
大气CO2浓度升高条件下稻稗共生系统中稗草对水稻光合生理的影响*
2022-03-22 11:05:38金殿玉谢立勇赵洪亮何雨桐林而达
中国农业气象 2022年3期
金殿玉,谢立勇**,赵洪亮,李 颖,韩 雪,何雨桐,林而达
大气CO2浓度升高条件下稻稗共生系统中稗草对水稻光合生理的影响*
金殿玉1,谢立勇1**,赵洪亮1,李 颖1,韩 雪2,何雨桐1,林而达2
(1. 沈阳农业大学农学院,沈阳 110161;2.中国农业科学院农业环境与可持续发展研究所,北京 100081)
稗草是水稻田的恶性杂草之一,严重影响水稻的生长发育和产量形成。为明确CO2浓度升高条件下稗草和水稻的光合生理响应及其竞争关系变化,以吉粳88为研究对象,利用开放式CO2浓度富集系统(FACE系统)开展模拟试验。试验设置2个CO2浓度,分别为自然大气CO2浓度(400µmol·mol−1)和高CO2浓度(550µmol·mol−1),高CO2浓度环境应用FACE系统进行调控;每种CO2浓度处理中设2种种植方式,分别为清种水稻和水稻与稗草混种,稗草与水稻种植密度比为1:5,在水稻各生育期测定相应的光合生理指标并进行分析。结果表明:CO2浓度升高使水稻每穴穗数显著增加,结实率也有所提高,最终使水稻产量显著提高;稗草与水稻混种使水稻结实率显著降低,水稻千粒重显著增加,最终使水稻产量显著降低;CO2浓度升高和稗草互作使水稻千粒重显著提高,但对产量影响并不显著。CO2浓度升高使水稻干物质量显著提高,稗草使水稻干物质量显著降低;而CO2浓度升高和稗草互作对水稻干物质影响不显著。CO2浓度升高使水稻剑叶净光合速率、胞间CO2浓度及SPAD值显著升高,使水稻剑叶气孔导度和蒸腾速率显著降低,稗草显著降低了水稻剑叶净光合速率、胞间CO2浓度、气孔导度、蒸腾速率、瞬时水分利用效率及SPAD值;CO2浓度和稗草互作使水稻剑叶净光合速率先降低后升高。水稻抽穗后CO2浓度升高对水稻光合作用的影响大于稗草对水稻光合作用的影响。
气候变化;二氧化碳;稗草;水稻;光合作用
全球气候变化对自然生态系统的影响广泛而深刻,受到越来越多的关注。农业是受气候变化影响最敏感领域之一,水热条件的变化直接影响作物生产以及粮食供给和安全。大气二氧化碳(CO2)浓度增高既是气候变化的驱动力,也直接影响作物光和代谢从而影响作物产量和品质。因此,全球针对大气CO2浓度升高对农作物相互影响开展了大量的研究。目前,大气CO2浓度已增至(403.3±0.1)μmol·mol−1,较工业革命前大气CO2水平提升了45%。尽管全球在努力减排CO2,但预计21世纪中叶大气中的CO2浓度将大概率达到或超过550μmol·mol−1,21世纪末将超过700μmol·mol−1[1]。CO2是作物光合作用的底物,通过影响作物的光合作用来影响其生长发育和产量形成。水稻是利用C3光合途径固定空气中的碳,属于C3植物(约占全球所有植物的95%)[2]。稗草属于C4植物,生长特点是发生速度快、发生量大。基于C3和C4光合作用途径的羧化动力学,随着大气CO2浓度的增加,C3作物的水稻与C4植物的稗草相对竞争力可能发生变化,因而对粮食生产提出新的威胁和挑战[3]。关于CO2浓度升高对作物影响的研究方法主要有开顶式气室(Open Top Chambers,OTCs)和开放式CO2浓度富集系统(Free-air CO2Enrichment,FACE)。由于FACE系统与自由大气相通,系统内部环境与外界大气环境相近,被认为是评估作物生产对CO2与气候变化的实际响应的最佳方法[4]。
杂草是限制水稻高产稳产的生物因素之一,杂草出现的持续时间、杂草密度、杂草种类和作物播种方法会显著影响竞争和产量损失的幅度,并可能导致水稻产量损失21%~79%[5−6]。北方稻田中,禾本科的稗草()是最优势杂草,生长速度快,相对生长速率显著高于水稻,给水稻造成郁蔽环境,并为病虫提供理想条件。稗草通过对光、温、水、气以及土壤养分的大量吸收和掠夺,严重弱化了水稻生长态势,影响水稻的产量和品质,甚至1m2水稻田有一株稗草,水稻即可减产11.6%[7]。清除稻田杂草一般使用除草剂,不仅增加成本,而且危害水稻幼苗,影响生长发育,更带来严重的农药残留等环境问题和食品安全等社会问题。水稻的根系性状,包括根长、根系干重、根系吸收面积和根系氧化活性,与水稻产量呈显著正相关[8−9],稗草显著降低了水稻抽穗期和灌浆期的根干重、长度密度、总吸收面积、活性吸收面、氧化活性、根和籽粒中玉米素+玉米素核苷含量、叶片光合速率、地上部干重以及籽粒中蔗糖合酶(SuSase)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)和淀粉合酶(StSase)活性[10]。当水稻与稗草一起种植时,随着稗草密度的增加,根系氧化活性、根系伤流液等含量降低[11]。地上部生物量、叶片光合速率和籽粒蔗糖合成酶活性等地上部性状在水稻产量形成中起着重要作用[12]。稗草与水稻的共生导致了水稻胞间CO2浓度的显著降低,稗草对水稻光合作用的影响主要是通过增强水稻的蒸腾速率、降低其气孔导度来实现的[13]。牛玺朝等[14]研究发现,稗草甚至可以导致稻米的品质显著下降。
CO2浓度升高条件下C3植物对C4植物的竞争优势增加[15−16]。稗草对水稻产量的影响因稻−稗共生时间、水稻品种和稗草种类等而有所差异,减产幅度较大,稻−稗共生时水稻剑叶光合速率的降低是导致水稻生产力下降的重要原因之一。水稻进入穗分化始期后稗草生长迅速,株高逐渐超过水稻。株高在水稻和稗草共生中发挥着重要作用,相对较高的植物更易捕获光照。此外,较高植物还会为邻近相对较矮的植物遮阴,使共生群体透光率降低,造成光合速率降低[17−18]。在稻稗共生的条件下,水稻增加茎的生物量分配,减少根的生物量分配;趋向于生长地上部分,以获取阳光进行光合作用,而稗草则刚好相反,趋向于生长地下部分,以吸收更多的营养[19]。FACE条件下水稻、稗草比例为1∶1时,水稻与稗草的生物量、产量、LAI、茎蘖和NAR比率均增加,水稻稗草的竞争关系发生变化,水稻(C3植物)竞争能力增加,稗草(C4植物)竞争能力下降[15]。孙富芝等[20]研究表明大气CO2浓度升高水稻相对于稗草的竞争优势增加,但进入生殖生长后水稻的竞争优势减弱;且水稻的竞争优势还与稗草发生的时期有关。当稗草相对水稻出土较晚时,其竞争能力明显降低。
Alberto等[21]采用气室水培试验设置不同种植比例的水稻和稗草,研究发现,在昼夜温度为27℃/21℃时,CO2浓度升至594μmol·mol−1使水稻地上部生物量显著增加47%,籽粒产量显著增加55%,而稗草的生物量和籽粒产量均无显著变化。在两物种混植小区,CO2浓度升高使水稻生物量的比例相对于稗草显著增加。两物种比较,CO2浓度升高使水稻的竞争优势增加,而稗草的竞争力减弱。然而在较高的生长温度(37℃/29℃)下,CO2浓度升高对水稻营养和生殖生长的促进作用有所降低,导致混合种植方式下水稻和稗草生物量之比较27℃/21℃下降,水稻和稗草产量之比的降低幅度更大。该试验表明,仅提高CO2浓度条件下水稻相对于稗草的竞争力增强,然而CO2浓度和温度同时升高的条件下,稗草的竞争力更强[22]。本研究旨在前人研究的基础上,进一步明确大气CO2浓度升高条件下,稗草与水稻共生系统中稗草对水稻光合生理的影响,预估气候变化背景下C3作物与C4植物的竞争关系和相对优势,为水稻高产栽培及稗草的防除管理提供理论依据。
1 材料与方法
1.1 试验地点
试验于2019年和2020年在中国农业科学院昌平实验基地(40º08′N,116º08′E)进行,该地属暖温带半湿润大陆性季风气候,年平均日照时数2618h,年平均气温12.6℃,年平均降水量507.0mm。试验区土壤为褐潮土,土壤中含有机质14.10g⋅kg−1,全氮0.82g⋅kg−1,速效磷20.0mg⋅kg−1,速效钾79.8mg⋅kg−1,pH值为8.33[23]。
1.2 FACE系统简介
FACE系统主要由CO2气体供应装置、控制系统和CO2传感器构成,FACE圈由8根CO2气体释放管组成八边形,圈直径为4m,圈中心有自动监测设备,对温度、湿度和风速等气象指标进行实时监测。CO2浓度通过计算机模块自动控制,并根据具体风向和风速控制释放管电磁阀的开合度和方向,以实现预定浓度(550μmol·mol−1)目标并保持相对稳定。
1.3 试验设计
试验设置2个CO2浓度,分别为大气CO2浓度(400µmol·mol−1)和高浓度CO2(550µmol·mol−1),高浓度CO2环境应用FACE系统进行调控。每种CO2浓度处理中分别设2个稗草密度处理,分别为不种植稗草和按比例种植稗草,稗草与水稻株数比为1:5,即每5穴水稻间隔种1穴稗草,故试验共形成4组处理组合,以大气CO2浓度下不种植稗草的处理为对照(CK),在大气CO2浓度下种植稗草的处理组合设为TB,在高浓度CO2下不种植稗草处理组合设为TC,在高浓度CO2下种植稗草处理组合设为CB。
盆栽用土全部过5mm筛,试验用盆长60cm、宽40cm、高35cm,每盆装土50kg。每盆6穴,5穴水稻,每穴3株;1穴稗草,每穴1株,每组处理栽培12盆,自水稻和稗草移栽到FACE圈内,即开始向FACE圈内连续增施CO2气体,使圈内CO2浓度稳定在550µmol·mol−1,直到试验结束。移栽之初,每盆施加氮肥(尿素)20g,磷肥(磷酸二铵)20g,钾肥(氯化钾)10g,在抽穗期施加穗肥(尿素)10g,四组处理水分管理方式相同,始终保持灌溉水充足。
2019年水稻和稗草的移栽日期为6月24日,10月10日收获;2020年水稻和稗草的移栽日期为6月10日,10月2日收获。通过对水稻的田间观察,分别于水稻分蘖期、拔节期、抽穗期、成熟期取样测定相关指标。随水稻生长逐渐提高管道位置,使之始终高于水稻顶端20cm,且与地面相平行。试验用水稻品种为吉粳88,生育期140d左右,需≥10℃积温2900~3100℃·d,由中国农业科学院作物科学研究所提供。稗草种子采自辽宁沈阳。
1.4 项目测定
1.4.1 光合指标的测定
采用美国Li-Cor公司生产的便携式光合气体分析系统Li-6400,在水稻抽穗后每隔10d每个处理选取3穴有代表性的水稻,测定水稻剑叶光合参数,包括净光合速率(n)、气孔导度(s)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。并计算水分利用效率(WUE),WUE=Pn/Tr。测定时间为9:00−11:30。每个指标测定5个叶片,取平均值。
生来平等绝非是这个世界的运转规律,所以有些事物自诞生起就必须经历磨难也是命中注定的。虽然赢得了第二次世界大战,但重建工作的负担和生产资源的匮乏却让英国人继续保持了对未来的悲观态度。尽管还有不少盟军留下的吉普车可供利用,但英国人仍旧对这样的现状不太满意,而罗孚汽车公司工程主管在沙滩上的肆意之作却成就了一个拥有70年传奇的全地形SUV品牌—路虎。
采用日本产的SPAD-502 型叶绿素仪,测定水稻剑叶的叶绿素色度比值SPAD(soil and plant analyzer development)值。通过测量叶片对两个波长段里的吸收率,评估当前叶片叶绿素的相对含量。在水稻抽穗后每隔10d每个处理选取3穴有代表性的水稻选取水稻剑叶进行测定,同一张叶片上测定上、中、下3部分,取平均值。
1.4.2 干物质量的测定
在水稻的分蘖期、拔节期、抽穗期、成熟期每个处理分别取3穴水稻将植株连根带土放入40目尼龙网袋里,用水浸泡约30min,然后用流水仔细冲洗根系,用吸水纸擦干,随后放入牛皮纸袋中105℃杀青30min,维持在75℃,烘干至恒重,分别用0.01g JTS-600A电子天平测量每株水稻干重。
1.4.3 产量指标的测定
水稻成熟后,对各处理取植株形态相近、稻穗均匀的10穴水稻样本逐一考察,记录每穴穗数、穗粒数,并计算结实率。将成熟期水稻全部脱粒,从样本中随机数出3组各1000粒,风干后称重(精确到0.01g),得到千粒重。
1.5 数据分析
采用Excel进行数据处理及图表绘制,SPSS25.0(statistical package for the social science)进行分析处理,包括方差分析,采用一般线性模型双因素分析来统计CO2浓度和稗草的影响及其相互作用。ANOVA用于进行处理之间的差异显著性检验。各处理的比较采用最小显著差数(LSD)法,凡超过LSD0.05(或LSD0.01)水平的视为显著(或极显著),否则视为无显著性差异。
2 结果与分析
2.1 CO2浓度升高条件下稗草对水稻产量及其构成因素的影响
由表1可知,与对照CK相比,高CO2浓度条件下水稻产量提高了23.8%,影响效果达极显著水平,自然大气浓度状态下稗草与水稻混种使水稻产量降低了4.1%,也达到极显著水平;但CO2浓度升高与稗草互作对水稻产量影响并未达到显著水平。从水稻产量构成上分析,高CO2浓度使水稻每穴穗数增加了16.8%,差异达到显著水平,而稗草减少了水稻每穴穗数,且二者互作未达到显著水平。CO2浓度增高使水稻结实率提高了1.2%,而稗草使水稻的结实率减少了3.2%,但CO2浓度升高与稗草互作对水稻结实率影响并不显著。CO2浓度升高提高了水稻千粒重,但影响未达显著水平,同时稗草与水稻共生条件下,也使得水稻千粒重提高了3.1%,二者互作使千粒重提高了5.3%,影响达到了显著水平。
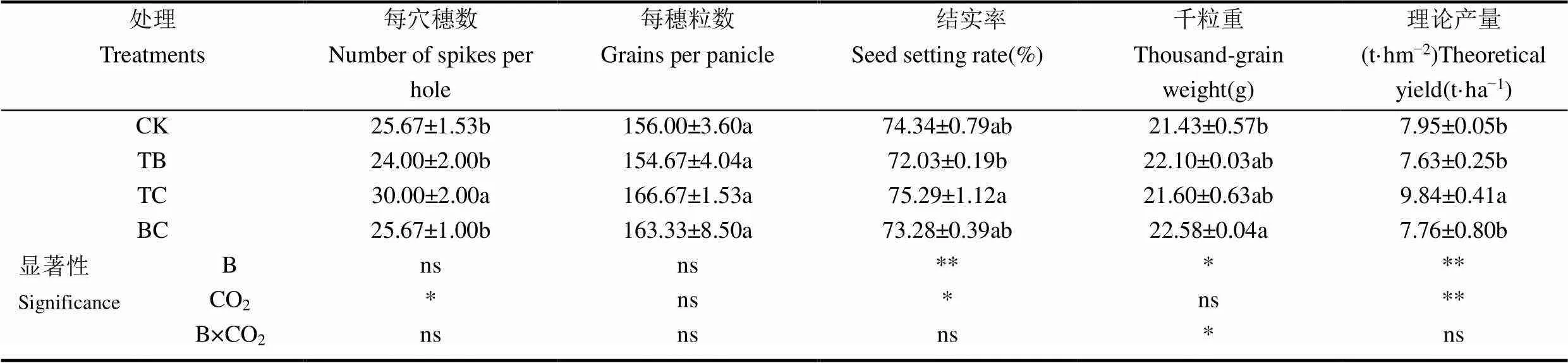
表1 不同处理水稻产量及其构成因素的比较
注:表中数值为平均值±标准差。CK为自然大气CO2(400µmol·mol−1)浓度条件下种植水稻,没有稗草;TB为自然大气CO2浓度条件下,水稻和稗草共生;TC为高CO2浓度(550µmol·mol−1)条件下种植水稻,没有稗草,BC为高CO2浓度条件下,水稻和稗草共生。小写字母表示处理间在0.05水平上的差异显著性。B指稗草。ns表示影响不显著,*和**分别表示在0.05和0.01水平上影响显著。下同。
Note: Data in the table are the mean ± standard deviation. CK: Atmospheric CO2(400µmol·mol−1) concentration and no barnyard grass; TB treatment: Atmospheric CO2concentration and planted barnyard grass with rice together; TC treatment: High concentration CO2(550µmol·mol−1) and no barnyard grass; BC treatment: High concentration CO2and planted barnyard grass with rice together. Lowercase indicates the difference significance among treatments at 0.05 level, B is barnyard grass. ns means that the influence of a factor is not significant,*and**indicate that the influence of a factor is significant at the level of 0.05 and 0.01, respectively. The same as below.
2.2 CO2浓度升高条件下稗草对水稻干物质积累的影响
对水稻干物质影响分析发现,在自然大气CO2水平下水稻中混种稗草对单株干物质积累量没有明显不利影响,甚至可能还有一定的促进作用,而单种水稻情况下CO2浓度升高对干物质积累具有明显的促进作用,此时混种稗草后完全抵消了CO2浓度升高产生的肥效作用。成熟期与对照(CK)相比,自然大气浓度状态下稗草与水稻混种(TB)减少了水稻干物质量,在抽穗期和成熟期分别减少了38.8%和6.4%;而CO2浓度升高处理(TC)增加了水稻干物质量,在水稻抽穗期和成熟期分别提高了100.9%和88.4%。CO2浓度升高与稗草互作(BC)使水稻干物质量在各发育期差异不显著。
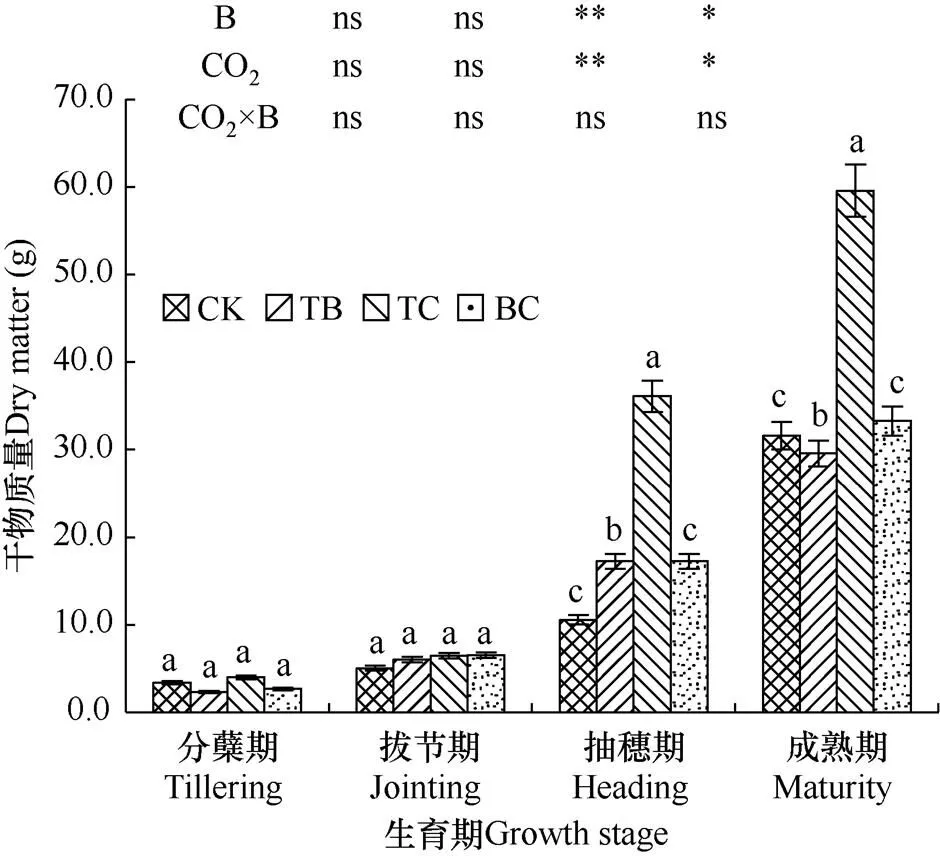
图1 不同处理水稻各生育期单株干物质量比较
注:短线表示均方差。下同。
Note:The bar is mean square error. The same as below.
2.3 CO2浓度升高条件下稗草对水稻叶绿素含量的影响
叶绿素相对含量(SPAD)表示作物叶片绿色程度,是预测作物产量的指标之一,一般水稻抽穗开花期叶片SPAD值与产量显著相关。由图2可知,与对照(CK)相比,自然大气浓度状态下稗草与水稻混种(TB)降低了水稻SPAD值,在抽穗后10d、20d、30d内分别降低了4.7%、6.3%和3.2%,影响达到显著水平,且在水稻抽穗后第10天和第30天达到极显著水平。而CO2浓度升高处理(TC)提高了水稻SPAD值,抽穗后10d、20d、30d内分别比对照提高了11%、7.2%和1.3%,在抽穗后20d内浓度升高对剑叶SPAD值的影响达到极显著水平。高CO2浓度与稗草互作(BC)使水稻剑叶SPAD值在抽穗后第10天和第20天分别增加了8.7%和9.1%,且在抽穗后20d的影响达到极显著水平。而在抽穗后第30天基本持平。结果表明CO2浓度升高在水稻抽穗前期对水稻SPAD值影响较大,但随着抽穗期光合速率增加,叶绿素相对含量减少,CO2浓度升高对水稻SPAD值影响逐渐减小。
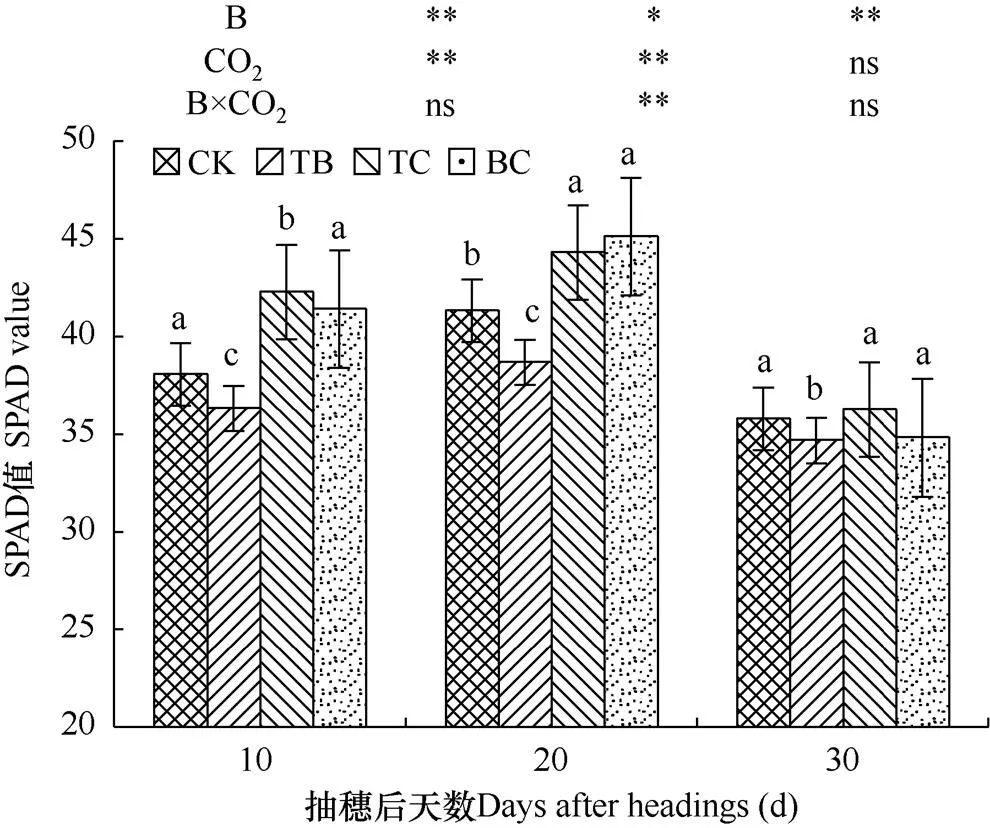
图2 不同处理水稻剑叶SPAD值的比较
2.4 CO2浓度升高条件下稗草对水稻生理指标的影响
2.4.1 对净光合速率的影响
净光合速率是作物重要的光合特征参数之一,其大小可以反映作物同化CO2的能力。由图3可见,与对照处理(CK)相比,自然大气浓度状态下稗草与水稻混种(TB)降低了水稻剑叶光合速率,在水稻抽穗后10d、20d和30d内分别降低了16.3%、8.1%和6.5%,稗草对水稻剑叶净光合速率的影响达到极显著水平。而CO2浓度增高处理(TC)增加了水稻剑叶光合速率,在水稻抽穗后10d、20d和30d内分别增加了8.0%、38.5%和65.1%,影响均达到极显著水平。高CO2浓度与稗草互作(BC)使水稻剑叶净光合速率在水稻抽穗后10d内降低了4.1%,在抽穗后20d和30d内分别增加11.7%、42.7%,影响均达到极显著水平。
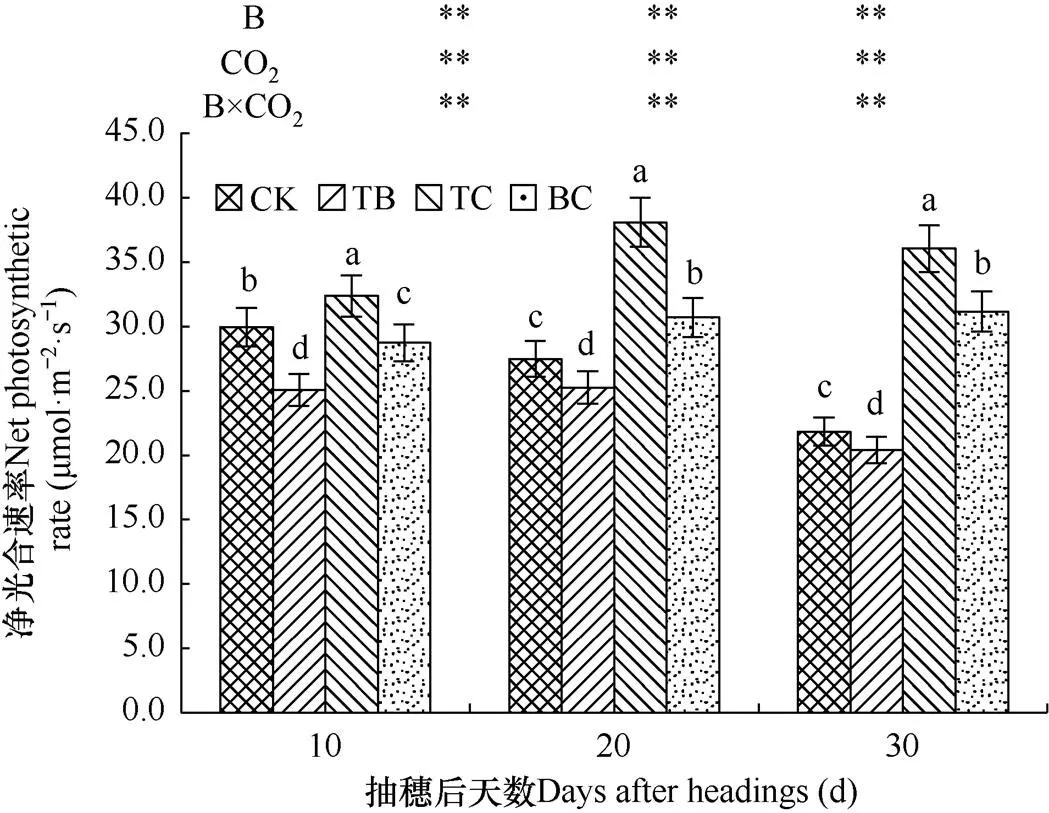
图3 不同处理水稻剑叶光合速率的比较
对水稻剑叶光合速率的分析表明,在水稻抽穗后,随着水稻株高及叶面积的增加,稗草对水稻剑叶净光合速率的影响逐渐减小,而水稻剑叶净光合速率随着CO2浓度提高显著增加,在水稻抽穗后期对水稻剑叶净光合速率影响更大。所以高浓度CO2与稗草互作在水稻抽穗后期使水稻剑叶净光合速率显著提高。
2.4.2 对气孔导度的影响
气孔导度表示气孔的开放程度,是表征作物光合、呼吸及蒸腾的主要指标。由图4可见,与对照处理(CK)相比,水稻抽穗后,自然大气浓度状态下,稗草与水稻混种(TB)和CO2浓度升高处理(TC)均降低了水稻剑叶气孔导度,前者在水稻抽穗后10d、20d和30d内分别降低40.7%、10.1%和7.5%,后者分别降低36.4%、7.8%和52.3%。且高浓度CO2处理的影响在水稻抽穗后达到极显著水平,稗草处理在水稻抽穗后20d对气孔导度的影响达到极显著水平,在水稻抽穗30d时达到显著水平。CO2浓度升高与稗草互作(BC)使水稻剑叶气孔导度在抽穗后10d、20d和30d内分别降低47.3 %、25.7%和56.0%,均达到极显著水平。
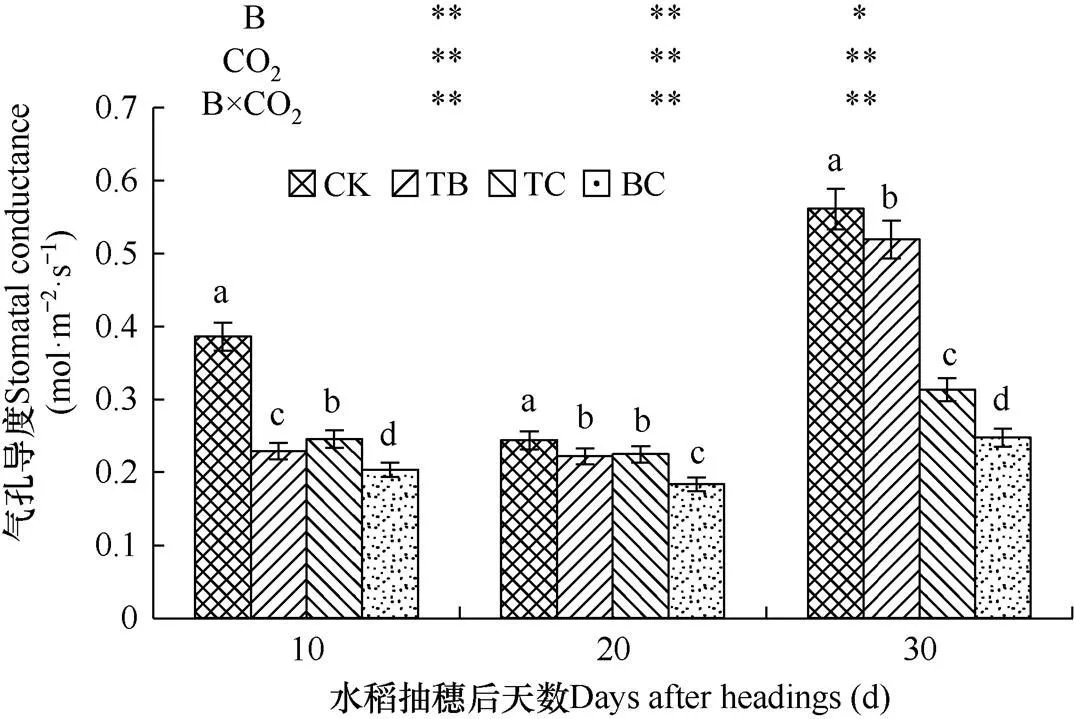
图4 不同处理水稻剑叶气孔导度的比较
2.4.3 对蒸腾速率的影响
蒸腾速率是一定时间内土壤水分通过植物叶片气孔向大气输送的水量,是反映植物水分代谢能力的重要指标。由图5可见,与对照处理(CK)相比,自然大气浓度状态下稗草与水稻混种(TB)降低了水稻剑叶蒸腾速率,在水稻抽穗后10d、20d和30d内分别降低了21.0%、5.4%和9.7%,且对水稻剑叶蒸腾速率的影响达到极显著水平;而CO2浓度增高处理(TC)降低了水稻剑叶蒸腾速率,在水稻抽穗后10d、20d和30d内分别降低了13.0%、5.2%和44.0%,对水稻剑叶蒸腾速率的影响在水稻抽穗后达到极显著水平。CO2浓度升高与稗草互作处理(BC)的水稻剑叶蒸腾速率,在水稻抽穗后10d、20d和30d内分别降低了26.2%、16.3%和54.4%,说明CO2浓度升高对水稻剑叶蒸腾速率的影响在水稻抽穗后达到极显著水平。
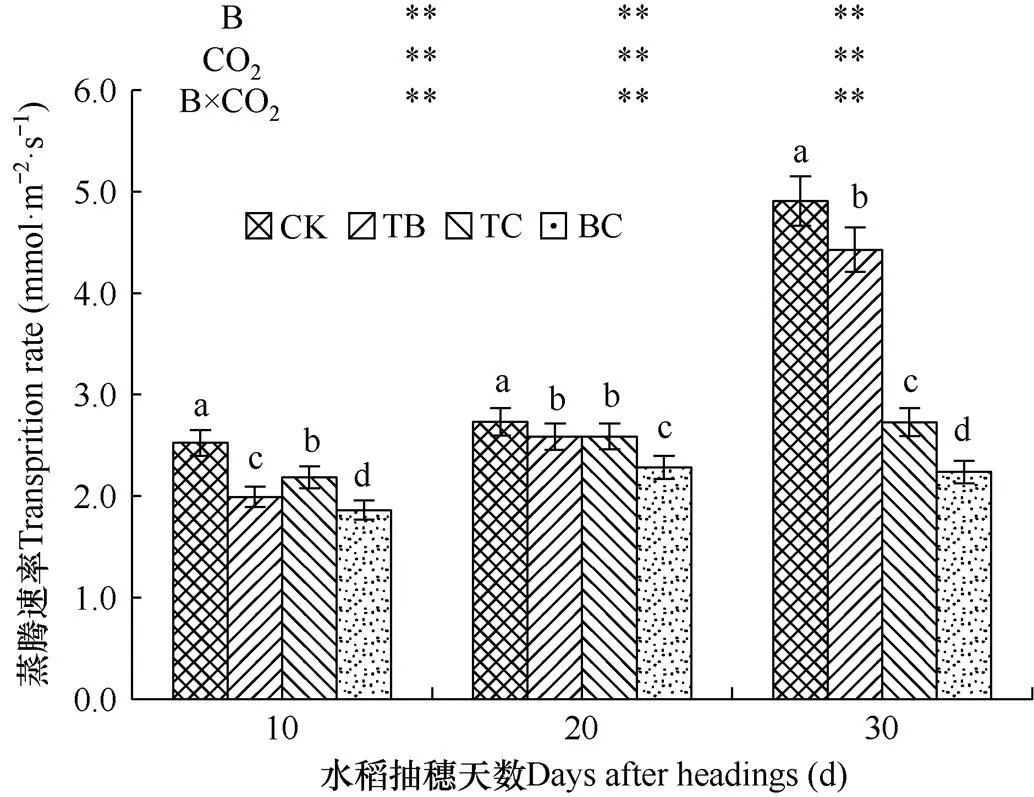
图5 不同处理水稻剑叶蒸腾速率的比较
2.4.4 对剑叶胞间CO2浓度的影响
剑叶胞间CO2浓度是反映水稻细胞光合利用、光呼吸的CO2动态平衡瞬间浓度的指标。由图6可见,与对照处理(CK)相比,自然大气浓度状态下稗草与水稻混种(TB)降低了水稻剑叶胞间CO2浓度,在抽穗后10d、20d和30d内分别降低了2.5%、1.5%和5.9%,而CO2浓度升高处理(TC)增加了水稻剑叶胞间CO2浓度,在水稻抽穗后10d、20d和30d内分别提高了110.1%、14.8%和12.7%,CO2浓度升高与稗草互作处理(BC)分别提高了47.2%、11.0%和5.4%。方差分析表明,尽管影响的效果相反,但是稗草处理、高浓度CO2处理以及高浓度CO2与稗草互作在水稻抽穗后,对水稻剑叶胞间CO2浓度的影响均达到极显著水平。当然,CO2浓度升高对水稻剑叶胞间CO2浓度影响程度大于稗草的影响程度。
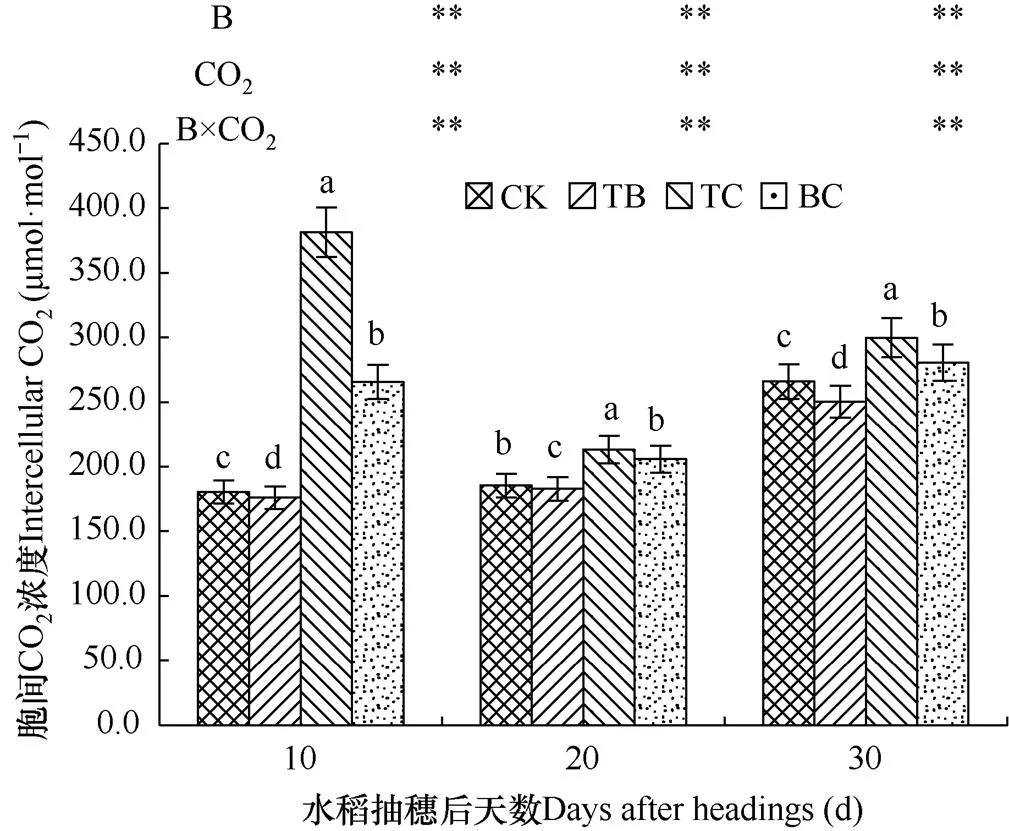
图6 不同处理水稻剑叶胞间CO2浓度的比较
2.4.5 对水分利用率的影响
水分利用率是作物在消耗等量水分情况下固定CO2的能力,反映了作物在生产中能量转化的效率,是作物叶片水分利用特征的基本生理参数。由图7可知,自然大气浓度状态下稗草与水稻混种(TB)降低了水稻水分利用率,在抽穗后10d、20d和30d内分别降低了21.7%、3.0%和15.6%,对水稻水分利用率的影响达到极显著水平。而CO2浓度升高处理(TC)在抽穗后10d内降低了7.9%,在抽穗后20d和30d内分别增加了46.5%和226.6%,对水稻水分利用率的影响达到极显著水平。CO2浓度升高与稗草互作(BC)使水分利用率较在水稻抽穗后10d内降低了29.2%,在水稻抽穗后20d和30d内分别增加了33.6%、136.7%,在水稻抽穗20d后对水稻水分利用率的影响达到极显著水平。
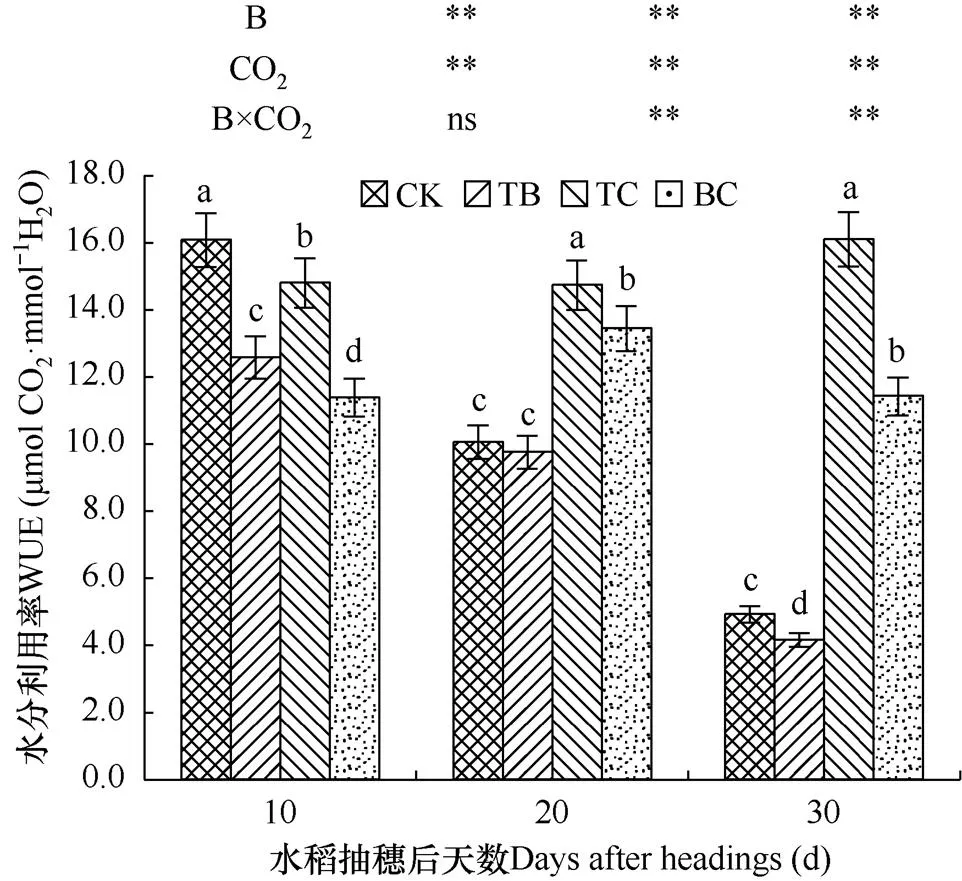
图7 不同处理水稻剑叶水分利用率比较
3 结论与讨论
3.1 讨论
本研究把稗草作为与CO2相似的一个外部影响因子,探索其对水稻光和生理及生长发育的影响。基本假设是CO2的作用是正向的,而稗草的作用是负向的,二者的交互作用是未知的。旨在通过本研究明确二者的影响效果,试图揭示基本机理,对未来气候变化背景下水稻生产管理有什么启示,以及如何应对不同的影响效果。试验结果与之前的假设基本吻合,但是结果更为复杂,需要更详尽的分析和讨论。
CO2浓度的增加促进了水稻光合作用,有利于水稻干物质形成,从而提高了水稻产量形成[24],从水稻产量构成上分析,CO2浓度增高提高了水稻分蘖数和每穴穗数,是导致水稻产量增加的主要原因[25]。本试验结果显示,高浓度CO2生长环境下水稻穗数平均增加16.8%,是导致水稻增产的主要因素。之前的研究同样表明水稻穗数增加对水稻产量增加的贡献最大[26−27]。进一步分析发现,CO2浓度增高,使水稻每穗粒数、千粒重、结实率都有一定幅度的促进作用,不过增幅小于其对水稻穗数的影响,对于提高产量作用较小。水稻进入拔节期时,稗草生长速度迅速增加,逐渐超过水稻,形成遮荫,使水稻群体透光率降低,从而影响水稻光合作用和光合产物输送,最终使水稻结实率、千粒重都有所降低,影响水稻产量形成。本试验结果表明,CO2浓度升高与稗草互作使水稻千粒重增加,但对水稻产量影响未达到显著水平,可见是稗草起到了干扰的作用。
本研究表明,自然大气浓度状态下稗草与水稻混种促使水稻净光合速率、叶片SPAD值、蒸腾速率都有所下降,其中净光合速率降幅最大,这些变化使水稻水分利用率降低,减少了水稻干物质的形成。叶片通过叶绿素进行光合作用,叶片叶绿素含量与植物光合作用紧密相关,CO2浓度升高促进叶绿素合成,提高了SPAD值。由于叶片光和能力和光和效率与SPAD值密切相关。稗草生长旺盛,对水稻形成遮阴导致对光能的捕获减少,从而影响叶绿素的合成,使得SPAD值降低,加之稗草的空间优势,进一步弱化了水稻光合作用,有研究表明水稻叶片SPAD值与水稻产量存在直接的正相关关系,随着叶片SPAD值的增大,产量也呈增加趋势[28−29]。中间的一系列生理过程和指标,包括水稻净光合速率、蒸腾速率、呼吸速率等自然影响着产量的最终形成[13,17]。
光合作用是植物体吸收光能,转化为生长所需的化学能的过程,是向生物提供碳水化合物以积累干物质的生化过程。CO2作为光合作用底物,其浓度升高直接影响水稻的生理生化过程,包括光合作用、呼吸作用和气孔导度以及地上地下生物量的形成。高浓度CO2引起叶片气孔关闭,造成气孔导度下降,使CO2进入叶肉细胞的阻力增大,但大气 CO2浓度的升高,使气孔内外CO2浓度梯度增加,细胞间隙CO2浓度相应升高,因此提高了叶肉细胞的光合速率。所以大气CO2浓度增高导致的部分气孔关闭,并不会抑制作物光合作用,短时间内随着CO2浓度的升高,作物的光合速率还是有所提高。CO2浓度增加还会使1,5−二磷酸核酮糖羧化酶(Rubisco) 活性增强,羧化速率增加,抑制C3作物光呼吸,使C3作物净光合速率显著提高[12, 30]。
高浓度CO2主要通过气孔导度、叶面积和冠层温度三者共同作用来影响蒸腾速率,即高CO2浓度导致作物叶片气孔的开张度缩小或关闭,气孔导度降低,阻力升高,蒸腾速率减小[31−33]。本研究发现CO2浓度越高,水稻光合速率和胞间CO2浓度越增加,而气孔导度和蒸腾速率越减少。而CO2浓度升高使水稻净光合速率提高,蒸腾速率降低导致水稻剑叶水分利用率显著提高,从而使水稻干物质量显著增加。水稻进入穗分化期后,稗草生长迅速,株高逐渐超过水稻,株高在水稻和稗草共生中发挥着重要作用,相对较高的植物更易捕获光照,促使其选择性吸收红光,反射远红光,导致相对较矮植物吸收远红光量增加,致使红光与远红光吸收比率降低。红光和远红光吸收比率的降低将引起植物一系列的生理反应,如气孔的发育与闭合、光敏色素A和光敏色素B间的转变,光敏色素B的变化将影响气孔发育和闭合,以及乙烯释放速率增加等[34−35]。而且由于稗草高于水稻,使水稻群体透光率降低,从而影响水稻光合。
3.2 结论
高CO2浓度条件下水稻每穴穗数增加了16.8%,结实率提高1.2%~1.7%,最终使水稻产量增加17.7%~23.8%;稗草与水稻共生条件下水稻结实率降低2.7%~3.2%,千粒重增加3.1%~4.5%,最终使水稻产量降低4.1%~21.2%。CO2浓度升高与稗草互作对水稻产量影响并不显著。高CO2浓度促进了水稻剑叶净光合速率、胞间CO2浓度及SPAD值的升高,使水稻剑叶气孔导度和蒸腾速率降低,稗草与水稻共生降低了水稻剑叶净光合速率、胞间CO2浓度、气孔导度、蒸腾速率、瞬时水分利用效率及SPAD值;高CO2浓度与稗草互作使水稻剑叶净光合速率先降低后升高,水稻抽穗后CO2浓度升高对水稻光合作用的影响大于稗草对水稻光合作用的影响。所以稻田稗草的危害主要在前期,管理的重点也应在水稻生长前期。
[1] IPCC.Climate change 2021:the scientific basis,contribution of working group Ⅰto the sixth assessment report of intergovernmental panel on climate change (IPCC) [M]. Cambridge:Cambridge University Press,2021.
[2] Idso K E,Idso S B.Plant responses to atmospheric CO2enrichment in the face of environmental constraints:a review of the past 10 years' research[J].Agricultural and Forest Meteorology,1994,69(3-4):153-203.
[3] Maria V,Martha T,Stella L,et al.Assessing the impact of increasing carbon dioxide and temperature on crop-weed interactions for tomato and a C3 and C4 weed species[J].European Journal of Agronomy,2013(50):60-65.
[4] 谢立勇,林而达,高西宁,等.二氧化碳浓度增高对作物影响研究方法问题与讨论[J].中国农业大学学报,2008,13 (3): 23-26.
Xie L Y,Lin E D,Gao X N,et al.FACE and its limitations on researches of impacts of elevated CO2 on crops[J].Journal of China Agricultural University,2008,13(3):23-28.
[5] 陈国奇,唐伟,李俊,等.我国水稻田稗属杂草种类分布特点:以9个省级行政区73个样点调查数据为例[J].中国水稻科学,2019,33(4):368-376.
Chen G Q,Tang W,Li J,et al.Distribution characteristics of echinocloa species in rice fields in China: a case survey on 73 sites from nine provincial administrative regions[J]. Chin J Rice Sci,2019,33(4):368-376.(in Chinese)
[6] Chauhan,Abugho.Effects of water regime,nitrogen fertilization,and rice plant density on growth and reproduction of lowland weed Echinochloa crus-galli[J]. Crop Protection,2013(54):142-147.
[7] 林文雄,何华勤,郭玉春,等.水稻化感作用及其生理生化特征的研究[J].应用生态学报,2001,12(6):871-875.
Lin W X,He H Q,Guo Y C,et al.Rice allelopathy and its physiobiochemical characteristics[J].Chinese Journal of Applied Ecology,2001,12(6):871-875.(in Chinese)
[8] Zhang H,Liu H,Hou D,et al.The effect of integrative crop management on root growth and methane emission of paddy rice[J].The Crop Journal,2019,7(4):444-457.
[9] Yang J C,Zhang H,Zhang J H.Root morphology and physiology in relation to the yield formation of rice[J].Journal of Integrative Agriculture,2012,11(6):920- 926.
[10] 付景,徐云姬,陈露,等.超级稻花后强、弱势粒淀粉合成相关酶活性和激素含量变化及其与籽粒灌浆的关系[J].中国水稻科学,2012,26(3):302-310.
Fu J,Xu Y J,Chen L,et al.Postanthesis changes in activities of enzymes related to starch synthesis and contents of hormones in superior and inferior spikelets and their relation with grain filling of super rice[J].Chin J Rice Sci.2012,26(3):302-310.(in Chinese)
[11] Wang X L,Zhang Z Y,Xu X M,et al.The density of barnyard grass affects photosynthesis and physiological characteristics of rice[J].Photosynt,2019(57):705-711.
[12] Xie L Y,Lin E D,Zhao H L,et al.Changes in the activities of starch metabolism enzymes in rice grains in response to elevated CO2concentration[J].International Journal of Biometeorology,2016,60(5):727-736.
[13] 李书燕,王彦辉,柏连阳,等.不同密度下抗性与敏感稗草对水稻氮素水平和光合作用的影响[J].植物生理学报,2020,56(12):2677-2682.
Li S Y,Wang Y H,Bai L Y,et al.Effects of different densities of resistant and sensitive barnyardgrass on nitrogen content and the photosynthesis in rice[J].Plant Physiology Journal, 2020,56(12):2677-2682.(in Chinese)
[14] 牛玺朝,户少武,杨阳,等.大气CO2浓度增高对不同水稻品种稻米品质的影响[J].中国生态农业学报,2021, 29(3):509-519.
Niu X C,Hu S W,Yang Y,et al.Effects of CO2concentration enrichment on the grain quality of different rice varieties[J].Chinese Journal of Eco-Agriculture,2021,29(3): 509-519.(in Chinese)
[15] 曾青,朱建国,刘刚,等.开放式空气CO2浓度增高条件下C3作物(水稻)与C4杂草(稗草)的竞争关系[J].应用生态学报,2002(10):1231-1234.
Zeng Q,Zhu J G,Liu G,et al.Effect of FACE on competition between a C3 crop (rice) and a C4 weed (barnyard grass Echinochloa crusgalli)[J].Chinese Journal of Applied Ecology,2002(10):1231-1234.(in Chinese)
[16] Ziska L H.The impact of elevated CO2on yield loss from a C3 and C4 weed in field -grown soybean[J].Global Change Biology,2000(6):899-905.
[17] 张自常,谷涛,杨霞,等.不同抗性稗草对水稻产量及其生理特性的影响[J].核农学报,2017,31(8):1594-1603.
Zhang Z C,Gu T,Yang X,et al.Effects of barnyard grass with different resistance on grain yield of rice and their physiological characteristics[J].Journal of Nuclear Agricultural Sciences,2017,31(8):1594-1603.(in Chinese)
[18] 张自常,李永丰,杨霞,等.干湿交替灌溉条件下不同种稗草对水稻光合特性和产量的影响[J].作物学报,2015, 41(11):1748-1757.
Zhang Z C,Li Y F,Yang X,et al.Effects of different species in echinochloa on photosynthetic characteristics and grain yield in rice under alternate wetting and moderate drying condition[J].Acta Agronomica Sinica,2015,41(11):1748- 1757.(in Chinese)
[19] 梁国华,汤瑞琼,李明霞.水稻—稗草在共培条件下的相互影响研究[J].种子科技,2019,37(10):164-165.
Liang G H,Tang R Q,Li M X.Interaction of rice and barnyard grass under co-culture[J].Seed Science and Technology, 2019,37(10):164-165.(in Chinese)
[20] 孙富芝,朱建国,曾青,等.大气CO2浓度升高条件下移栽水稻与不同出苗时间稗草竞争的响应差异[J].中国水稻科学,2007(2):203-208.
Sun F Z,Zhu J G,Zeng Q,et al.Difference of competition responses between rice and barnyard grass with different seedling emergence time under elevated atmosphere CO2[J].Chinese J Rice Sci,2007(2):203-208.(in Chinese)
[21] Alberto M P,Ziska L H,Cervancia C R,et al.The Influence of increasing carbon dioxide and temperature on competitive interactions between a C3 crop, rice () and a C4 weed (Echinochloa glabrescens)[J]. Functional Plant Biology, 1996, 23(6): 795-802.
[22] Zhang Z,Gu T,Zhao B,et al.Effects of common Echinochloa varieties on grain yield and grain quality of rice[J].Field Crops Research,2017(203):163-172.
[23] 张凤哲,谢立勇,赵洪亮,等.大气CO2浓度升高与施加生物炭对水稻的影响分析[J].中国农业气象,2021,42(4): 287-296.
Zhang F Z,Xie L Y,Zhao H L,et al.Interactive effects of high CO2concentration and biochar addition on root system and yield of rice[J].Chinese Journal of Agrometeorology,2021,42(4):287-296.(in Chinese)
[24] Elizabeth A,Ainsworth H.Rice production in a changing climate:a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration[J].Global Change Biology,2008,14(7):1642-1650.
[25] Kim H Y,Lieffering M,Kobayashi K,et al.Effects of free-air CO2enrichment and nitrogen supply on yield of temperate paddy rice crops[J].Field Crops Research, 2003(83):261-270.
[26] 户少武,张欣,景立权,等.高浓度CO2对稻穗不同位置籽粒结实和米质性状的影响[J].应用生态学报,2019,30(11): 3725-3734.
Hu S W,Zhang X,Jiang L Q,et al.Effects of elevated CO2concentration on grain filling capacity and quality of rice grains loca-ted at different positions on a panicle[J]. Chinese Journal of Applied Ecology,2019,30(11):3725- 3734.(in Chinese)
[27] 王惠贞,赵洪亮,冯永祥,等.北方水稻生育后期剑叶可溶性物质含量及植株生产力对CO2浓度增高的响应[J].作物学报,2014,40(2):320-328.
Wang H Z,Zhao H L,Feng Y X,et al.Response of soluble substances content in flag leaves during late growth stage and plant productivity of rice to elevated CO2in North China[J].Acta Agronomica Sinica,2014,40(2):320-328.(in Chinese)
[28] 黄影华,张华杰,陈秋玉,等.不同生育期水稻叶片SPAD值与产量相关性研究[J].仲恺农业工程学院学报,2021, 34(1):1-7.
Huang Y H,Zhang H J,Chen Q Y,et al.Correlation between SPAD values of rice leaf and yield among different growth stages[J].Journal of Zhongkai University of Agriculture and Engineering,2021,34(1):1-7.(in Chinese)
[29] 欧阳杰,王楚桃,何光华,等.水稻灌浆中后期功能叶中叶绿素含量及其变化趋势与谷物产量关系研究[J].西南农业学报,2012,25(4):1201-1204.
Ouyang J,Wang C T,He G H,et al.Study on relationship between different functional leaf chlorophyll content and its trends in mid and late period of rice filling and grain yield[J].Southwest China Journal of Agricultural Sciences, 2012(4): 1201-1204.(in Chinese)
[30] Sage R F,Kubien D S.The temperature response of C3 and C4 photosynthesis[J].Plant,Cell & Environment,2007,30(9): 1086-1106.
[31] 邵在胜,赵轶鹏,宋琪玲,等.大气CO2和O3升高对水稻汕优63叶片光合作用的影响[J].中国生态农业学报,2014, 22(4):422-429.
Shao Z S,Zhao Y P,Song Q L,et al.Impact of elevated atmospheric carbon dioxide and ozone concentrations on leaf photosynthesis of ‘Shanyou 63’ hybrid rice[J].Chinese Journal of Eco-Agriculture,2014,22(4):422-429.(in Chinese)
[32] 杨连新,王云霞,朱建国,等.开放式空气中CO2浓度增高(FACE)对水稻生长和发育的影响[J].生态学报,2010, 30(6):1573-1585.
Yang L X,Wang Y X,Zhu J G,et al.What have we learned from 10 years of free-air CO2enrichment(FACE) experiments on rice? Growth and development[J].Acta Ecologica Sinica,2010,30(6):1573-1585.(in Chinese)
[33] Jeffrey B L,Hartwell A,Kenneth B,et al.Rice responses to drought under carbon dioxide enrichment 2:photosynthesis and evapotranspiration[J].Global Change Biology,1997, 3(2):129-138.
[34] Afifi M,Swanton C.Maize seed and stem roots differ in response to neighbouring weeds[J].Weed Research,2011, 51(5):442-450.
[35] Boccalandro H E,Rugnone M L,Moreno J E,et al.Phytochrome B enhances photosynthesis at the expense of water-use efficiency in arabidopsis[J].Plant Physiology, 2009,150(2):1083-1092.
Impacts of Barnyard Grass on Photosynthesis and Physiology of Rice under Elevated Atmospheric CO2Concentration
JIN Dian-yu1, XIE Li-yong1, ZHAO Hong-liang1, LI Ying1, HAN Xue2, HE Yu-tong1, LIN Er-da2
(1.College of Agronomy, Shenyang Agricultural University, Shenyang 110161, China; 2. Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081)
Barnyard grass is one of the vicious weeds in rice field, which affects seriously the growth and development of rice and reduces the yield of rice. In order to clarify the effect of barnyard grass on rice growth and development and yield formation under the condition of elevated atmospheric carbon dioxide (CO2) concentration, the experiment was conducted with the Free-Air CO2Enrichment system (FACE system), taking rice Jijing 88 as material. The experiment included 4 treatments: atmospheric CO2concentration (400µmol·mol−1) without barnyard grass, rice planting and barnyard grass together with atmospheric CO2concentration, elevated CO2concentration (550µmol·mol−1) without barnyard grass, rice planting and barnyard grass together with elevated CO2concentration. Physiological indicators were measured and analyzed at the tillering stage, jointing stage, heading stage, and mature stage of rice. The results showed that elevated CO2concentration increased significantly the number of rice ears per hole and the seed setting rate, and eventually increased the rice yield. Barnyard grass significantly reduced the rice seed setting rate, and reduced rice yield ultimately. Interaction of elevated CO2concentration and barnyard grass increased significantly thousand-grain weight of rice, yet, the interaction did not affected rice yield significantly. Elevated CO2concentration increased significantly the dry matter of rice, and barnyard grass reduced significantly the dry matter of rice. However, the interaction of elevated CO2concentration and the barnyard grass had no significant difference. Elevated CO2concentration increased significantly the net photosynthetic rate of the flag leaf of rice, the concentration of intercellular CO2of the flag leaf, and the Soil and Plant Analyzer Developrnent (SPAD) value of rice flag leaf. Barnyard grass treatment reduced significantly the net photosynthetic rate of the flag leaf of rice, the concentration of intercellular CO2in the flag leaf, the stomatal conductance, the transpiration rate, the instantaneous water use efficiency, and the SPAD value. The interaction of elevated CO2concentration and barnyard grass reduced the net photosynthetic rate of rice flag leaves at early stage, and increased it at later stage. In conclusion, Elevated CO2concentration after heading stage had a greater impact on rice photosynthesis than barnyard grass on rice photosynthesis.
Climate change;Carbon dioxide;Barnyard grass;Rice;Photosynthesis
金殿玉,谢立勇,赵洪亮,等.大气CO2浓度升高条件下稻稗共生系统中稗草对水稻光合生理的影响[J].中国农业气象,2022,43(3):204-214
10.3969/j.issn.1000-6362.2022.03.004
2021−06−30
国家自然科学基金(41875141;41175097)
谢立勇,教授,研究方向为气候变化与低碳农业,E-mail:xly0910@163.com
金殿玉,E-mail:2263460190@qq.com
猜你喜欢
中国水稻科学(2021年6期)2021-11-18 12:06:30
节水灌溉(2021年5期)2021-05-26 06:49:38
中国农业科技导报(2020年12期)2020-03-15 10:50:08
浙江农业科学(2019年1期)2019-01-25 05:25:34
新课程·下旬(2017年7期)2017-08-14 11:12:50
扬子江(2016年1期)2016-05-19 22:06:26
西北林学院学报(2015年5期)2015-01-03 05:12:20
东北农业大学学报(2014年1期)2014-01-14 09:00:42
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
植物营养与肥料学报(2011年2期)2011-10-26 03:52:46
