
水稻剑叶形态与单株产量的基因定位分析
2014-01-14 09:00:42邹德堂王晋王敬国刘化龙刘宇强贾琰
东北农业大学学报 2014年1期
邹德堂,王晋,王敬国,刘化龙,刘宇强,贾琰
(东北农业大学农学院,哈尔滨 150030)
水稻剑叶形态与单株产量的基因定位分析
邹德堂,王晋,王敬国,刘化龙,刘宇强,贾琰
(东北农业大学农学院,哈尔滨 150030)
以剑叶形态和单株穗重差异较大的亲本小白粳子和空育131以及其后代F3群体为试验材料,构建包含99个标记的遗传连锁图谱,对剑叶长度、剑叶宽度、剑叶面积、剑叶基角、剑叶张角和单株穗重进行相关性分析和QTL定位研究。在水稻第1、3、4、5、6、7、9条染色体上共检测到17个QTL。其中控制剑叶长度的QTL 3个,控制剑叶宽度的QTL 4个,控制剑叶面积的QTL 2个,控制剑叶基角的QTL 3个,控制剑叶张角的QTL 2个,控制单株穗重的QTL 3个。为水稻剑叶形态、理想株型的改良和分子辅助选择育种提供理论依据。
水稻;剑叶形态;单株穗重;数量性状基因座位
水稻(Oryza sativa)原产于中国,是重要的粮食作物,传统水稻遗传育种中,把提高水稻产量作为首要育种目标,在生产实践中发现,理想株型和水稻高产育种密切相关[1]。
叶片是水稻制造有机养料的重要器官,也是水稻进行光合作用重要场所。以水稻叶片为代表的“源”单位和以单株穗重为代表的“库”单位有紧密联系。董国军等认为,剑叶夹角是构成水稻株型的主要因素之一,减少剑叶角度有利于增加水稻的光合作用,提高产量,促使不育系不易授粉,杂交种的繁殖能力下降[2]。
我国对水稻剑叶形态的研究起步较早,早在20世纪80年代初期,沈福成提出在水稻高光效育种中,着重对水稻剑叶的长度、宽度、抽穗期剑叶与茎秆的夹角和比叶重进行遗传研究[3]。随着分子生物学的快速发展,学者们开始关注水稻剑叶各农艺性状诸如剑叶长、宽、面积和许多生理性状的基因定位和克隆研究[4]。蔡晶利用两个DH群体在两个环境下运用区间作图的方法检测到控制剑叶长的QTL 13个,检测到8个控制剑叶宽度的QTL,检测到控制剑叶基角的QTL共9个[5]。沈波等利用247个株系、158个标记的重组自交系群体对叶片长度、宽度、周长和长宽比进行基因定位,共在9个区间检测到24个具有加性效应的QTL,其中贡献率最高的达到32.5%,同时该研究表明,除加性效应以外,上位效应也对群体的叶片形态的遗传控制起重要作用[6]。童汉华等对水稻生育后期剑叶的形态和生理进行QTL定位研究发现,在染色体的特定区段内存在控制多个数量性状的基因位点,这也是一因多效性的重要体现[7]。研究者大多对产量及产量相关性状进行基因定位研究,但对剑叶形态指标研究相对较少,本试验以剑叶和产量性状差别较大的品种小白粳子和空育131为亲本的杂交后代群体为材料,利用分子标记技术对水稻剑叶长、剑叶宽、剑叶面积、剑叶基角、剑叶张角、单株穗重进行QTL定位分析,揭示剑叶性状和产量性状关系,为理想株型的选育和分子辅助选择提供依据。
1 材料与方法
1.1 试验材料
选用剑叶和产量性状相差较大的小白粳子和空育131为亲本,经F1自交后得到F2,再将F2群体自交得到229个单穗,进一步获得的F2∶3家系为本试验的供试群体。
1.2 试验方法
2011年将试验材料种植于东北农业大学香坊实验实习基地,采用旱育苗方式,4月11日浸种,4月16日播种,5月26日移栽。随机区组设计,两行区,5 m行长,株距10 cm,行距30 cm,每穴插单株。水稻生长至齐穗期后,在两行区内随机取5株挂牌作为5次重复,测量这5个单株的所有分蘖剑叶性状和穗重并取平均数作为试验结果。主要测量指标包括剑叶长度(FLL)、剑叶宽度(FLW)、剑叶面积(FLA)、剑叶基角(FLBA)、剑叶张角(FLSA)和单株产量(EW)。其中剑叶长和宽直接用直尺测量,以cm为单位;剑叶面积用叶面积仪测量;剑叶基角指叶片基部与茎秆的夹角,剑叶张角指叶尖到叶耳连线与茎秆的夹角,使用半径35 cm的量角器测量;待水稻完全成熟后,将一株水稻的各穗沿穗颈节处剪下,用天平称量一株所有穗的总重量,并求出5个单株的平均数,以g为单位。
1.3 图谱构建
根据F2群体中各个标记的多态性信息,通过QTLIciMapping 3.0进行连锁分析并构建遗传连锁图谱,使用Kosambi函数,将重组率转换成图距单位(cM),采用MapChart 2.2版本绘制连锁图谱。将筛选出来的99个标记划分为12个连锁群,根据标记间的距离和顺序绘制一个包含99个SSR标记的遗传连锁图谱。该遗传图谱的总长度为1 238.42 cM,最远的两个标记之间距离为42.38 cM,最近距离为0.46 cM,平均距离12.51 cM。
1.4 数据处理
采用DPS软件对水稻单株穗重、剑叶长、剑叶宽、剑叶基角、剑叶张角、剑叶面积进行相关性分析。采用QTLIciMapping3.0软件进行QTL定位,将图谱数据与表型数据导入软件,以LOD值大于或等于2.5作为选取QTL的标准。定位出的QTL以McCouch等提出的QTL命名方法为参照。采用性状的英文缩写、阿拉伯数字和连字符“-”共同组合命名。
2 结果与分析
2.1 亲本与群体的表型性状
所考查亲本和群体的6个性状统计参数值如表1所示,这6个性状在亲本间差异较大,相应性状的QTL分析有较好的遗传背景[8]。空育131株型较矮,单株产量高于小白粳子,剑叶长、剑叶宽、剑叶面积、剑叶基角、剑叶张角均低于小白粳子。后代存在正负向超亲分离现象,变异范围大,受到多基因控制,表现出数量性状遗传的特点。其中剑叶长、剑叶宽、剑叶面积的群体平均值大于亲本平均值,单株穗重、剑叶基角、剑叶张角的群体平均值小于亲本平均值。单株穗重、剑叶长、剑叶宽、剑叶面积、剑叶基角、剑叶张角的变异系数依次为30.1%、15.03%、8.49%、20.23%、41.65%和36.42%,剑叶基角的变异系数最大,达41.65%,剑叶宽的变异系数最小,为8.49%,说明剑叶基角的变化范围较大,剑叶角度形态差异较明显,而剑叶宽度的变化范围较小。
对F3群体表型数据进行统计分析,单株穗重、剑叶长、剑叶宽、剑叶面积、剑叶基角、剑叶张角均符合正态分布,其表型分布如图1所示。
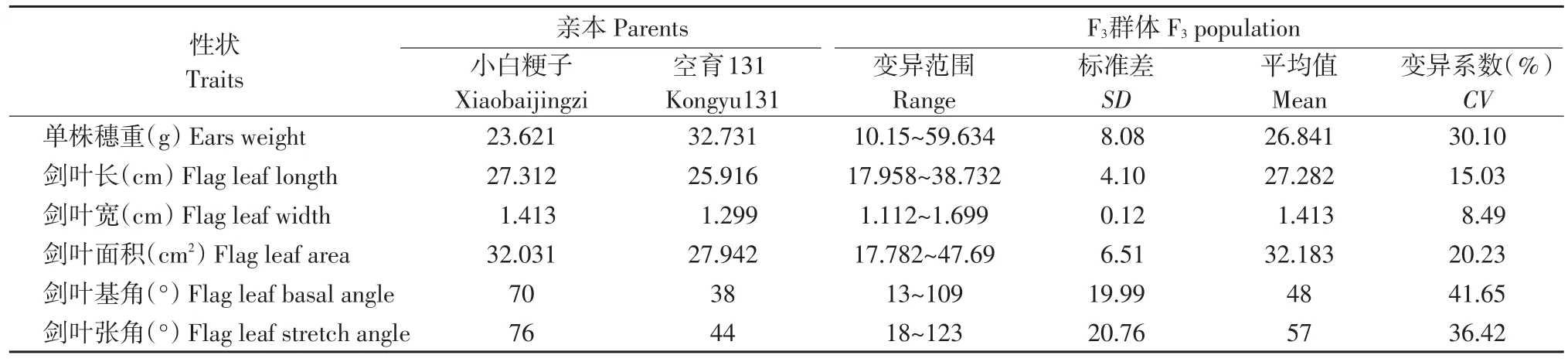
表1 亲本与F3群体的表型分析Table 1 Phenotypes analysis of parents and F3population

图1 亲本与F3群体各性状的表型分布Fig.1 Phenotypic distribution of traits among F3population and parents
2.2 各性状间的相关性分析
对剑叶长、剑叶宽、剑叶面积、剑叶基角、剑叶张角、单株穗重六种农艺性状进行相关性分析。由表2可知,剑叶长与剑叶宽、剑叶面积、单株穗重呈极显著正相关,增加剑叶长度可以增加单株穗重。剑叶宽与剑叶面积呈极显著正相关,与单株穗重呈显著正相关。剑叶面积与单株穗重呈极显著正相关,增加剑叶面积可以提高产量。剑叶的长度、宽度和面积与剑叶的角度两项指标均未达显著水平。剑叶基角与剑叶张角呈极显著正相关,增大剑叶的基角可以使剑叶张角增大。剑叶基角和剑叶张角与单株穗重都达极显著负相关,增大剑叶角度会导致产量下降,这与张克勤等研究结果相一致[9]。王一平等研究发现,剑叶长、剑叶宽和穗重都达极显著正相关[10]。
2.3 QTL定位
利用包括99个SSR标记的水稻分子连锁图谱对剑叶长、宽、面积、基角、张角和单株穗重进行QTL定位研究。在水稻的第1、3、4、5、6、7、9条染色体上共检测到17个QTL。其中控制剑叶长度的QTL 3个,控制剑叶宽度的QTL 4个,控制剑叶面积的QTL 2个,控制剑叶基角的QTL 3个,控制剑叶张角的QTL 2个,控制单株穗重的QTL 3个。
分别在第4、5、7条染色体上检测到与剑叶长相关的QTL,贡献率最大的是qFLL-4,为6.24%,加性效应为-1.51,其增效等位基因来源于父本空育131,表现为后代剑叶长度的缩小,qFLL-5和qFLL-7的效应方向为正,其增效等位基因来源于母本小白粳子,表现为后代剑叶长度的增加,这三个数量性状位点总的贡献率为17.91%,可解释为表型变异的17.91%。

表2 F3群体六个农艺性状之间的相关性分析Table 2 Correlation coefficients of six agronomy traits in F3population
在第3条染色体的RM523-RM231区间,第4条染色体的RM349-RM470区间,第5条染色体的RM509-RM146区间,第9条染色体的RM215-RM410区间各检测到一个控制剑叶宽度的QTL,其中qFLW-3和qFLW-4的加性效应为负,增效等位基因来源于父本空育131,表现为后代剑叶宽度的减小,qFLW-4的贡献率最大,达21.11%。这4个QTL分别解释5.73%、21.11%、3.95%、9.6%的表型变异。
在第4条染色体的RM349-RM470区间和第5条染色体的RM509-RM146区间检测到两个控制剑叶面积的QTL,qFLA-4的贡献率为14.96%,加性效应方向为负,可解释为表型变异的14.96%,增效等位基因来源于空育131。qFLA-5的贡献率为8.41%,加性效应方向为正,可解释为表型变异的8.41%,增效等位基因来源于小白粳子。
共检测到3个控制剑叶基角的QTL,其中第1条染色体上1个,第7条染色体上两个,分别位于RM336-RM180和RM180-RM1377标记区间内,贡献率5.44%~32.53%,这两个数量性状位点的加性效应方向均为负,等位基因来源于空育131,表现为后代剑叶基角角度的减小。
在第1和7条染色体的RM1-RM1247区间和RM1377-RM214区间共检测到2个控制剑叶张角的QTL,贡献率分别为5.68%和7.7%,其中qFLSA-1的效应方向为正,其增效等位基因来源于母本小白粳子,qFLSA-7的效应方向为负,其增效等位基因来源于父本空育131。
共检测出3个控制控制单株穗重的QTL,分别位于第3、6和7条染色体,贡献率6.18%~10.17%,qEW-3表示为负效应,其增效等位基因来源于小白粳子,后代表现为单株产量的减少,位于第6和7条染色体的效应方向为正,表现为后代单株产量的增加。

表3 各性状的QTL定位和效应分析结果Table 3 Effect analysis results of QTL mapping of six traits

图2 群体六个性状的QTL定位图谱Fig.2 QTLs for six traits in the pupulation
3 讨论与结论
剑叶各性状和单株产量均呈连续性分布,超亲现象明显,受到微效多基因控制[11-12],可进行QTL定位研究。Wang等研究发现剑叶角度与产量呈显著负相关[13],结果与本研究一致。本研究在第4条染色体的RM349-RM470区间内和第5条染色体的RM509-RM146区间内都分别检测到同时控制剑叶长度、剑叶宽度和剑叶面积的数量性状位点,在标记区间RM349-RM470内检测到3个QTL的加性效应值都为负,表明剑叶长度或者宽度减少会使剑叶面积减少。在标记区间RM509-RM146内检测到3个QTL的加性效应值都为正,表明剑叶长度或者宽度增加会使剑叶面积增加,这也是一因多效性的具体体现,Zhuang等认为控制不同性状遗传位点的一因多效或者紧密连锁可能是引起性状之间相关性的重要原因[14]。沈波等通过对剑叶长、宽、面积、倒二叶长等表型性状变化规律研究发现,这些性状都是受相同的QTL或基因作用,在多数情况下,在染色体的同一区段内同时检测到控制多个叶片性状的QTL[6]。
本研究共定位到叶片相关性状QTL 14个,单株穗重相关性状QTL 3个。王一平等在第3、4、5、6、7、11条染色体上定位到影响剑叶宽度的数量性状位点[10],本研究在第3、4、5条染色体上同样检测到,但是在第6、7、11染色体上并未检测到,可能是由于分离群体相对不稳定,剑叶角度变异受到多基因控制,易受到环境影响的原因[3]。其中在第3、4条染色体上曾多次报道过控制剑叶宽的位点,但是由于群体不同和分子标记方法的不同,不能确定是否为同一位点[5]。李睿等在第5条染色体的RM598-RM430区间内检测到一个控制剑叶长度的QTL[15],与本研究检测出的qFLL-5相邻,两者效应方向一致,可能是同一位点,具体位置可能在RM509-RM430之间,本研究在第5条染色体的RM509-RM146区间段内检测出一个控制剑叶面积的QTL,和李睿定位的区间段RM430-RM384相邻,两者比较得出在RM509-RM384区间段内存在一个控制剑叶面积的数量性状位点可能性极大。Li等在第2、5、6、7、9条染色体上都检测到影响剑叶角度的QTL[16],本研究大多未检测到,可能是由于环境不同和标记数目相对较少的原因。马大鹏等利用含有177个SSR标记的重组自交系群体对单株产量进行定位研究[17],在第6条染色体上检测到1个QTL,效应方向与本研究一致,很可能是同一位点。
[1]Donald C M,Hamblin J.The biological yield and harvest index of cereals as agronomic and plant breeding criteria[J].Advances in Agronomics.1976,28:361-405.
[2]董国军,藤本宽,滕胜,等.水稻剑叶角度的QTL分析[J].中国水稻科学,2003,17(3):219-222.
[3]沈福成.水稻剑叶长、宽、角度及比叶重的遗传[J].贵州农业科学,1983(6):18-25.
[4]李文滨,赵雪.2009年大豆分子标记及辅助选择育种研究进展[J].东北农业大学学报,2010,41(1):139-148.
[5]蔡晶.水稻剑叶性状的遗传分析和QTL定位[D].北京:中国农业科学院,2009.
[6]沈波,庄杰云,张克勤,等.水稻叶片性状和根系活力的QTL定位[J].遗传学报,2003,30(12):1133-1139.
[7]童汉华,梅捍卫,邢永忠,等.水稻生育后期剑叶形态和生理特性的QTL定位[J].中国水稻科学,2007,21(5):493-499.
[8]刘春燕,齐照明,韩冬伟,等.大豆产量相关性状的多年多点QTL分析[J].东北农业大学学报,2010,41(11):1-9.
[9]张克勤,戴伟民,樊叶杨,等.水稻剑叶角度与主穗产量的遗传剖析[J].中国农学通报,2008,24(9):186-192.
[10]王一平,曾建平,郭龙彪,等.水稻顶部三叶与穗重的关系及其QTL分析[J].中国水稻科学,2004,19(1):13-20.
[11]Yan Y M,Wang X X.Study on genetics of the flag leaf shape in the crossing between indica and japonica[J].Hereditas(Beijing), 1990,12(1):1-4.
[12]Xia Z Y.Studies on inheritance and selection of the leaf shape in Keng rice[J].Acta Agronomica Sinica,1983,9(4):275-280.
[13]Wang X M,Cui K,Guo S Y,et al.Study on the influence of plant type to yield in rice[J].Jilin Agric Sci,1996(1):54-56.
[14]Zhuang J Y,Lin H X,Lu J,et al.Analysis of QTL×enviroment interaction for yield components and plant height in rice[J].Theor Appl Genet,1997,95(5-6):799-808.
[15]李睿,赵姝丽,毛艇,等.水稻剑叶形态性状QTL分析[J].作物杂志,2010(3):26-29.
[16]Li Z K,Paterson A H,Pinson S R M,et al.RFLP facilitated analysis of tiller and leaf angles in rice(Oryza sativaL.)[J]. Euphytica,1999,109(2):79-84.
[17]马大鹏,罗利军,汪朝阳,等.利用重组自交系群体对水稻产量相关性状的QTL分析[J].分子植物育种,2004,2(4):507-512.
QTL analysis of flag leaf characteristics and ears weight in rice
ZOU Detang,WANG Jin,WANG Jingguo,LIU Hualong,LIU Yuqiang,JIA Yan
(School of Agriculture, Northeast Agricultural University,Harbin 150030,China)
F3group derived from the cross of Xiaobaijingzi/Kongyu131,which were significantly different in flag leaf size and ears weight,was used in QTL analysis of the flag leaf size in rice.A total of 99 QTLs referring the flag leaf characteristics,including morphological characteristics(length, width,area,basal angle,stretch angle)and ears weigth were detected.Total 17 QTLs were detected on chromosome 1,3,4,5,6,7 and 9,respectively.Among them,the QTLs for the flag leaf length,width, area,basal angle and stretch angle were 3,4,2,3,2,respectively,the QTLs for the ears weight were 3.The application of the QTLs for flag leaf characters can provide the basis data to improve flag leaf traits,perfect plant types of rice and marker-assisted selection.
rice;flag leaf characteristics;ears weight;quantitative trait loci
S572
A
1005-9369(2014)01-0023-06
2012-02-27
“十二五”农村领域国家科技计划项目(2011BAD35B02-01);国家科技支撑项目(2011BAD16B11)
邹德堂(1965-),男,教授,博士,博士生导师,研究方向为水稻分子遗传育种。E-mail:zoudt@163.com
时间2014-1-9 22:50:19[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140109.2250.017.html
邹德堂,王晋,王敬国,等.水稻剑叶形态与单株产量的基因定位分析[J].东北农业大学学报,2014,45(1):23-28.
Zou Detang,Wang Jin,Wang Jingguo,et al.QTL analysis of flag leaf characteristics and ears weight in rice[J].Journal of Northeast Agricultural University,2014,45(1):23-28.(in Chinese with English abstract)
猜你喜欢
中国水稻科学(2021年6期)2021-11-18 12:06:30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
中学生数理化(高中版.高二数学)(2020年12期)2021-01-13 09:18:18
河北理科教学研究(2020年3期)2021-01-04 01:49:32
新疆农业科学(2020年1期)2020-02-14 03:22:46
浙江农业科学(2019年1期)2019-01-25 05:25:34
新高考·高二数学(2018年6期)2018-11-19 08:57:44
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
扬子江(2016年1期)2016-05-19 22:06:26
新高考·高一数学(2016年3期)2016-05-19 09:08:30
