
SSR荧光检测解析224份山西玉米自交系的遗传结构与遗传关系
2022-03-02 06:29尚春树常利芳白建荣
中国农学通报 2022年5期
李 锐,尚 霄,尚春树,常利芳,闫 蕾,白建荣
(1山西农业大学农学院,太原 030031;2山西强盛种业有限公司,太原 030031)
0 引言
杂种优势的利用是大幅度提高玉米产量和改良品质的有效途径。目前,玉米已成为中国第一大粮食作物,是中国北方包括山西的主要粮食作物。山西由于多样的生态环境,形成了5个玉米生态种植区域[1],几乎包括了全国乃至全世界的玉米生态类型,每年审定的品种在100个以上。面对目前玉米品种种质资源的与日俱增,对玉米遗传种质的亲缘关系及遗传差异的分析和掌握程度,是改良自交系和利用杂种优势的基础,在玉米种质创新和利用工作中具有重要意义。
SSR标记已广泛应用于玉米种质资源的遗传多样性研究[2-13]。虽然传统的电泳技术已进行了多次优化,但仍难以满足试验中高效、准确、高通量的需求,因此,毛细管荧光标记技术逐渐受到关注。该技术是以DNA测序技术为基础发展起来的检测方法,具有高效、自动化等优点。易红梅、郝晨阳的研究表明,用毛细管荧光引物标记技术检测效率显著高于聚丙烯凝胶电泳,结果更为精确、灵敏,更适用于材料的高通量检测分析[14-15]。到目前为止,已有许多利用荧光标记进行玉米[16-17]和其他作物[18-21]种质资源遗传多样性的研究报道,但DNA自动测序仪成本较高,采用荧光引物标记也需要较高成本。为克服其缺点,近些年来,新发展了内插荧光染料标记法的毛细管电泳荧光检测技术,该种染料背景荧光非常低,与DNA的结合力强,然而当其结合到核酸上时,产生很强的荧光以及高的量子产率。由于不需要标记引物,使用该染料标记法的设备成本约DNA自动测序仪的五分之一,实验成本也大幅下降,实验结果同样准确。另外,该方法还有DNA自动测序仪没有的模拟电泳胶图,形象、直观,非常受到研究人员的欢迎[22-24]。
山西强盛种业有限公司是山西最大的种业公司,也是全国种业骨干企业。二十多年来,公司一直坚持育、繁、售一体化策略,每年审定品种5个以上,有山西省内、海南及新疆制种基地,每年推广、销售大量的种子,是亿元级企业。该公司玉米育种团队多年来收集了大量的玉米种质,在此基础上改良、培育了许多优良自交系,这些自交系在很大程度上代表了山西的玉米种质,但还没有对它们的遗传多样性及遗传结构进行分析。本研究用毛细管电泳荧光检测技术,对该公司多年收集和选育的优良自交系进行遗传多样性及遗传结构的分析,为山西省的商业育种发展提供有力支撑。
1 材料和方法
1.1 材料
1.1.1 试验材料 300份玉米自交系由山西强盛种业有限公司提供。经SSR标记分析后,选择了每份材料在每个位点都是扩增单位点的材料224份,其中包括目前生产上主要推广品种‘先玉335’、‘先玉508’、‘先玉696’和‘郑单958’的亲本自交系‘PH6WC’(‘先玉335’,‘先玉508’,‘先玉696’的母本)、‘PH4CV’(‘先玉335’父本)、‘PHB1M’(‘先玉696’的父本)、‘PH5AD’(‘先玉508’的父本)、‘郑58’(‘郑单958’母本)和‘昌7-2’(‘郑单958’父本)。试验在山西农业大学农学院分子设计育种实验室,于2018年6月—2020年3月完成。
1.1.2 SSR引物 采用中华人民共和国农业行业标准(NY/T 1432—2014《玉米品种鉴定DNA指纹方法》)的20对核心引物和20对辅助核心引物[25]。
1.2 实验方法
1.2.1 基因组DNA提取、纯化和检测 采用CTAB法[26]混合提取群体DNA,每个群体包括2个样本,每个样本由30个单株(随机取样)的黄化芽苗混合而成。采用CTAB法提取并纯化DNA,经纯度和浓度检测后,-20℃保存备用。
1.2.2 PCR扩增及产物检测 扩增反应在C1000扩增仪上进行。PCR反应总体系为20 μL,10xPCR Buffer(25 mmol/L Mg2+)2 μL,2.5 mmol/L dNTPs 1 μL,10 μmol/L SSR引物各 1 μL,Taq DNA 聚合酶 0.5 U,DNA模板80 ng,ddH2O补足体积。PCR反应程序为95℃预变性5 min,1个循环;94℃变性1 min,60℃复性30s,72℃延伸1min,共35个循环;最后于72℃延伸10min。
扩增产物采用美国AATI公司Fragment Analyzer 96毛细管核苷酸片段分析系统进行电泳检测,获取所有扩增产物的峰型电泳图。Tag DNA聚合酶购自宝生物工程(大连)有限公司;引物和dNTP等试剂购自上海生工生物技术有限公司。
1.2.3 数据统计和分析 利用PROSize 3.0软件将荧光检测获取的峰型电泳图转换成扩增产物的片段长度。利用PowerMarker V3.25软件统计每对引物的等位变异数和多态性信息量(PIC);标记索引系数(MI)按公式(1)计算。

Allele为该引物的等位变异数。利用Structure V2.3.4软件进行类群划分,研究材料的类群个数通过ΔK确定,画出基于模型的群体遗传结构图,计算各自交系的Q值。利用NTSYS-PC 2.1软件按照UPGMA方法对各材料进行聚类分析。统计每份材料的特有等位变异数(只有1个供试材料有的等位变异)。
2 结果与分析
2.1 遗传多样性分析
224个玉米自交系在40个SSR位点的遗传多样性汇总表1。共检测到171个等位变异,每个位点检测出2~8个等位变异,平均4.28个;其中,bnlg439wl位点和bnlg249k2最高,各有8个等位变异;4个位点等位变异最低,只有2个。40个位点的多态性信息量PIC值变化 范 围 在 0.05~0.77,平 均 每 个 位 点 0.44;其 中bnlg249k2位点的PIC值最高,达0.77。标记索引指数MI变化范围在0.16~6.13,平均每个位点为2.02。基因多样性变化范围在0.05~0.79之间,平均每个位点0.50。6个位点有优势等位变异(等位变异频率>80%)出现,其中在umc1335y5位点的3个等位变异中,2号等位变异的频率高达90.5%;同样在phi072k4位点的3个等位变异中,1号等位变异的出现频率高达94.6%,而phi053k2位点中2个等位变异的频率比较均匀(未列出)。这些高频率等位变异的出现,说明种质遗传多样性均匀度较差,从而降低了其遗传多样性指数。因此,将等位变异多少及分布2个指标相结合,可以全面衡量不同种质在某一位点的遗传多样性。
2.2 特异等位变异分析
在224份自交系中,有7份种质共存在特异等位变异9个,分布在8个SSR位点。其中,223号材料有3个特异等位变异,其余6份种质各有1个特异等位变异(表2)。按位点分析,umc2084w2位点出现的特异等位变异为2个,其余7个位点上各有1个(表1)。种质的特异等位变异是其身份鉴定的重要特征,但也是相对的。随着分析种质数量的增加,特异等位变异的数目会趋于减少;但随着检测位点的增加,特异等位变异的数目有可能增加。

表1 224份自交系在40个核心SSR标记的遗传多样性

表2 有特异等位变异的自交系
续表1
2.3 遗传结构分析及类群的遗传多样性分析
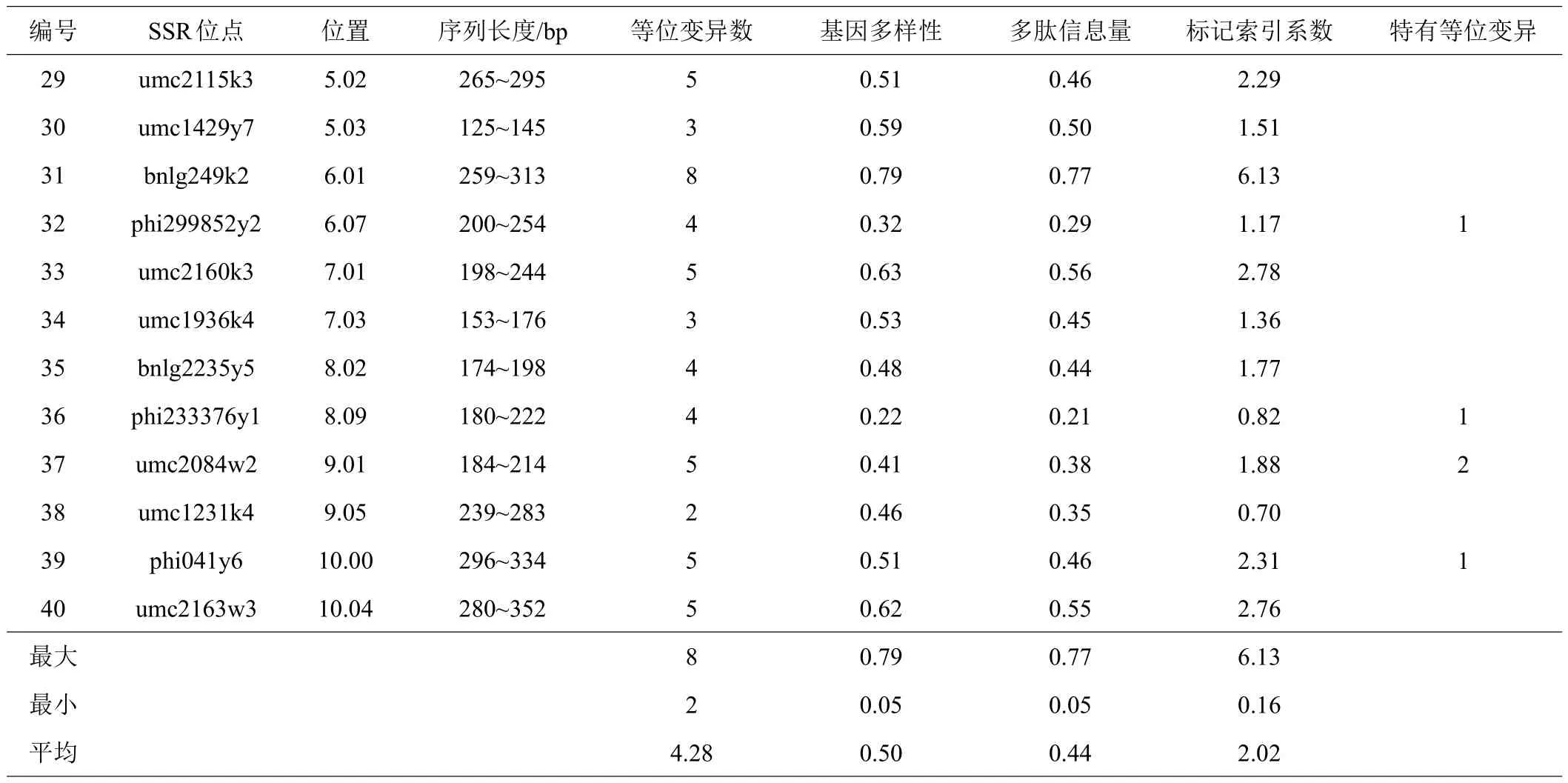
应用Structure2.3.4软件,对224个玉米自交系进行群体遗传结构分析,当类群数(K值)在2~10之间变化时,ΔK值在K=3时出现明显峰值,表明224份自交系可以划分为3个类群(图1)。其中104份划分为Reid群,以‘PH6WC’和‘郑58’为代表,占参试样本数的46.4%;88份划分为 Lancaster群,以‘PH4CV’、‘PHB1M’、‘ PH5AD’为代表,占参试样本数的39.3%;32份划分为唐四平头群,以‘昌7-2’为代表,占参试样本数的14.3%(图2)。根据最大Q值分布,Q值>0.8和0.9的自交系分别占64.3%和61.2%。然而,224个玉米自交系中有18个Q值<0.6,占自交系总数的8.0%,表明每个类群有部分自交系含有其他类群遗传成分,即存在跨类群自交系(表3)。在3个类群中,塘四平头群和兰卡斯特群群的平均等位变异数为4,瑞德群的最低为3;多肽信息量、基因多样性与标记索引系数的趋势类似,塘四平头群最高,兰卡斯特群次之,瑞德群最低(表4)。这些结果表明,塘四平头群的遗传多样性最高,兰卡斯特群次之,Reid群最低。

表3 模型聚类的各群自交系最大Q值分布

表4 模型聚类的各类群遗传多样性

图1 ΔK的变化曲线
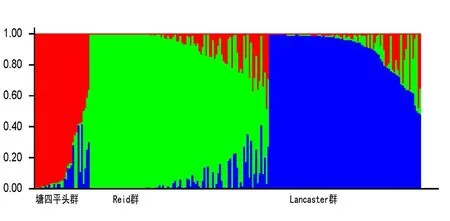
图2 224份自交系的遗传结构
2.4 遗传距离聚类分析
利用40个SSR标记的171个等位变异,对224个自交系进行遗传距离聚类。在相似系数大约为0.71处可分为5大聚类群,结合各聚类支中是否含有不同种质类群的代表性自交系,确定类群的名称(图3)。从图中看出,瑞德群、兰卡斯特群是两个最大的群,二者数量之和超过了材料的90%;有10份材料属于唐四平头群;另有10个自交系的类群归属不明确,定为其他群。兰卡斯特群可明显看出有335父本群、696父本群与508父本群和其他兰卡斯特群,即有亚类分化趋势。
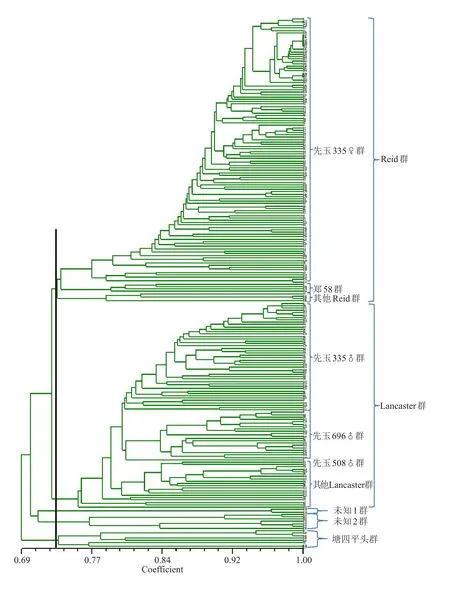
图3 224份玉米自交系SSR标记遗传距离聚类图
3 讨论
当前,商业育种已成为玉米育种的主要趋势[27-28]。玉米商业育种的关键一步就是分析公司现有种质资源并重新建立杂种优势群。因此,本研究对近年来育种企业在商业化育种中获得的玉米自交系进行了遗传分析。
3.1 应用合适的标准测验种
中国曾经有6个标准测验种(‘齐319’、‘黄早四’、‘B73’、‘Mo17’、‘掖478’和‘丹340’)作为群体划分依据,它们分别代表中国玉米生产上主要应用的PB、四平头、PA、兰卡斯特、瑞德和旅大红骨等杂种优势群[29]。经过多年采用多种技术路线分析,已经把中国玉米种质合并为3个杂种优势群(列),即A群、B群和D群[30]。商业育种是设计育种,玉米商业化育种中种质改良与种质创新的基本策略是循环育种,它们的共同基础是统一和简化的杂种优势模式[31]。如果杂种优势类群数目过多,育种效率有降低的可能。在明确杂种优势模式下用向两个方向推开的育种策略进行选系是玉米商业化育种所需[32]。这些年来,按照这个原则,公司主要用本地种质对优良杂交种先锋系列和‘郑单958’的亲本进行了改良,选育了大量的自交系。越是引进和拥有丰富的种质资源,越是需要简化杂种优势模式,这样才便于管理和实践,这就非常需要确定少而精且实用的标准测验种。因此,本研究没有用大多数研究常规用的标准测验种,而选择了大面积推广品种‘先玉335’、‘先玉508’、‘先玉696’和‘郑单958’的亲本作标准测验种。研究结果准确反映了改良过程中的育种策略,也能对获得材料可进行针对性的分析,说明笔者选择的标准测验种是非常正确和有效的。
3.2 所选用SSR标记的通用型及结果可比性
许多研究在玉米杂种优势类群划分时使用的SSR标记类型及数量因不同的研究者而异,因而,研究结果对其他研究者的参考价值不太大。本研究所用的40个SSR标记,是北京农林科学院玉米研究中心通过10多年的努力,从3000多个SSRs中经过反复筛选后确定的一套核心标记,具有稳定可靠、揭示的多样性水平高、且部分标记与农艺性状连锁的特点,现已成为中华人民共和国农业行业标准《玉米品种鉴定DNA指纹方法》的20对核心引物和20对辅助核心引物,非常具有通用型[33]。该套引物非常适合研究中国玉米种质资源,已广泛应用于中国玉米资源指纹构建、一致性鉴定及遗传结构分析[34]。本研究采用的毛细管电泳系统和农业部《玉米品种鉴定DNA指纹方法》所用的电泳系统及统计方法完全相同,结果具有可比性。这也是玉米商业化育种技术的标准化和流程化所需要的标准检测条件,体现了商业育种体系的价值和效率[28]。
3.3 遗传多样性分析
研究结果表明,每个位点检测出2~8个等位变异,平均4.28个;平均每个位点多态性信息量PIC、标记索引指数、基因多样性指数分别为0.44、2.02和0.50;有些位点存在优势等位变异。这一切说明,这些自交系的遗传多样性不太丰富,这主要是由于大部分的自交系定向由骨干自交系而来,加上环境压力和人为选择,导致同一单位选育的自交系遗传相似度较高,血缘关系较近。因此,在自交系的改良和创育中,需要加大其他血缘及地方种质的引用;另外,在分析和选择中,加强每个自交系特有等位变异的分析,对含有特有等位变异的自交系要重点注意及有目标、有计划地选留,加大群体的遗传多样性。另外,分析群体内的等位基因多样性与频率来判断群体间的遗传距离及对选择的响应,达到既实现在群体内积累有益基因,又没有缩小群体间的距离,从而实现有益基因分别向两个不同群体积累,为利用杂种优势奠定基础[31]。
3.4 综合考虑两种聚类方法结果,获得全面的分析
研究结果显示,模型聚类中18份(8.0%)自交系Q值<0.6,这可能由以下原因造成。一是由于这些材料在选择中没有向两边推,导致杂优类群属性不典型,属于中间性材料;另一方面,也有可能是在改良中有其他的育种方案,这些自交系有较多其他血缘成分造成。在遗传聚类中,90%的材料聚在了瑞德群和兰卡斯特群中。两种方法划分群体的依据不同但结果大致一致,模型聚类可通过各自交系的Q值分布解析各自交系的遗传基础,细化自交系间的分群;遗传聚类则反映了自交系间的遗传距离及亚群间的遗传关系。因此,综合两种方法的结果去考虑,可准确地进行杂种优势类群划分及遗传关系分析,这更有利于指导田间的育种工作、提高育种效率。
4 结论
本研究根据40个位点的SSR标记,分别采用模型聚类法和遗传距离聚类法,对山西省商业育种公司的224份种质进行了遗传结构分析、优势类群的划分及亲缘关系的确定。其结果既能结合田间表现对不适合的材料进行淘汰,提高选择效率,又能在优选的基础上指导进一步种质改良与创新。同时,在大群分类的基础上,选择不同的亚群有计划地进行亲本组合,选配杂交种。因此,本研究为这些自交系的改良和利用及选配新杂交组合奠定了理论依据,也为其它玉米育种者提供了研究思路与方法作参考。
猜你喜欢
广西植物(2022年8期)2022-09-07
江苏农业科学(2022年6期)2022-04-15
绿色科技(2019年14期)2019-11-19
中学生物学(2019年7期)2019-10-17
江苏农业科学(2019年5期)2019-09-02
江苏农业科学(2017年17期)2017-11-15
中学生物学(2017年7期)2017-08-23
科技资讯(2016年32期)2017-03-31
农民致富之友(2016年5期)2016-10-21
广西农学报(2015年3期)2015-10-14
