
具Michaelis-Menten型收获项的双时滞捕食-食饵系统Hopf分支 *
2022-03-12 09:14吕堂红周林华
中北大学学报(自然科学版) 2022年1期
王 菲,吕堂红,周林华
(长春理工大学 数学与统计学院,吉林 长春130022)
0 引 言
在生物学中, 捕食-食饵系统是一类十分重要的描述种群间竞争关系的系统. 多数学者针对捕食-食饵系统的动力学性质包括稳定性、吸引性等进行了丰富的研究, 并且成果斐然[1-4]. 食饵种群除了被捕食者捕食之外, 捕食-食饵种群的双方还会被人类所收获. 文献[5]分析了一类收获率为常数的系统, 对平衡点的稳定性准则进行研究. 文献[6]主要分析了具有收获项的修正 Leslie-Gower 的捕食-食饵模型的鞍结点分支、跨临界分支、Hopf分支等的稳定性, 以及捕食者捕获和生长速率对所提出的模型的影响, 确定了保证物种共存的可行的收获率上限. 文献[7-8]研究了具有非线性的Michaelis-Menten型收获项的模型, 考虑到人为的捕获具有一定的限度, 此类收获项更贴合实际.
文献[9]分析了以下形式的捕食-食饵模型:

(1)
式中:x,y分别代表时间为t时, 种群x和种群y的种群密度;a,b代表存在于种群双方内部的密度制约;c,e的符号代表种群间的关系;q代表人类对两种群的收获;E代表应用到食饵的收获效用, 当E增大到较大的数值时,h1与人类得到的收获呈负相关, 当食饵量减小到较小的数值时,h2与人类的收获能力呈正相关.
文献[9]阐述了系统正平衡点几个性质, 进而探讨了正平衡点附近产生的一个Hopf分支的存在性. 众多学者在对捕食-食饵系统进行研究探讨时发现, 实际上, 不同群体之间的互动是延迟的, 且这种滞后效应会促使系统的动力学行为愈加复杂. 为了修正以往没有考虑到延迟效应的原有模型, 文献[10]分析了具有两个时滞的扩散Lotka-Volterra捕食-食饵系统, 研究了共存平衡点的稳定状态, 得到了在一种特殊情况下, 系统中存在空间非齐次周期解的Hopf分支的结论. 文献[11]通过考虑Holling-III型功能反应函数, 提出了一种具有妊娠期时滞捕食-食饵模型, 得到时滞可以通过Hopf分支引起种群密度的不稳定性和小振幅振荡的结论.
随着研究的深入, 时滞项和收获项逐渐成为捕食-食饵系统的研究重点. 因此, 本文在模型(1)的基础之上, 为了修正以往没有考虑延迟效应的原有模型, 将种群x具有妊娠期、种群y具有妊娠期作为主要考虑因素, 分别引入时滞τ1和τ2, 得出如下具有Michaelis-Menten型收获项的双时滞捕食-食饵系统

(2)
式中:τ1代表食饵种群的妊娠期;τ2代表捕食者种群的妊娠期;f代表食饵种群的出生率;d代表捕食者种群的死亡率; 除此之外的参数与模型(1)意义相同.

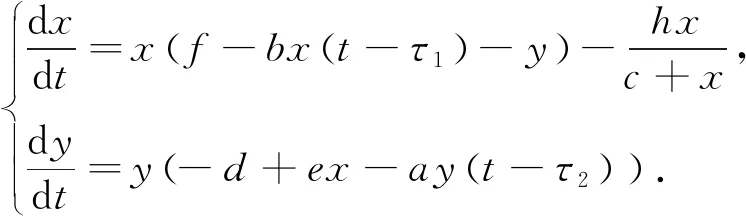
(3)
目前对于模型(3)的研究中, 对分支的周期解的研究甚少. 本文将深入考虑这一问题, 把时滞τ1,τ2作为分支参数, 把零点定理[12]、规范型理论和中心流形定理作为主要研究方法, 研究了模型(3)的动力学性质.
1 局部稳定性及Hopf分支
定理1若是a(f-bc)>(ec+d)且c(af+d)>ha, 那么系统(3)存在仅此一个正平衡点
证明如果系统(3)存在平衡点, 则应满足方程组

(4)
由式(4)中的第二个方程可得
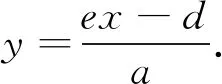
(5)
将式(5)代入式(4)中的第1个方程, 有
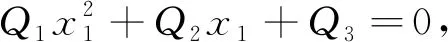
(6)
其中
Q1=-(ab+e),
Q2=a(f-bc)-(ec+d),
Q3=c(af+d)-ha.
当a(f-bc)>(ec+d)且c(af+d)>ha时, 定理1成立.
系统(3)在正平衡点E*处的Jacobi矩阵为

(7)
其中

P12=-x*,P21=ey*,
则有系统(3)的特征方程
λ2+Aλ+B+(Cλ+D)e-λτ1+(Eλ+F)e-λτ2+
Ge-λ(τ1+τ2)=0,
(8)
其中
A=-(P11+P22),B=P11P22-P12P21,
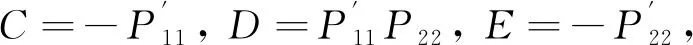
对于两个时滞可能存在的五种不同组合, 进行以下讨论:
情形一:τ1=τ2=0.
系统(3)的特征方程(8)变为
λ2+(A+C+E)λ+B+D+F+G=0.
(9)
因为A+C+E>0, 若:
H1):B+D+F+G>0.
此时, 特征方程(9)的两个根均具有负实部[13], 那么系统(3)的正平衡点是局部渐近稳定的.
情形二:τ1>0,τ2=0.
系统(3)的特征方程(8)变为
λ2+(A+E)λ+(Cλ+D+G)e-λτ1+
B+F=0.
(10)
令λ=iω1(ω1>0)是此方程的根, 代入到式(10) 有
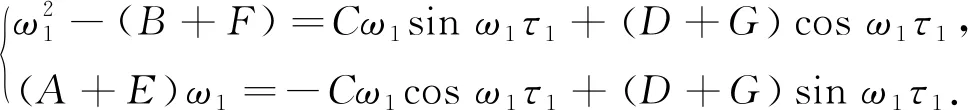
(11)
两边分别平方相加可得
(B+F)2-(D+G)2=0.
(12)

其中
p1=(A+E)2-C2-2(B+F),
q1=(B+F)2-(D+G)2.
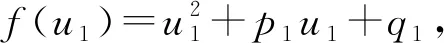
H2):p1>0,q1>0,
那么式(12)的根都是非负根, 进而可得式(10) 的任一根的实部皆为负.
H3):q1<0.
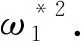
τ1k=

(13)
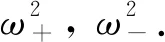

(14)
对式(10)关于τ1求导, 有

(15)
计算后有
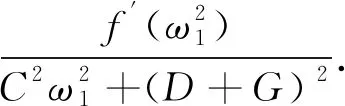
(16)

如果H5)成立, 那么横截性条件便成立:

则有如下结论:
定理2对于系统(3), 假设τ1>0,τ2=0且H1)成立:
1) 如果H2)成立, 则τ1>0时, 系统(3)的平衡点E*是局部渐近稳定的.
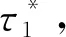
情形三:τ1=0,τ2>0.
此时系统(3)的特征方程(8)变为
λ2+(A+C)λ+(Eλ+F+G)e-λτ2+B+D=0.
(17)
令λ=iω2(ω2>0)是该方程的根, 代入到式(17)有

(18)
两边分别平方后相加可得
D)2-(F+G)2=0.
(19)
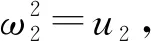
其中
p2=(A+C)2-2(B+D)-E2,
q2=(B+D)2-(F+G)2.
经计算,p2>0,q2>0, 则有
定理3对于系统(3), 当τ1=0,τ2>0时, 在H1)条件下结论同定理2.
情形四:τ1=τ2=τ>0.
定理4针对系统(3), 有τ1=τ2=τ.存在τ0, 使稳定性结论同定理2(2). 其中
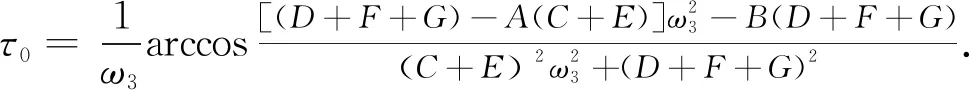
(20)
与情形二同理可证.
情形五:τ1>0,τ2>0.
考虑式(3)中τ1在稳定的区间,τ2作为参数.设λ=iω4为式(8)的根, 代入到式(8)有
消去τ2, 有
h1(ω4)-2h2(ω4)sinω4τ1+2h3(ω4)cosω4τ1=0,
(21)
其中
(B2+D2-F2-G2),
H6): 式(21)至少具有有限正根.

i=1,2,…,j;k=0,1,2,….
(22)
令

ω*=ω4i.
对式(8)关于τ2求导, 有
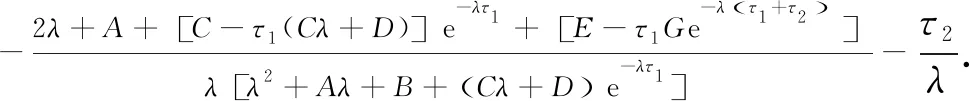
(23)
经计算有

(24)
其中
A1=A+Ccosω*τ1+Ecosω*τ2-
τ1[(Cω*sinω*τ1+Dcosω*τ1)+Gcosω*(τ1+τ2)],
A2=(A+Ccosω*τ1)ω*2-(Dsinω*τ1)ω*,
A3=2ω*-Csinω*τ1-Esinω*τ2-
τ1[(Cω*cosω*τ1-Dcosω*τ1)-Gsinω*(τ1+τ2)],
A4=ω*3-(Csinω*τ1)ω*2+(B+Dcosω*τ1)ω*.
又因为P>0, 假设
H7):Q>0,
则
由上述讨论, 有

2 局部Hopf分支方向及其稳定性
研究在τ1=τ2=τ=τ0条件下, 应用文献[14]提出的两种定理确定系统(3)的Hopf分支的表达式.
先令U(t)=(u(t),v(t))T∈R2, 其中u(t)=x(τt),v(t)=y(τt),τ=τ0+μ,μ∈R, 那么系统(3)在C=C([-1,0],R2)上改写成一类常见的泛函微分方程
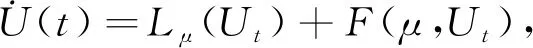
(25)
式中:Lμ∶C→R2,F∶R×C→R2分别由以下形式给出

(26)
F(μ,φ)=(τ0+μ)(F1(μ,φ),F2(μ,φ))T,
(27)
其中
φ=(φ1,φ2)∈C([-1,0],R2),
a13φ1(0)φ2(0),
F2(μ,φ)=b11φ1(0)φ2(0)+b12φ2(-1)φ2(0).
b11=e,b12=-a.
显而易见, 利用Riesz表示定理, 存在一个有界变差的二阶矩阵
η(θ,μ):[-1,0]→R2,
使得

(28)
并且
式中:δ(θ)是Dirac-delta函数.
对于φ∈C1([-1,0],R2), 定义
和
于是, 方程(25)可改写为

(29)
式中:U=(u,v),Ut(θ)=U(t+θ),θ∈[-1,0].
对于ψ∈C1([-1,0],(R2)*), 定义A=A(0)的伴随算子A*为
和一个双线性型

然后, 将A和A*所在的特征方程的特征根iω0τ0与-iω0τ0的特征向量表示为为q(θ)和q*(s).于是
A(0)q(θ)=iω0τ0q(θ),
A*(0)q*(s)=-iω0τ0q*(s).
通过计算, 可以得到


这里

下面给出μ=0处存在的中心流形C0的计算方法, 不妨设Xt为μ=0时方程(25)的解, 规定
z(t)=〈q*(s),Xt〉,
W(t,θ)=Xt(θ)-2Rez(t)q(θ)=
在中心流形C0上, 有
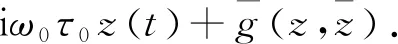
(30)
由式(29)和式(30)得
即
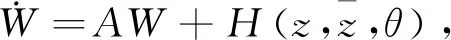
其中:
通过比较系数可得
综合式(27)并比较系数可得
其中,W20(θ),W11(θ)的计算结果如下:
E=(E1,E2)T∈R2,G=(G1,G2)T∈R2是常向量, 它们分别为以下两个代数方程的解
由此可以得到

(31)

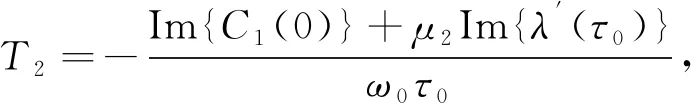
(32)
式中:C1(0)由式(31)给出, 经计算得出μ2,β2,T2的值.故有
定理6令τ=τ0时, 式(32)的表达式确定了分支周期解在中心流形上的性质, 结论如下:
1) Hopf分支的方向由μ2判断.μ2>0(μ2<0), 则Hopf分支是超临界的(次临界的);
2) 分支周期解的稳定性由β2确定.β2<0(β2>0), 则周期解是稳定的(不稳定的);
3) 分支周期解的周期由T2确定.T2>0(T2<0), 则周期解的周期增加(减少).
3 数值模拟
在进行理论分析后, 为了检验其正确性, 选择合适的数对参数进行赋值, 原系统可以表示为
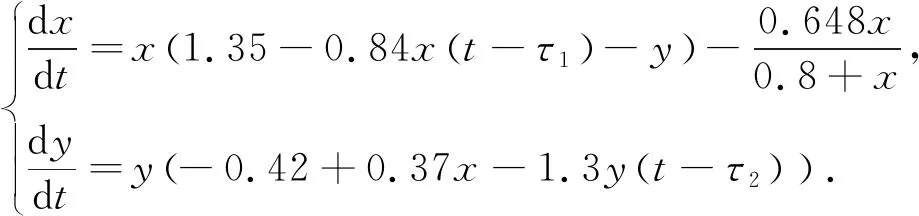
(33)
情形一:当τ1=τ2=0时, 系统(33)的平衡点的波动如图 1 所示.

图 1 当τ1=τ2=0时, 系统(33)平衡点的变化图
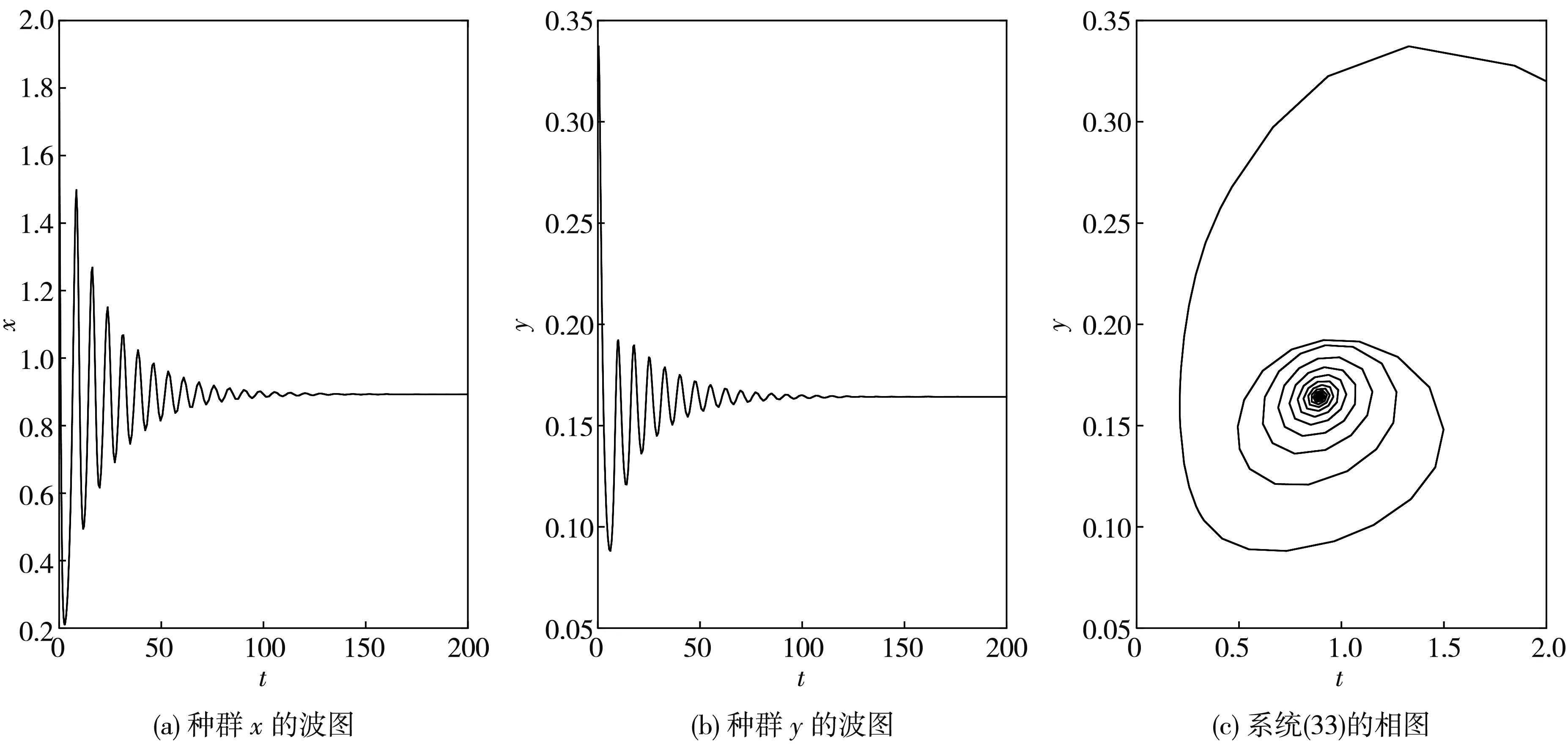
图 2 当时, 系统(33)的波图和相图
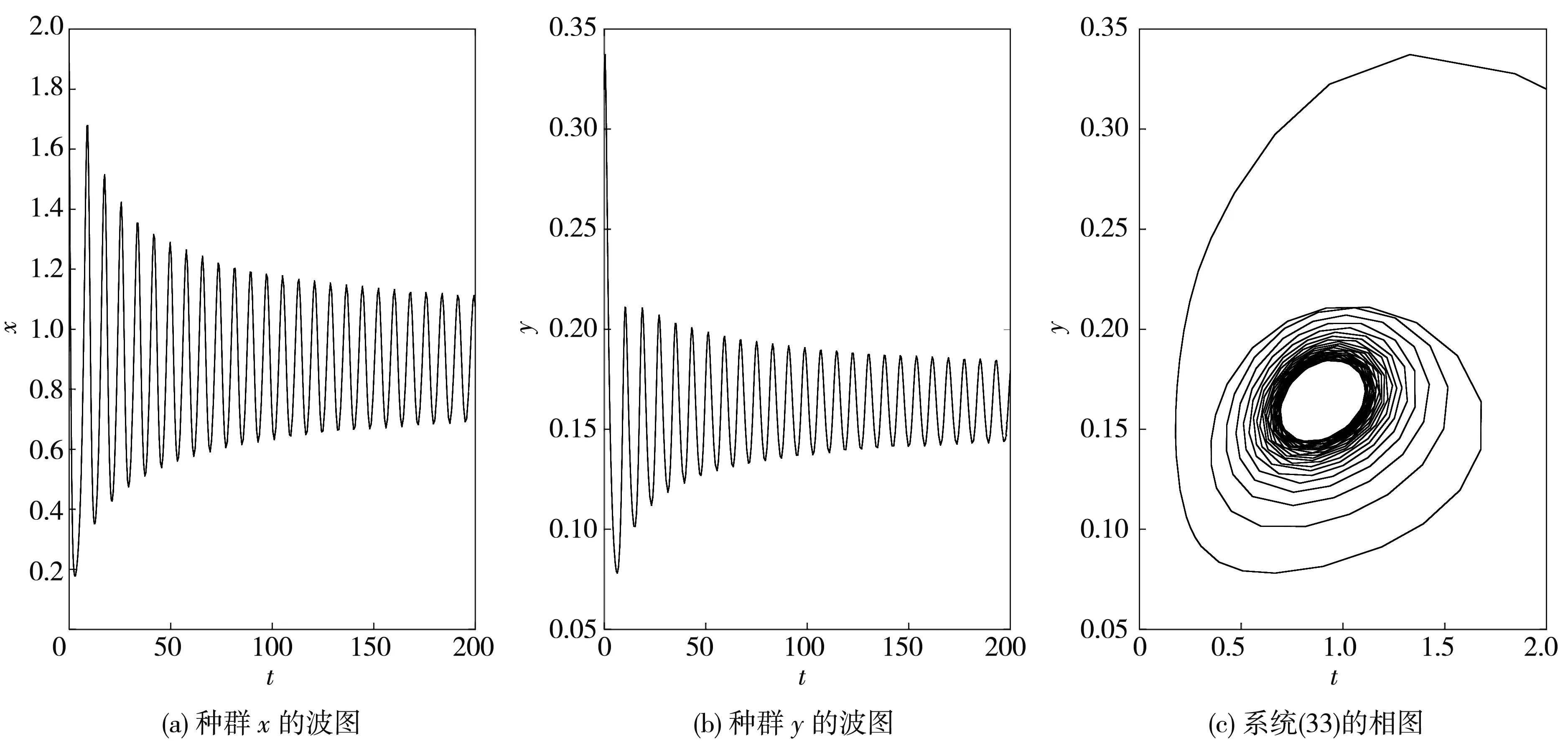
图 3 当时, 系统(33)的波图和相图
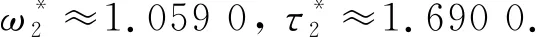
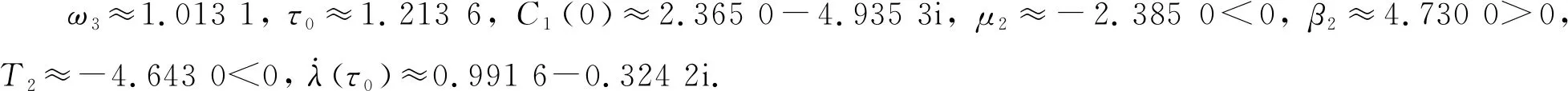
根据定理6可得, 系统(33)在此处发生的Hopf分支是次临界的, 而且分支周期解是不稳定的、分支周期减少. 当τ=1.2<τ0≈1.213 6时, 平衡点E*是渐近稳定的, 如图 6 所示; 当τ=1.35>τ0≈1.213 6时, 平衡点E*稳定状态被打破, 如图 7 所示.
情形五:当τ1>0,τ2>0时, 固定τ1=1.2, 进行计算后得出ω*≈0.659 4,τ*≈1.982 7.那么当τ2=1.8<τ*≈1.982 7时, 平衡点E*是渐近稳定的, 如图 8 所示; 当τ2=2.1>τ*≈1.982 7时, 平衡点E*稳定状态被打破, 如图 9 所示.
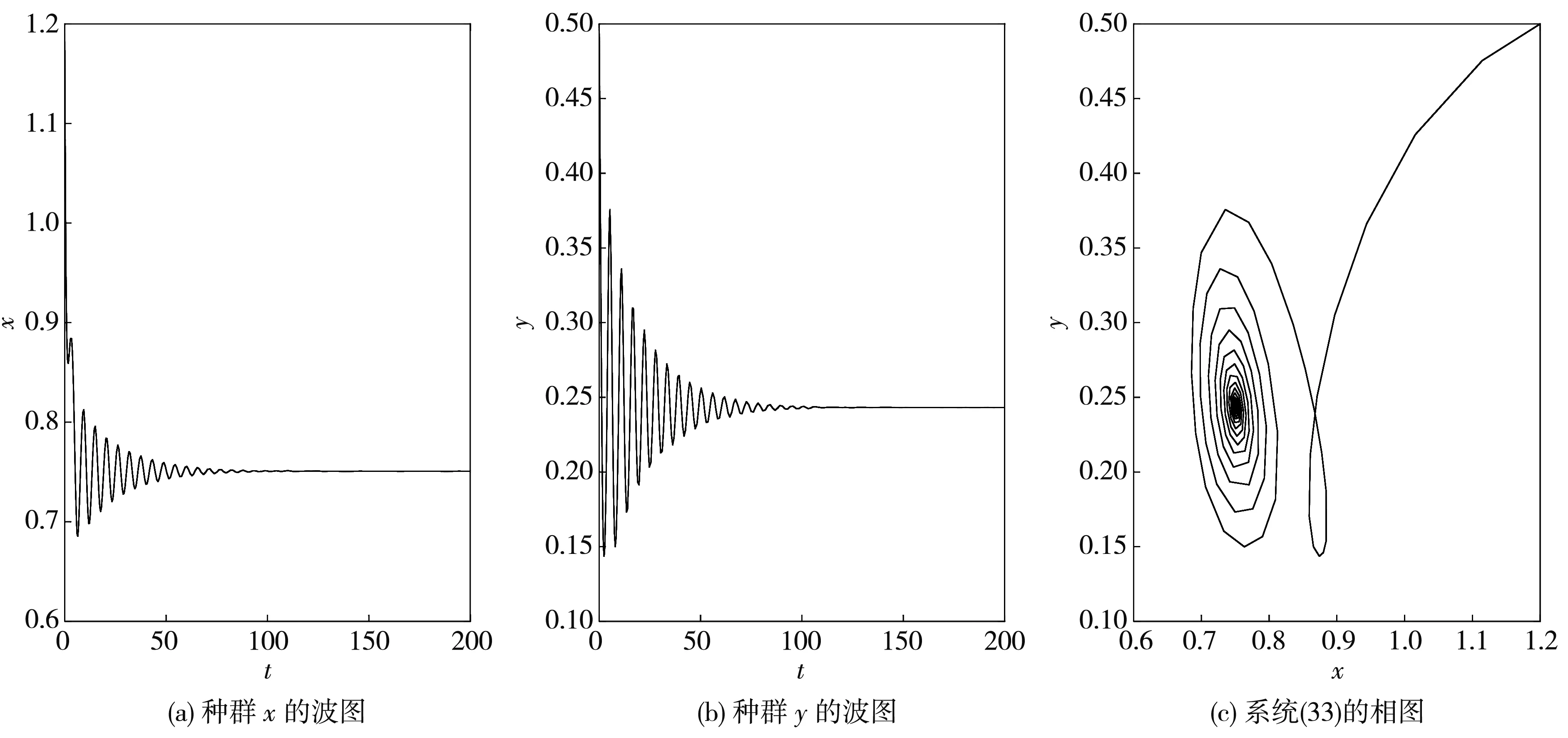
图 4 当时, 系统(33)的波图和相图

图时, 系统(33)的波图和相图

图 6 当τ1=τ2=τ=1.2<τ0≈1.213 6时, 系统(33)的波图和相图
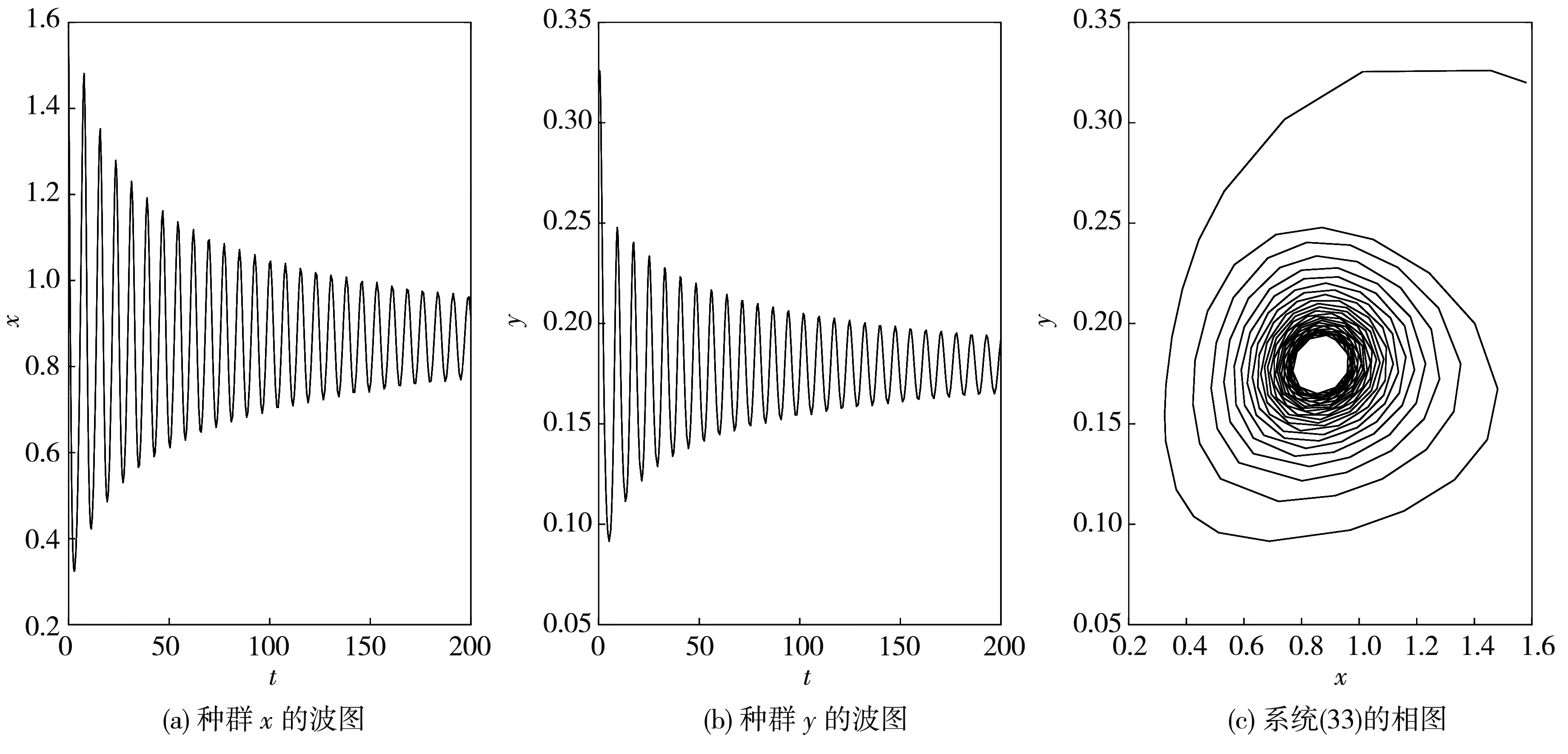
图 7 当τ1=τ2=τ=1.35>τ0≈1.213 6时, 系统(33)的波图和相图

图 8 当τ1=1.2, τ2=1.8<τ*≈1.982 7时, 系统(33)的波图和相图
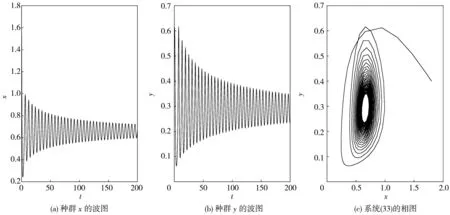
图 9 当τ1=1.2, τ2=2.1>τ*≈1.982 7时, 系统(33)的波图和相图
4 结 论
本文以时滞τ1,τ2为参数对一类具有Michaelis-Menten型收获项的双时滞捕食-食饵系统进行了探讨. 经过理论分析及验证发现, 只考虑种群x的妊娠期时滞τ1时, 时滞变化对系统的持续生存没有影响; 只考虑种群y的妊娠期时滞τ2, 或同时考虑种群x和种群y的妊娠期时滞时, 种群的时滞变化会影响其种群数量的变化. 当时滞未达到临界值时, 系统呈稳定状态; 当时滞达到临界值时, 系统的稳定状态被破坏, 此时发生Hopf分支并产生周期解.
猜你喜欢
中北大学学报(自然科学版)(2022年4期)2022-09-23
吉首大学学报(自然科学版)(2022年1期)2022-08-11
吉首大学学报(自然科学版)(2022年1期)2022-08-10
黑龙江大学自然科学学报(2022年1期)2022-03-29
计算机系统应用(2021年10期)2022-01-06
廊坊师范学院学报(自然科学版)(2021年2期)2021-09-10
教育教学论坛(2019年27期)2019-07-30
智富时代(2018年4期)2018-07-10
智富时代(2018年4期)2018-07-10
上海师范大学学报·自然科学版(2018年3期)2018-05-14
