduction pathway.DELLA proteins interact with downstream TFs in the GA signaling pathway, functioning as core negative regulators and repressing multiple GA responses. The GA signal transduction pathway includes signal perception and signal transduction,involving mainly GID1, DELLA proteins, and other regulatory factors (such as F-box type proteins) that mediate DELLA protein degradation. Without GA binding, the N-terminal extension of GID1 has a flexible structure that is highly sensitive to protease treatment[6]. The SCF complex belongs to a group of E3 ubiquitin ligases and is involved in the ubiquitination of target proteins.SCF E3 ligases are composed of four proteins: cullin1/Cdc53, Rbx1/Roc1/Hrt1, Skp1, and an F-box protein [46]. The F-box protein is involved in multiple signal transduction pathways (GA, JA, ABA,and indole-3-acetic acid (IAA)) as well as various plant responses including drought resistance,salt stress,and heavy metal and temperature stress.The F-box protein recruits the targets and binds to Skp1 at the F-box motif, which consists of ~50 amino acids in the N-terminal region [46,47]. The F-box protein recognizes targets and provides substrate specificity to E3 ligases owing to its secondary structure, which has protein–protein interactions in the C-terminal domain.
F-box proteins SLY1 and GID2 are essential components of SCFSLYand SCFGID2E3 ubiquitin ligase complexes in Arabidopsis and rice, respectively [12]. Both SLY1 and rice GID2 genes encode highly similar F-box proteins, which accumulate extremely high levels of DELLAs. Loss-of-function sly1 and gid2 mutants are GAinsensitive dwarfs,which positively regulate the gibberellin signaling pathway in Arabidopsis and rice [48]. SLY1 and GID2 are also involved in ubiquitination and subsequent degradation of DELLAs.Both SLY1 and GID2 interact directly with DELLAs in the DELLAGA-GID1 complex, indicating that they recruit DELLA proteins for degradation by the 26S proteasome [46]. Increased extracellular GA levels alter conformation in the C-terminal domain of GID1,causing GID1 to bind to GA. The formation of GA-GID1 induces a conformational switch of N-Ex of GID1 to cover the GA-binding pocket, as well as to create a binding domain (exposed hydrophobic surfaces) for DELLA.

Fig.3. Gibberellin signal transduction pathway. Gibberellin-GID1-DELLA: a pivotal regulatory module for plant growth and development. Binding of a bioactive GA induces a conformational switch in the GID1 receptor that promotes interaction with DELLA proteins.Recruitment of an F-box protein promotes polyubiquitination of DELLA by the SCFSLY1/GID2 E3 ubiquitin ligase and subsequent degradation by the 26S proteasome. Degradation of DELLA induces the GA–DELLA-mediated responses.
Binding of GA to its receptor GID1 protein promotes the formation of the GA-GID1-DELLA complex,by inducing a conformational change in the C-terminal domain of DELLA proteins.This stimulates polyubiquitination of DELLA by the SCFSLY1/GID2E3 ubiquitin ligase and subsequent degradation by the 26S proteasome (Fig. 3). Thus,GA promotes plant growth by stimulating degradation of DELLAs.DELLA accumulation inhibits plant growth, resulting in semidwarfism as seen in Green Revolution cultivars. Since the 1960s the Green Revolution has greatly increased cereal crop yields,enabled by wide adoption of fertilizer-responsive semi-dwarf high-yielding cultivars.Wheat Rht-B1b and Rht-D1b alleles encode growth-repressing DELLA genes containing a stop codon that confer dominant GA-insensitive dwarfism. Recent studies [49,50] have revealed potential mechanisms for these alleles. Rht-B1b and Rht-D1b genes can produce N-terminal truncated proteins by restarting translation, and such proteins can be expressed in wheat using transgenic methods.An allelic mutant strain of Rht-D1c with multiple copies of Rht-D1b was not sensitive to GA-mediated degradation,resulting in severe dwarfing of wheat plants[49].
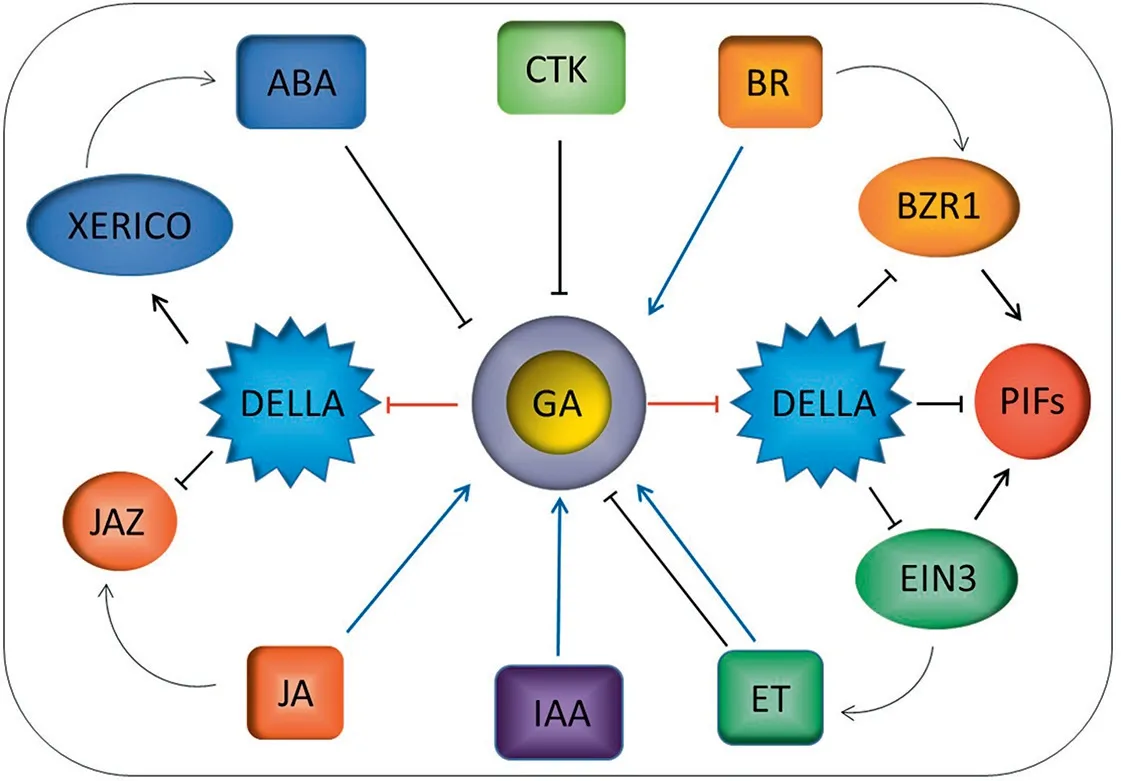
Fig. 4. Crosstalk between DELLA involved in GA signaling and other phytohormones. The GA-GID1-DELLA regulatory module is indicated by red lines. Signals that promote bioactive GA accumulation are labeled in blue lines, whereas signals that reduce GA levels are indicated by black lines.
5. Functions of DELLA proteins
Members of the DELLA protein family are essential for regulating many aspects of plant growth and development,including seed germination, flowering, hypocotyl hook formation, and defense response. In this section, we review some of the main functions of DELLA proteins.
5.1. Crosstalk between DELLAs and other phytohormone signal transduction pathways
DELLAs function directly not only in the GA signaling pathway,but also in other hormone signal transduction pathways in plants.The ethylene signaling pathway regulates root growth by modulating growth-repressing effects of DELLAs [51]. Ethylene influences the biological activity of GA, increasing levels of DELLA proteins and inhibiting expression of flowering-associated genes, thus delaying flowering. DELLAs also interact with DNA-binding domains of ETHYLENE INSENSITIVE 3/EIN3-LIKE 1 (EIN3/EIL1) in the GA signal transduction pathway (Fig. 4). EIN3/EIL1 activates transcription of HOOKLESS 1 (HLS1),a central regulator mediating the input of multiple signaling pathways,by directly binding to its promoter.Thus,GA triggers DELLA degradation,subsequently promoting apical hook formation in cooperation with ethylene partly by inducing expression of HLS1 by relieving the repression of DELLA proteins on EIN3/EIL1 [52]. The RIN-H2 zinc finger factor XERICO, a putative DELLA target, promotes accumulation of ABA,which antagonizes GA effects. Thus, DELLAs may restrict GA responses by modulating both GA and ABA pathways to reduce impacts of abiotic stress.The DELLA protein PROCERA,a gibberellin response inhibitor, promotes ABA-induced stomatal closure and gene transcription via the upregulation of the ABA transporter AIT1.1 in tomato [53]. In summary, the crosstalk between GA and ABA is likely mediated at multiple levels during hormone biosynthesis, signaling, and transport.
GA negatively regulates both stress responses and plant development via crosstalk with the JA signaling pathway (Fig. 4). DELLA mediates JA signaling via competitive binding to JA ZIM-domain 1(JAZ1) protein, a key repressor of JA signaling in Arabidopsis [54].Without GA,stabilized DELLAs bind to JAZ1 and release JASMONATE INSENSITIVE1(JIN1/MYC2),promoting JA signaling.In contrast,GAmediated degradation of DELLAs enhances JAZ1 binding to MYC2 and suppresses MYC2-dependent JA-signaling outputs[55,56].DELLAs also function in modulating the relative strength of JA in response to pathogen stress.GA promotes JA biosynthesis through DELLAs to induce expression of MYB genes (MYB21, MYB24, and MYB57),thereby promoting stamen filament elongation[57].
JA and salicylic acid (SA) are essential defense hormones. DELLAs modulate the plant immune response by controlling the balance of JA/SA signaling, indicating the indispensable role of GA signaling in plant defense against pathogen infection[58].The critical function of SA is to regulate plant defense responses against pathogens. In response to infection with Pst DC3000, DELLA and the plant immune regulator gene ENHANCED DISEASE SUSCEPTIBILITY 1 form a regulatory feedback module, which fine-tunes the plant growth–defense tradeoff in Arabidopsis [59]. As a suppressor of JA, GhJAZ2 interacts with DELLAs and indirectly activates the GA signal pathway to regulate flowering, seed germination, fruit branch angle, and other growth and development processes in Gossypium hirsutum [60].
Brassinosteroids (BRs) are another class of hormones for plant growth and development.Plants defective in BR biosynthesis or signal transduction pathways show dramatic growth-deficient phenotypes, including dwarfism,reduced apical dominance and fertility,delayed senescence and flowering, and light-grown phenotypes in the dark [61,62]. BR and GA both regulate photomorphogenesisassociated hypocotyl elongation in Arabidopsis [63,64], and act interdependently via direct interaction between BZR1 and GA-inactivated DELLA transcription regulators (Fig. 4). DELLAs directly interact with BZR1 and block BZR1-DNA binding both in vitro and in vivo, inhibiting expression of target genes. Thus, GA promotes DELLA degradation and releases DELLA-mediated inhibition of BZR1 during hypocotyl elongation.BR and GA have complex crosstalk depending on species or tissue.Under physiological conditions,BR promotes GA accumulation by regulating the expression of GA metabolic genes including GA20ox-2, GA3ox-2, and GA2ox-3 to stimulate cell elongation in rice[65].BR promotes GA biosynthesis and an increased GA level destabilizes DELLA suppressor to further release BZR1 activity in Arabidopsis[66,67].
GA promotion of cell elongation requires BR signaling,whereas BR or active BZR1 suppress the GA-deficient dwarf phenotype.Both BZR1 and PIFs are required for GA promotion of hypocotyl elongation.When GA levels are low,DELLAs accumulate and directly inactivate PIF4, thereby blocking expression of cell elongation genes when plants are grown in dark, cold, shaded or high-temperature conditions[68–70].Thus,the DELLA-BZR1-PIF4 complex functions directly in timing of hypocotyl elongation and co-regulates plant growth and development (Fig. 4). IAA, the most abundant auxin,controls growth of Arabidopsis roots by modulating the cellular response to GA[71].The polar transport of IAA is closely associated with the level of DELLAs in root tips.Inhibition of polar IAA transport or removal of the apical tip reduces the degradation of DELLAs,terminating root growth. Antagonistic interactions between GA and cytokinin(CTK) may also regulate plant growth and development.CTK stimulates the expression of GAI and GRA by inhibiting the expression of gibberellin 3-oxidase,which is particularly important for the control of bioactive GA levels[72].
5.2. Role of DELLA protein in seed germination
When seeds are mature, the embryo stops growing and enters dormancy. In this stage, seeds cannot germinate until they sense favorable environmental conditions. Seed germination influenced not only by external factors such as light and soil components,but also by hormone regulation. DELLA-mediated GA signaling is involved in seed germination [32,73]. PHYTOCHROMEINTERACTING FACTOR 3-LIKE5 (PIL5/PIF1/bHLH015), a basic helix-loop-helix (bHLH) TF, inhibits seed germination by reducing bioactive GA levels in the absence of light (Fig. 5). Phytochromes promote the degradation of PIF1 in seeds, leading to increases of bioactive GA and degradation of DELLA proteins and finally triggering seed germination.PIF1 binds directly to GAI and DELLA protein REPRESSOR OF GA 1–3 (RGA) promoter regions to activate their expression. DELLAs may also interact with SPATULA, a PIF homolog,and SCARECROW-LIKE 3(SCL3)to cross-regulate seed germination and root growth, respectively (Fig. 5). SOMNUS encodes a CCCH-type zinc finger protein that inhibits light-dependent seed germination by regulating ABA and GA metabolic genes [74]. The DELLAs-ABA-INSENSITIVE 3 (ABI3)-ABI5 complex combines with the SOMNUS promoter to activate SOMNUS expression and other high-temperature-inducible genes,thereby regulating seed germination at high temperatures [75]. INDUCER OF CBF EXPRESSION 1 physically associates with ABI5 and DELLA proteins,and negatively regulates ABA responses during seed germination [76].

Fig. 5. Roles of DELLAs and DELLA interactions in plant growth and development.Proteins that interact with DELLA are highlighted in brown and blue.DELLAs act as transcriptional activators and repressors (ovals and rounded rectangles, respectively)to regulate plant development.FLC,FLOWERING LOCUS C;ICE1,INDUCER OF CBF EXPRESSION1; SPT, SPATULA; SCL3, SCARECROW-LIKE 3; SOC1, SUPPRESSOR OF OVEREXPRESSION OF CO1; RAP2.3, RELATED TO APETALA2.3; ARR1, ARABIDOPSIS RESPONSE REGULATOR1.
Seed germination is determined by multiple effects of plant hormones and extensive crosstalk among GA, ABA, ethylene, and BR[77,78]. Seed germination is regulated antagonistically mainly by ABA and GA,which respectively promote and repress seed germination.DELLAs differentially regulate the expressions of multiple protein kinase genes to control seed germination in crosstalk[79].
5.3. Role of DELLA proteins in flowering
The transition from vegetative to reproductive development is the most vital phase transition in plant development. Flowering is both important and essential for many aspects of agriculture.Many crops consist of flowers or their products(fruits and seeds),including the cereals that provide most of the calories for a majority of the world’s human and livestock populations.DELLAs can delay the floral transition by interfering with the transcriptional activities of SQUAMOSA PROMOTER BINDING PROTEIN–LIKE (SPL) proteins[80,81]. SPLs directly activate floral-promoting MADS box genes and the plant-specific TF LEAFY (LFY) to promote the juvenile-toadult phase transition and flowering [82,83]. DELLAs can interact with SPLs to interfere with SPL transcriptional activity and consequently delay floral transition by inactivating miR172 in leaves and MADS box genes under long-day conditions(Fig.5).
GA promotes flowering by activating LFY and SUPPRESSOR OF OVEREXPRESSION OF CO1 (SOC1). However, DELLAs delay flowering by inhibiting the expression of both LFY and SOC1 under shortday conditions[25].CONSTANS(CO),a B-box zinc finger TF,is a key component that accelerates flowering in Arabidopsis under long-day conditions [84,85]. DELLAs physically interact with CO to inhibit the association of CO with nuclear factor YB2 (NF-YB2),resulting in the repression of FLOWERING LOCUS T transcription in leaves and delay of flowering under long-day conditions [86].GAI and RGL1 positively and negatively modulate flowering time by interacting with respectively WRKY TFs (WRKY12, WRKY13)and WRKY75 in Arabidopsis[87,88].Four Arabidopsis RING proteins(BOTRYTIS SUSCEPTIBLE1 INTERACTOR(BOI),BOI-RELATED GENE1(BRG1),BRG2,and BRG3)repress a subset of GA responses including seed germination, the juvenile-to-adult phase transition, and flowering by binding to the promoter regions of GA-responsive genes and repressing their transcription [89].
FLOWERING LOCUS C (FLC) is another central floral regulator that integrates autonomous and vernalization pathways to finetune flowering time. DELLAs interact with FLC and increase the transcription inhibition ability of FLC.Recent study[45]suggested that the interaction between the C-terminus MADS domain of FLC and the LHRI domains of DELLAs integrates various signaling inputs in flowering-time control. In Arabidopsis, a proportion of DELLAs is also conjugated to the small ubiquitin-like modifier protein, which stabilizes DELLAs. The small ubiquitin-like modifier protein protease-deficient OVERLY TOLERANT TO SALT 1 can control DELLA activity in filament elongation and stamen development under salt-stress conditions [90]. Cold-induced or constitutive expression of C-repeat binding factor 1 promotes RGA accumulation and reduces plant growth. Transgenic plants that constitutively express C-repeat binding factor 1 accumulate less bioactive GA and as a consequence exhibit dwarfism and late flowering[91]. In tomato, PROCERA (PRO) is the only DELLA protein, and loss-of-function mutations in the PROCERA gene result in a delayed-flowering phenotype [92].
5.4. Role of DELLAs in hypocotyl elongation and apical hook development
GAs promote growth of etiolated seedlings and hypocotyl elongation.DELLAs block PIF transcriptional activity and regulate gene expression by binding with PIF3 and PIF4 promoters, and thereby stop PIF-mediated hypocotyl elongation [68,69] (Fig. 5). TYPEONE PROTEIN PHOSPHATASE 4(TOPP4),acts as a positive regulator in the GA signaling pathway to regulate hypocotyl elongation.TOPP4 directly binds and dephosphorylates RGA and GAI,confirming that the TOPP4-controlled phosphorylation status of DELLAs is associated with their stability [93]. DELLAs interact with AUXIN RESPONSE FACTOR 6 (ARF6) to form the BZR-ARF-PIF/DELLA(BAP/D) transcriptional module, which coordinately regulates cell elongation of the hypocotyl [94–98].
PIFs,a subgroup of bHLH TFs,are emerging as signal integrators in multiple hormone pathways during plant growth and development. Phytochromes inhibit hypocotyl elongation resulting from the degradation of PIFs, and simultaneously stimulate accumulation of DELLAs. The binding of DELLAs to the bHLH domain of PIFs blocks their DNA binding to their target gene promoters,leading to short hypocotyls in light-grown plants [69]. Up-regulation of PIFs(PIF4 and PIF3-LIKE1)rapidly stimulates expression of DELLA genes,such as GAI and RGA, in apical buds of Populus after exposure to a short photoperiod, resulting in decreased sensitivity to GA [99].In turn, down-regulated GA levels resulting from modulated expression of GA 20-oxidase under a short photoperiod may increase DELLA protein stability [99]. Thus, the interaction between DELLA and PIFs integrates GA and light signals to regulate plant growth and development,especially in photomorphogenesis.
Crosstalk between GA and ethylene signaling has long been known to occur in the regulation of Arabidopsis apical hook development.Plant responses to ethylene are mediated by regulation of EIN3-binding F-box protein 1 and 2 dependent degradation of the ethylene-regulated TF EIN3[100].DELLAs physically associate with DNA-binding domains of EIN3/EIL1 proteins,which in turn repress EIN3/EIL1-regulated HLS1 expression and apical hook formation(Fig. 5). RELATED TO APETALA2.3, a DELLA-interacting protein, is involved in the regulation of apical hook development[101].Thus,GA and ethylene signaling coordinate to regulate apical hook development at least partly via the interaction between DELLAs and EIN3/EIL1 proteins.
5.5. Role of DELLAs in nitrogen use efficiency (NUE) and yield
Plant height is influences not only lodging resistance, but photosynthesis, assimilate distribution, and grain yield. In Hordeum vulgare, the DELLA gain of function mutant Sln1d exhibited reduced plant height [102]. Knockdown of SLR1 by RNA interference (SLR1-RNAi) resulted in taller rice, whereas plant height was reduced when the SLR1 protein was overexpressed [103].DELLA proteins associate with TFs to control stem elongation in response to gibberellin. Growth-regulating factors (GRFs), also known as 14-3-3 proteins, belong to a small family of TFs distributed in all known seed plants. Several reports support the notion that GRFs are involved in various aspects of tissue differentiation and organ development. GRFs contain a highly variable C-terminal domain and two highly conserved protein domains in their N terminus: WRC (Trp–Arg–Cys) and QLQ (Gln–Leu–Gln).The C-terminal regions of GRFs are essential for their transactivation activity and are highly variable with respect to the composition and number of amino acid residues [104]. Thus, they show only limited similarities to one another. The WRC domain contains a nuclear localization signal and a C3H-type zinc (Zn)finger motif, which is involved in DNA binding [105]. The QLQ domain is a stretch of ~36 amino acid residues and shows similarity to the N-terminal part of the yeast SWITCH2/SUCROSE NONFERMENTING 2 (SWI2/SNF2) chromatin remodeling ATPase that interacts with SNF11 [104]. The QLQ domain mediates the interaction between GRFs and transcriptional coactivator GRFINTERACTING FACTORs (GIFs) [106].
The root system and tillering are plant architecture traits that strongly influence crop yield. In the GA signaling pathway, the receptor GID1 and SKP1-CUL1-F-box type E3 ligase SCFSLEEPY1/GID2together promote degradation of the DELLA repressor proteins to relieve their repression of GA. Molecular genetic study [107] have revealed that MONOCULM1 encodes homologous GRAS proteins that act as key factors promoting tillering in rice. A recent study[108] showed that GAs may block SNF1-RELATED KINASES2 activity and promote degradation of MONOCULM1 mediated by the ANAPHASE-PROMOTING COMPLEX/CYCLOSOME tiller enhancer,ultimately inhibiting root growth and tillering in rice. GRF4 is a transcriptional regulator of multiple nitrogen-metabolism genes.GRF4 has an especially highly conserved protein N-terminal domain, which enables it to interact with DELLAs to regulate the expression of various nitrogen-metabolism genes in the GA pathway. GRF4 and GRF4-GRF-interacting factor 1 (GIF1) TFs form a plant-specific complex to regulate transcriptional networks, such as for nitrogen absorption and metabolism [109]. GRF4 promotes both nitrogen uptake and assimilation,whereas the DELLA protein SLR1 inhibits these processes by reducing GRF4 accumulation and GRF4-GIF1 activation in rice [110]. SLR1 can also bind GRF4 to compete with the GRF4 and GIF1 interaction and interfere with GRF4-GIF1 complex formation, thereby reducing NUE. Thus, balancing GRF4-DELLA and increasing GRF4 abundance will improve the NUE of semi-dwarf Green Revolution cultivars (Fig. 5).
NITROGENMEDIATED TILLER GROWTH RESPONSE 5 (NGR5),which is identical to the previously reported SMALL ORGAN SIZE 1 and REDUCED LEAF ANGLE1, interacts with DELLA to boost rice yield by increasing both tiller number and NUE in nextgeneration Green Revolution rice[111–114](Fig.5).NGR5 encodes a TF with an APETALA2 DNA binding domain. The loss-of-function mutant ngr5 in rice displays fewer tillers under high nitrogen concentrations, and its expression level is strongly induced by high nitrogen supply.NGR5-driven recruitment of polycomb repressive complex 2 (PRC2) promotes repressive histone H3 lysine 27 trimethylation modification of branching inhibitory genes Dwarf14 and squamosa promoter binding protein-like-14, thereby increasing nitrogen supply and tiller number. NGR5 competes with DELLAs to interact with the gibberellin receptor GID. Thus, a motif in the APETALA2-R2 (repeated units 2) domain of NGR5 promotes the interaction of NGR5-GID1, causing polyubiquitination of NGR5 by the SCFGID2E3 ubiquitin ligase and subsequent destruction by the 26S proteasome. Thus, the regulation of GA-NGR5-DELLA influences NUE to increase the sustainability of crop production for food security. The interaction between GRF4-NGR5 and DELLAs can increase yield and NUE, particularly at relatively low levels of nitrogen supply.
5.6. Role of DELLAs in stem elongation, trichome development, and root meristem
A recent study [115] showed that DELLAs can bind to several class I TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR (TCP) proteins at the shoot apex to regulate plant height.As plant-specific TFs,TCP proteins are present only in angiosperms and are involved in strigolactone signal transduction pathways.DELLAs prevent TCP from activating cell-cycle genes to regulate stem elongation by binding to the DNA-recognition domain of TCP TFs(Fig.5).JA and GA synergistically regulate trichrome development via interaction between JAZs and DELLAs. JAZ proteins bind and inhibit the activity of a wide array of TFs, including MYC2, MYC3, and MYC4, which regulate major aspects of JAmediated gene expression. Both JAZs and DELLAs interact with the WD-repeat/bHLH/MYB complex to modulate JA and GA synergy in regulating trichome development (Fig. 5).
CKs and GAs perform antagonistic regulation of multiple developmental processes such as shoot apical meristem and root growth(Fig. 5). ARABIDOPSIS RESPONSE REGULATOR 1, a primary CKresponse TF, activates the gene SHY2/IAA3, which negatively regulates the PIN genes encoding auxin transport facilitators to balance root meristem establishment and maintenance.RGA interacts with ARABIDOPSIS RESPONSE REGULATOR 1 to control the root meristem in Arabidopsis [116].
5.7. Role of DELLAs in other aspects of plant development
DELLAs have also been reported to function in regulating stomatal closure,axillary bud formation,and fruit development.Stomatal closure is a common adaptation response of plants to the onset of drought. In tomato, the DELLA protein PROCERA acts on guard cells by promoting stomatal closure. The loss-of-function procera mutant exhibited increased stomatal conductance and rapid wilting under water-deficit stress. Transgenic tomato overexpressing constitutively active stable DELLA proteins displayed the opposite phenotype [117]. The formation of lateral branches contributes to the remarkable developmental plasticity of plants,allowing plants to alter their architecture to adapt to stressful environmental conditions.Tillering,the process of branching in crop plants,determines the number of panicles per plant, a key component of grain yield usually negatively correlated with plant height[103].As axillary buds are precursors of lateral branches and tillers,their formation is associated with GA content: low GA content promotes axillary bud formation and exogenous applications of GA reduce axillary bud formation [118].
In Arabidopsis,DELLA proteins can interact with SPL9 and attenuate the repressing activity of SPL9, promoting the initiation of axillary buds. In wild-type tomato plants, the oldest axillary buds commence growth before newer buds closer to the apex. In the procera mutant,the growth of axillary buds furthest from the apex is repressed, while those closest to the apex grow normally [119].In rice,overexpression of SLR1 resulted in plants with increased tiller number, whereas tiller number was decreased in SLR1 knockdown plants [103]. Fruit development is regulated by DELLA. In tomato, the DELLA protein SlDELLA interacted with auxinsignaling components SlARF7/SlIAA9 to regulate fruit initiation[120]. Silencing of DELLA resulted in parthenocarpic fruits, which were smaller when mature, with a distinctive elongated shape[121].
In summary, DELLAs interact with numerous proteins and participate in crosstalk with other phytohormone signal transduction pathways.These findings may offer strategies for plant breeding to improve crop plant architecture.
6. Transcriptional regulation and post-translational modifications of DELLA
6.1. Transcriptional regulation of DELLA
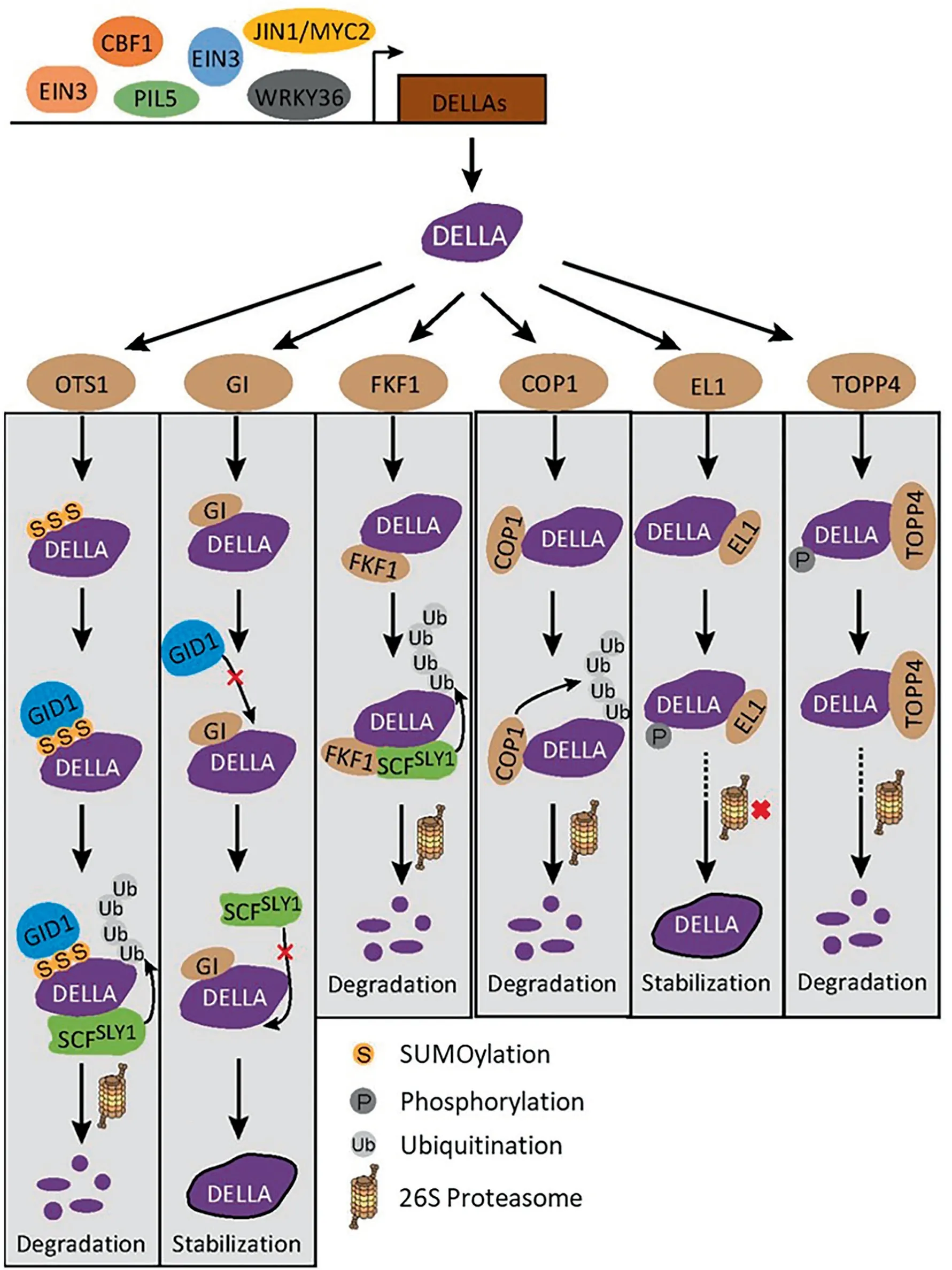
Fig.6. The regulation network of DELLA.At the transcript level,upstream regulators PIL5,EIN3,WRKY36,and JIN1/MYV2 bind directly to DELLA promoters to increase their expression.At the posttranslational level,OTS1,GI,FKF1,COP1,EL1,and TOPP4 interact with DELLA protein and mediate SUMOylation,ubiquitination,phosphorylation,and dephosphorylation of DELLA.
The roles of DELLA in the GA signaling pathway imply that the regulation of DELLA abundance and stability is critical to maintaining normal plant growth and development. In fact, the expression of DELLA is tightly regulated by several upstream regulators,including PIL5, EIN3, WRKY36, JIN1/MYC2, and CBF1. PIL5 is a light-labile bHLH TF that interacts with phytochrome, which can directly bind to G-box elements(CACGTG)in the promoters of GAI and RGA and regulate their expression in Arabidopsis seeds [122](Fig. 6). EIN3, a TF in the ethylene signaling pathway, strongly induces promoter activity of RhGAI1 by directly binding to EBS sites(AATTCAAA) in Rosa hybrida [123]. WRKY transcription factors are a superfamily of plant TFs, which contain one or two conserved WRKYGQK motifs followed by a C2H2 or C2HC zinc-finger motif.OsWRKY36 in rice activates transcription of SLR1 by directly binding to W-boxes ((T)TGAC(C/T)) in its promoter [124]. IN1/MYC2 encodes a MYC TF involved in the JA signaling pathway. When JA is present, the JA-receptor/F-box COI1 promotes the polyubiquitylation and subsequent destruction of degradable JAZ1, leading to the release of JIN1/MYC2. The released JIN1/MYC2 promotes RGL3 expression by directly associating with a G-box-related motif(CACATG) in the promoter of RGL3 [125]. The CBF/DREB1 protein CBF1 is a member of AP2/EREBP (Apetala2/ethylene-responsive element binding protein) transcriptional regulators. Overexpression of CBF1 increased both DELLA gene transcripts and protein accumulation in Arabidopsis, indicating that CBF1 could modulate DELLA-accumulation both transcriptionally and posttranslationally[91].
6.2. Posttranslational modifications of DELLAs
In the GA signaling pathway, the binding of GA with the GA receptor stimulates GID1-GA-DELLA complex formation, which in turn targets DELLA protein ubiquitination by SCFSLY1E3 ubiquitin ligase, and the ubiquitinated DELLAs are subsequently degraded by the 26S proteasome system to trigger GA responses [126]. In addition to the GA receptor GID1, other proteins have been reported to modulate the stability of DELLA. SUMOylation, a post-translational modification of target proteins, is involved in regulating DELLA stability.SUMO proteases OTS1 and OTS2 deconjugate SUMO from DELLA protein, and seedlings lacking OTS1 and OTS2 exhibited increased DELLA levels in Arabidopsis[127](Fig.6).The SUMOylated DELLA enables the interaction with GID1 via the SIM-SUMO domain, subsequently inducing ubiquitination and degradation of DELLA proteins.GIGANTEA(GI)is a core component implicated in gating of the response to GAs via regulation of pivotal factors in the signaling by this hormone [128]. GI interacts with DELLA, preventing access of the GA receptor GID1 to RGA protein and interfering with the ubiquitination and degradation of DELLA.FLAVIN-BINDING KELCH REPEAT F-BOX 1, encoding an F-box protein with three domains (LOV, F-box, and Kelch repeat), is a key component of the SCF-type E3 ligase complex. The Kelch repeat domain of FKF1 interacts with the GRAS domain of RGA and GAI to negatively modulate DELLA protein abundance by promoting DELLA protein ubiquitination and degradation [129].
Besides the GA-dependent pathways,there is a GA-independent process for DELLA polyubiquitination and degradation.CONSTITUTIVELY PHOTOMORPHOGENIC1 is an E3 ubiquitin ligase that promotes proteasome-dependent degradation of several TFs. With the help of its functional partner SUPPRESSOR of phyA-105 protein,COP1 interacts with DELLA proteins RGA and GAI in nuclear bodies,leading to their polyubiquitination and degradation by the 26S proteasome[130].Protein phosphorylation and dephosphorylation are the major posttranslational modifications regulating diverse cellular processes. Phosphorylation of SLR1 is important for the regulation of its activity and stability. In rice, the early flowering 1 (el1) mutant displays enhanced GA responses [131]. Functional characterization reveals that EL1, which encodes a casein kinase I, interacts with the rice DELLA protein SLR1 and phosphorylates the N-and C-termini of SLR1. Phosphorylation of SLR1 suppresses the GID1-GA-mediated degradation of SLR1 and sustains its activity, inhibiting the GA signaling pathway. TOPP4 interacts with DELLA protein and dephosphorylates DELLA protein to promote the GA-induced degradation of DELLA proteins through a 26S proteasome pathway [93].
7. Conclusion and perspectives
DELLA proteins function as key transcriptional regulators that mediate the effects of GA on plant growth and development.DELLAs are members of the plant-specific GRAS protein family,which possibly originated in mosses and ferns. DELLAs consist of a variable N-terminal region and a highly conserved C-terminal GRAS domain. The former is essential for binding to GID1 and the latter is necessary for GA repression. During evolution, the original DELLAs lacked the ability to interact with GID1, and the GID1-DELLA interaction originated in lycophytes. For this reason,it is desirable to investigate when and how DELLA shaped its functional N-terminal region during plant evolution. DELLA growth-repression functions in land-plant growth including seed germination and stress responses arose after the divergence of the lycophytes and before that of the gymnosperms.It is of interest to determine in future investigations how exactly DELLAs and the N-terminal region evolved in charophytes or another early organism. DELLAs may promote coordination between transcriptional programs during plant evolution and function as regulatory hubs in transcriptional programs in higher plants [132], but it will be important to elucidate the molecular mechanisms of the process of DELLAs acting during land-plant evolution.
Members of the DELLA gene family have been studied mainly in vascular plants. The DELLA gene family includes AtRGL1, AtGAI,AtRGA, AtRGL2, and AtGRL3 in Arabidopsis [32]. D8 and SLR1 of the DELLA gene family are found in maize and rice, respectively[133,134]. However, more comprehensive studies of the DELLA gene family are needed,especially in non-vascular plants.Comparative studies of DELLAs between non-vascular and vascular plants will shed light on the role of DELLA in the process of plant vessel formation.Identifying potential members of the DELLA gene family and their biological functions in non-vascular plants are of great interest as well. DELLA proteins are key components in the evolution of the GA signaling pathway:they have functioned in interactions with GID1 and regulation of downstream signal transduction throughout plant development since the divergence from lycophytes. However, the phylogenetic origin and evolution of DELLAs and the interaction of GID1 with DELLA are not fully understood and invite study.
Accumulated evidence [5,25,51,57,61,68,69] indicates that DELLAs can act as key regulators during crosstalk between gibberellin and other signaling pathways which are involved in almost all processes of plant development including seed germination,stem elongation,leaf expansion,flowering induction,hypocotyl elongation,and apical hook development.DELLAs interact with various kinds of TFs and other proteins to regulate the GA signaling pathway. Besides the developmental processes above, identifying new biological roles of GA, especially potential roles in plant adaptation,is a major area of study.GA biosynthesis and signaling pathways have been solved using Arabidopsis mutants. Although GA biosynthesis mutants and DELLA mutants have been widely used in agriculture for more than 50 years, it is still a mystery how DELLA mediation of GA signaling pathways contributes to crop agronomic traits. Identifying DELLA-regulatory mechanisms in agronomic traits may lead to improvement of crop quality and yield.
CRISPR/Cas9-based targeted genome-editing technologies have emerged as a powerful technique.These tools are widely employed in crop genome engineering, both for exploring gene function and as a modern breeding technique for trait improvement,defined as the development of new crop varieties with high yield, resistance to biotic and abiotic stresses, and high nutritional value [135].Recently,the CRISPR-Cas9 editing system has been widely applied for modifying endogenous genes. In Brassica napus, fatty acid desaturase 2 gene (FAD2) encodes an enzyme that catalyzes the desaturation of oleic acid; modification of FAD2 using the CRISPR/Cas9 system resulted in increased oleic acid content in seeds[136].In tomato,CRISPR/Cas9 mutagenesis of the engineered self-pruning 5G (SP5G) gene conferred early flowering and more bushiness, which in turn resulted in an early harvest [137,138].
DELLA and its components are crucial regulators that function directly in growth and development during various plant developmental stages.DELLA governs agricultural traits such as seed germination, shoot and root elongation, reproductive transitions,flowering,tillering,and fruit patterning,which modulate grain size and yield.In the future,with the help of genome editing tools such as CRISPR-Cas9 along with standard genetic engineering tools, targeted editing of DELLA and its components in crops will provide a much more elaborate blueprint for remodeling and developing crop species with desirable agronomic traits. For example, researchers can edit DELLA to obtain semi-dwarf mutant cultivars with improved lodging resistance and higher yield in rice, wheat and maize. Editing of DELLA also makes it possible to obtain earlymaturing cultivars in tomato and cotton.Modifying DELLA may lead to new crops that meet the demands of diverse environments and human needs,and may even start a new Green Revolution.
CRediT authorship contribution statement
Huidan Xue:Writing - original draft.Xiang Gao:Software.Peng He:Writing-review&editing.Guanghui Xiao:Supervision.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (32070549), Young Elite Scientists Sponsorship Program by CAST (2019-2021QNRC001), Shaanxi Youth Entrusted Talent Program (20190205), Fundamental Research Funds for the Central Universities (GK202002005), State Key Laboratory of Cotton Biology Open Fund (CB2020A12, CB2021A21, CB2021A05),and the China Postdoctoral Science Foundation (2020M683549).

- The Crop Journal的其它文章
- Far-red light: A regulator of plant morphology and photosynthetic capacity
- A rice XANTHINE DEHYDROGENASE gene regulates leaf senescence and response to abiotic stresses
- Dissection of heterotic loci for grain yield using interconnected chromosome segment substitution lines in rice
- A soybean NAC homolog contributes to resistance to Phytophthora sojae mediated by dirigent proteins
- The boron transporter SiBOR1 functions in cell wall integrity, cellular homeostasis, and panicle development in foxtail millet
- RNAi-mediated suppression of the abscisic acid catabolism gene OsABA8ox1 increases abscisic acid content and tolerance to saline–alkaline stress in rice (Oryza sativa L.)