
舟山群岛东侧海域蟹类群落结构及其与环境因子的关系
2022-02-18 12:58牛威震韩磊俞存根郑基许永久颜文超
海洋学报 2022年2期
牛威震,韩磊,俞存根*,郑基,许永久,颜文超
(1.浙江海洋大学 水产学院,浙江 舟山 316022)
1 引言
舟山群岛地处长江口与杭州湾入海口,其渔场环境受台湾暖流、浙江沿岸流和黄海冷水团的交互影响,水质肥沃,具有较高的初级生产力水平和丰富的饵料生物,是众多鱼类、虾蟹类等生物优良的产卵场和索饵场。蟹类是海洋无脊椎动物的重要类群之一,从潮间带到大陆架浅海以及大洋深海底均有分布,它不仅是海洋生物群落中的重要生物类群,很多种类还是人类自古以来的重要捕捞对象。东海的蟹类资源十分丰富,根据俞存根等[1]研究可知,东海分布的蟹类有324 种,可食用蟹类约有20 种。对东海及舟山渔场的蟹类资源研究报道已有不少,如沈嘉瑞和刘瑞玉[2]、戴爱云等[3]对东海蟹类的分类做过研究,俞存根等[4-12]对东海蟹类种类组成、区系特征、数量分布、群落结构特征进行了报道,并研究了细点圆趾蟹(Ovalipes punctatus)、日本蟳(Charybdis japonica)、锈斑蟳(Charybdisferiatus)、武士蟳(Charybdis miles)等主要经济蟹类的生物学特性,评估了东海蟹类的资源量等。薛利建和卢占晖[13]借助生物多样性分析了东海中部蟹类群落结构特征;陈小庆等[14]通过划分生态群落研究了东海中南部蟹类群落结构与环境因子之间的关系;张洪亮等[15]分析了浙江南部近岸蟹类春季群落结构特征,并与东海海域的生物多样性和优势种等进行了比较;卢衎尔等[16]研究报道了浙江近海蟹类群落结构与环境因子的关系。舟山渔场的蟹类资源,主要集中在舟山沿岸海域,如刘惠等[17-18]在研究舟山沿岸海域甲壳类群落结构特征、生态位及其与种间联结性时有所涉及。本文根据2018 年4 月(春季)和10 月(秋季)在舟山群岛东侧海域开展渔业资源底拖网调查所获得的蟹类数据资料,开展调查海域蟹类资源种类组成、优势种、生物多样性等群落结构特征,同时分析了蟹类数量分布及其群落结构与温、盐等环境因子之间的关系。因关于舟山群岛东侧海域蟹类群落分析的文章较少,希望对本研究区域蟹类群落结构,特别是舟山蟹类群落结构与舟山及长江口毗连海域生态系统关系的分析可作为基础研究资料进行分享。本研究可进一步丰富该海域蟹类群落生态学资料,为蟹类资源的合理利用和科学管理提供科学依据。
2 材料与方法
2.1 数据来源
本文所用数据采于2018 年4 月(春季)和10 月(秋季)在舟山群岛东侧海域开展的渔业资源底拖网调查所获得的蟹类资料,调查共设置16 个站位,调查范围为29°10′25.54′′~30°45′0.00′′N,122°53′3.88′′~124°48′28.31′′E,调查船为“浙渔科2”科考船,主机功率为1 007 kW,渔船有配备卫星导航设备、探测仪(测深仪)、甚高频VHF 以及其他助渔助航设备,拖网网具规格为1 020 目×80 mm。调查时每站位作业时拖拽约1 h,拖速约为3.5 kn,将每个调查站位所捕获的渔获物全部装入样品袋,并记录站位号和样品袋号,冰鲜保存后带回实验室进行生物学测定。测重仪器为电子天平,精确度为0.1 g。调查方法按《海洋渔业资源调查规范》(SC/T 9403-2012)执行。温度、盐度及水深调查测量与渔业资源底拖网调查同步进行,温度、盐度及水深等环境因子数据采用温盐深仪(Conductance Temperature Depth,CTD)同步获取,调查站位设置如图1 所示。
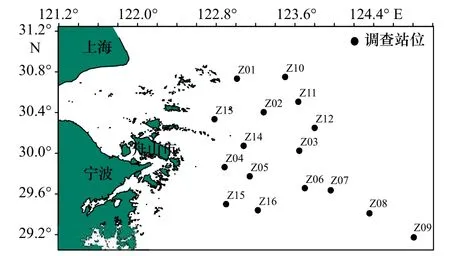
图1 舟山群岛东侧海域渔业资源调查站位分布Fig.1 Distribution of fishery resources survey stations in the eastern sea area of the Zhoushan Islands
2.2 数据处理
2.2.1 优势种计算公式
蟹类优势种采用相对重要性指数(Index of Relative Importance,IRI)来确定,其计算公式为
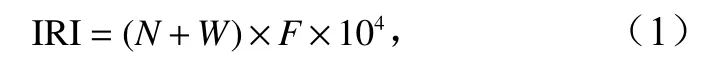
式中,N和W分别代表为某种蟹类的尾数及重量占蟹类总尾数及总重量的百分比;F为某种蟹类出现的站位数占调查总站位数的百分比。IRI 大于1 000 的视为优势种,100~1 000 的视为常见种[19]。
2.2.2 生物多样性分析
采用Shannon-Wiener 多样性指数(H′)、Pielou 均匀度指数(J′)、Margalef 丰富度指数(D)来计算分析调查海域的蟹类生物多样性,公式分别为

式中,Pi是第i种蟹类生物量占总生物量的比例;S为蟹类总种数;N为蟹类总尾数。
2.2.3 群落结构相似性分析
本文运用Bray-Curtis 相似性系数,并采用Cluster等级聚类和非度量多维标度分析(Non-Metric Multi-Dimensional Scaling,NMDS)方法分析调查海域蟹类组成特征。计算公式为

式中,B表示Bray-Curtis 相似性系数;S表示种类数;xij表示第i物种在j站位中的渔获重量;xim表示第i物种在m站位中的渔获重量。以胁迫系数衡量NMDS 二维点阵分布效果,通常认为胁迫系数小于0.05时,具有很好的代表性;胁迫系数为 [0.05,0.1) 时,结果基本可信;胁迫系数为 [0.1,0.2) 时,具有一定解释意义。
2.2.4 种类更替率
以种类更替率分析蟹类种类的季节更替,公式为
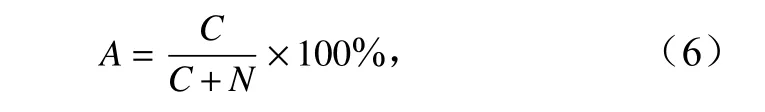
式中,A为更替率,表示与上一个季节比较的更替情况;C为两季节间种类增加数或减少数;N为两季节间相同种类数。
2.2.5 典范对应分析
采用典范对应分析(Canonical Correspondence Analysis,CCA)方法分析蟹类群落与环境因子之间的关系,具体计算利用软件CANOCO 5.0,排序结果用站位-环境因子关系双序图表示。
本文蟹类生态习性特征参考文献[20]的相关研究;蟹类种名录和种类更替率参照文献[21-22]的研究报道;调查站位图和蟹类数量分布图采用Surfer 11.0 软件绘制,生物多样性分析采用Excel 2003 和Primer 5.0 软件完成,春、秋季渔获量之间显著性检验使用SPSS 21.0 完成。
3 结果
3.1 种类组成及优势种
根据2018 年4 月(春季)与10 月(秋季)两个航次底拖网调查取样样品,共鉴定出蟹类40 种,隶属于12 科23 属,其中,春季29 种,隶属于9 科18 属,秋季35 种,隶属于11 科21 属。物种组成上,两季均以梭子蟹科种类数最多(春、秋季均为12 种),其次是玉蟹科(春、秋季均为5 种),春、秋季蟹类种类数更替率为20%。蟹类生物种类组成如表1 所示。
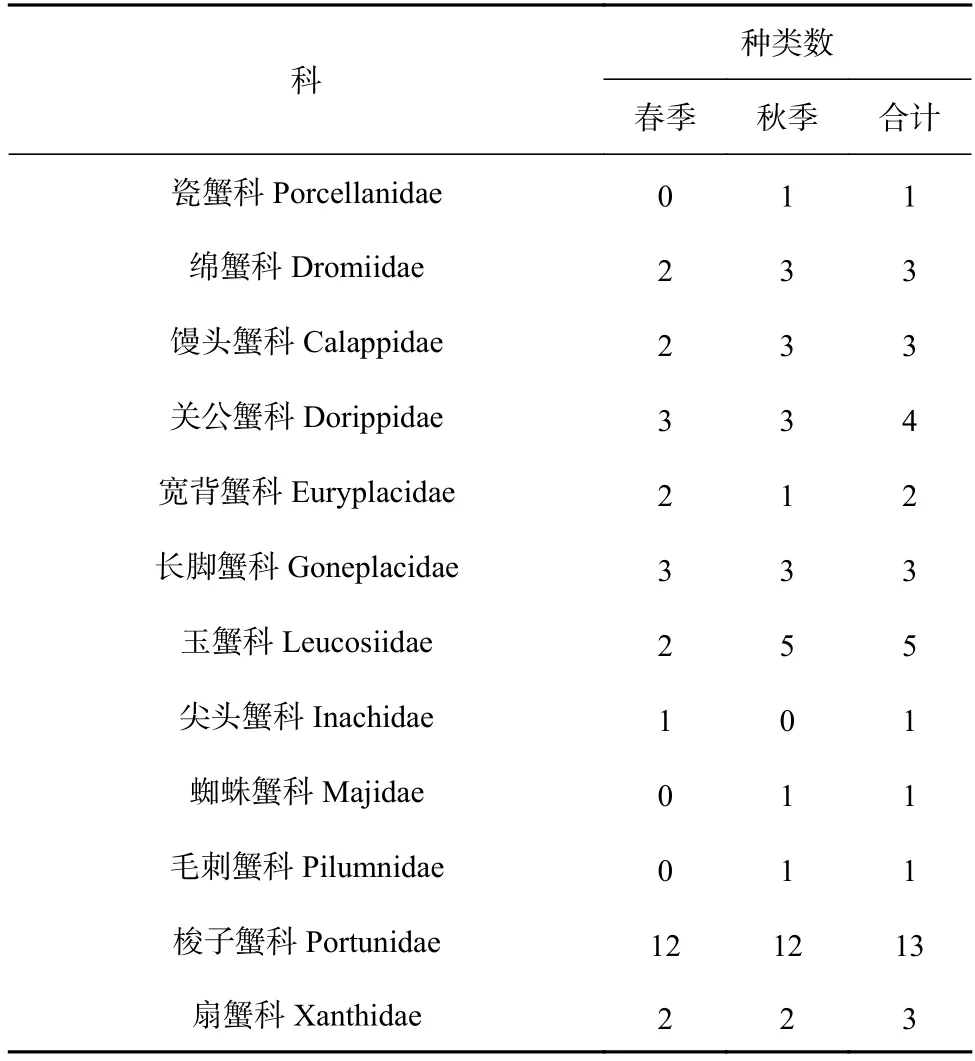
表1 调查海域春秋季蟹类种类数的组成Table 1 The composition of crab species in spring and autumn in the investigated sea area
蟹类优势种组成如表2 所示,春季优势种为双斑蟳(Charybdis bimaculata),常见种有日本英雄蟹(Achaeusjaponicus)、绵蟹(Dromia dehaani)、日本蟳、银光梭子蟹(Portunus argentatus)、长手隆背蟹(Carcinoplax longimana)、三疣梭子蟹(Portunus trituberculatus)6 种;秋季优势种为双斑蟳和三疣梭子蟹,常见种为日本蟳。从生态习性上来看,调查海域春、秋季蟹类区系特征主要为广温、广盐性的暖水种。

表2 调查海域主要蟹类相对重要性指数及生态习性Table 2 The relative importance index and ecological habits of the main crabs in the investigated sea area
3.2 生物多样性
调查海域附近各站位春、秋季蟹类生物多样性及平面分布如表3、图2 至图4 所示。
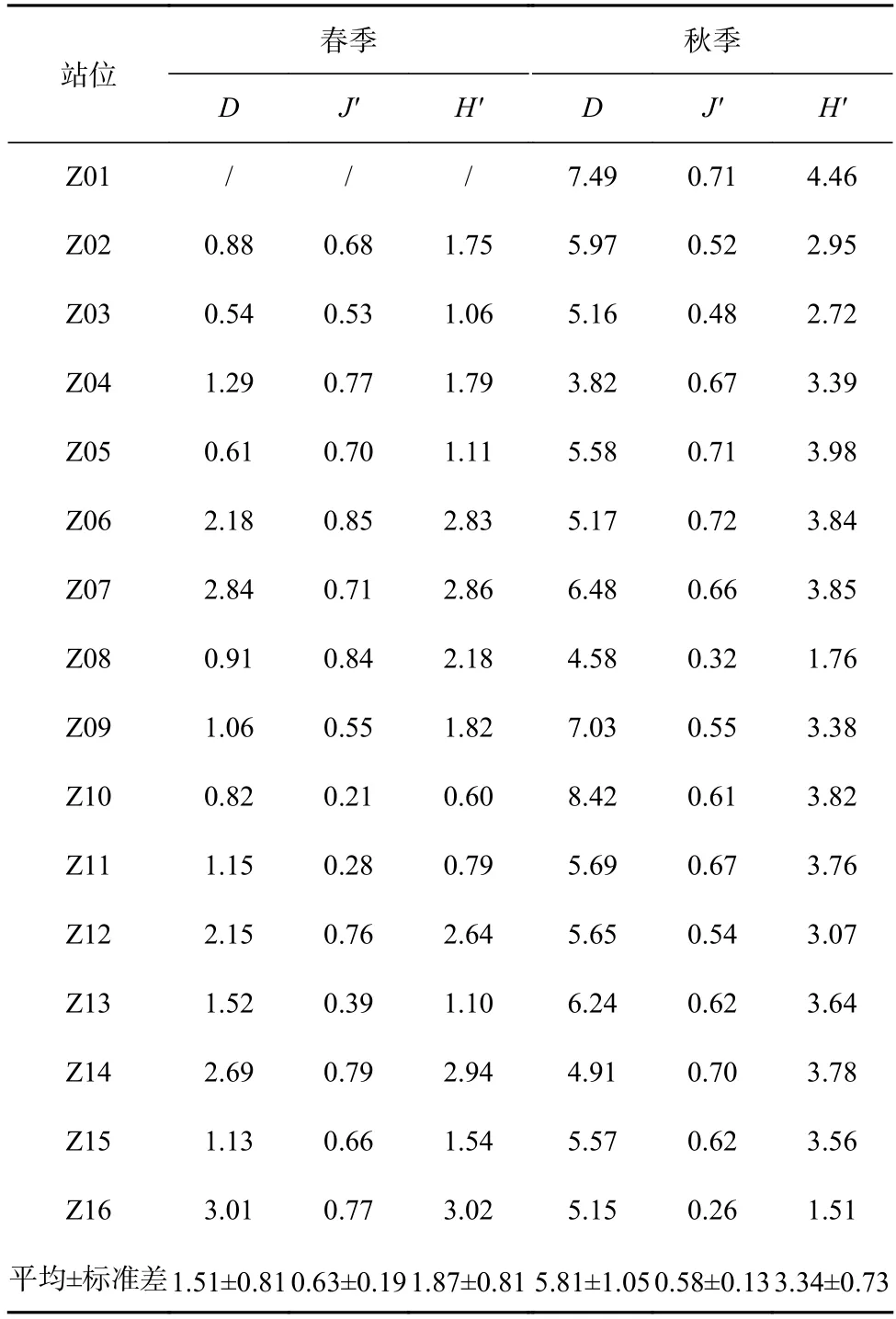
表3 调查海域各站位蟹类生物丰富度指数(D)、均匀度指数(J')和多样性指数(H')Table 3 Richness index (D),evenness index (J') and diversity index (H′) of crab in each station in the investigated sea area
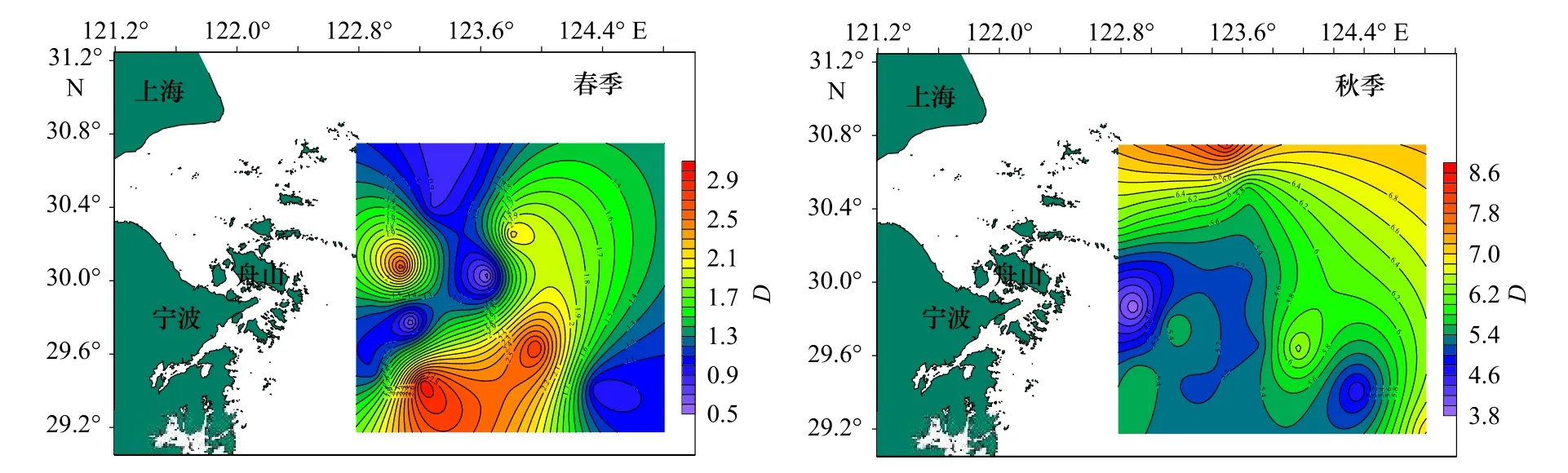
图2 调查海域蟹类丰富度指数(D)的平面分布Fig.2 Plane distribution of crab richness index (D) in the investigated sea area
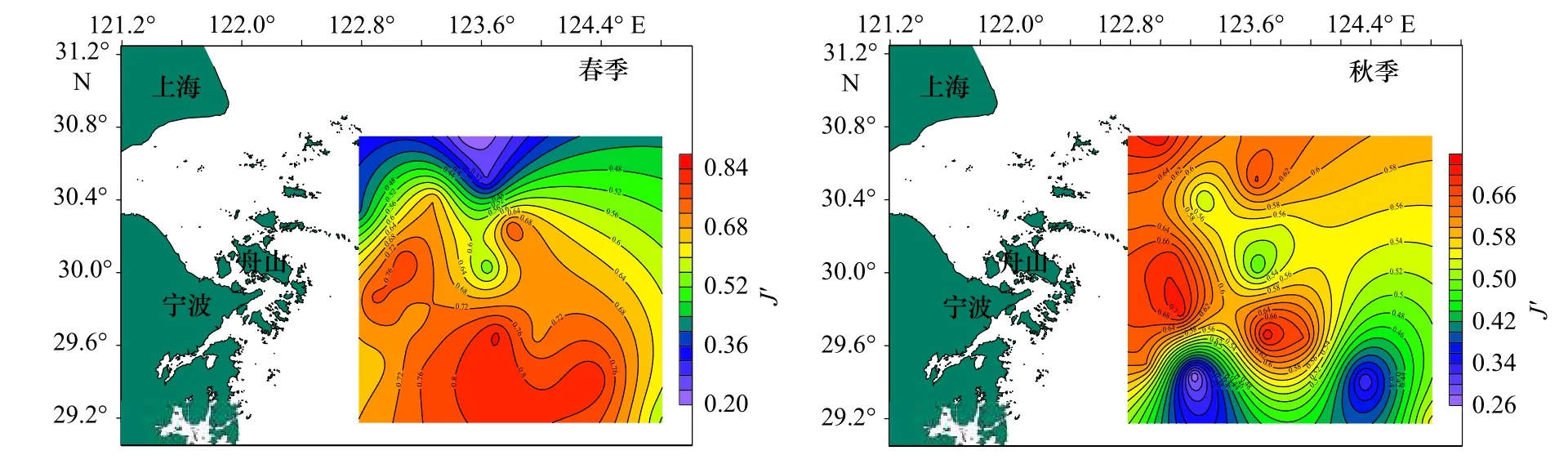
图3 调查海域蟹类均匀度指数(J')的平面分布Fig.3 Plane distribution of crab evenness index (J') in the investigated sea area
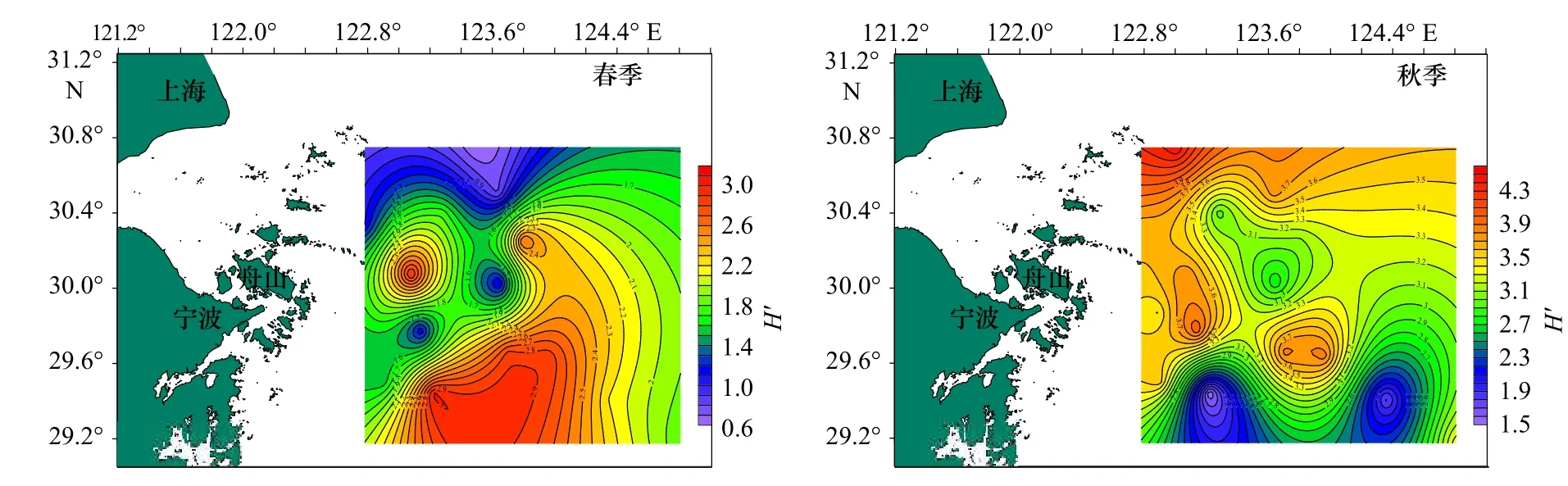
图4 调查海域蟹类多样性指数(H')的平面分布Fig.4 Plane distribution of crab diversity index (H') in the investigated sea area
从表3 中可以看出,春季,蟹类种类丰富度指数(D)分布在0.54~3.01 之间,平均值为1.51,以Z16 站位最高,Z03 站位最低;均匀度指数(J')分布在0.21~0.85 之间,平均值为0.63,以Z06 站位最高,Z10 站位最低;生物多样性指数(H')分布在0.60~3.02 之间,平均值为1.87,以Z16 站位最高,Z10 站位最低。
秋季,蟹类种类丰富度指数(D)分布在3.82~8.42 之间,平均为5.81,以Z10 站位最高,Z04 站位最低;均匀度指数(J')分布在0.26~0.72 之间,平均为0.58,以Z06 站位最高,Z16 站位最低;多样性指数(H')分布在1.51~4.46 之间,平均值为3.34,以Z01 站位最高,Z16 站位最低;总的来看,春、秋季调查海域不同站位蟹类的指数差异较大,且两个季节的不同指数最高值和最低值分别出现在不同站位。
总体来说,春、秋季生物指数平面分布季节变化明显,秋季3 项指数较高处比春季较高处均有向北部移动迹象,并且3 项指数均为北部海域高于南部海域。比较春、秋两季的生物指数,除均匀度相差不大外,丰富度及多样性均表现为秋季大于春季。
3.3 时空分布
春、秋季调查海域蟹类每小时渔获重量分布如图5所示。春季,调查海域各站位平均每小时蟹类重量渔获范围为132.6~10 875.2 g/h,平均渔获率为2 015.1 g/h,最高渔获率出现在Z09 站位,为10 875.2 g/h,Z07 站位、Z12 站位、Z10 站位等依次递减。其余站位每小时蟹类渔获重量均低于调查海域的平均值,Z05 站位最低,仅为132.6 g/h。秋季,调查海域各站位平均每小时蟹类重量渔获范围为2 619.1~32 382 g/h,平均渔获率为12 676.3 g/h,最高渔获率出现在Z16 站位,为32 382 g/h,Z01 站位、Z03 站位、Z14 站位、Z08 站位和Z05 站位等依次递减,其余站位每小时鱼类渔获重量均低于调查海域的平均值,其中以Z10 站位最低,仅为2 619.1 g/h。
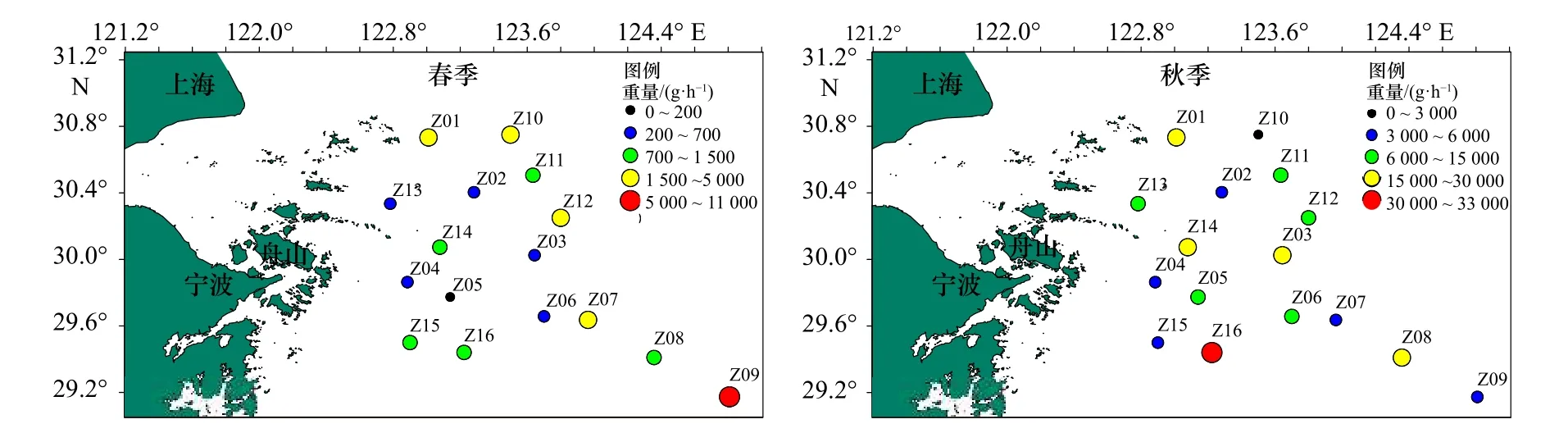
图5 调查海域各站位蟹类重量分布Fig.5 Weight distribution of crab at different stations in the investigated sea area
春、秋季调查海域蟹类每小时渔获尾数分布如图6 所示。春季,调查海域各站位平均每小时蟹类尾数渔获范围为15~4 784 ind./h,平均渔获率为502 ind./h,最高渔获率出现在Z09 站位,为4 784 ind./h,其次是Z10 站位,为1 550 ind./h,其余站位平均每小时蟹类渔获重量均低于调查海域的平均值,其中以Z01 站位最低,仅为15 ind./h。秋季,调查海域各站位平均每小时蟹类尾数渔获范围为335~4 373 ind./h,平均渔获尾数为1 147 ind./h,最高渔获率出现在Z12 站位,为4 373 ind./h,渔获率较高的还有Z01 站位、Z03 站位、Z11 站位、Z05 站位、Z04 站位和Z15 站位,其余站位平均每小时蟹类渔获尾数均低于调查海域平均值,其中以Z04 站位最低,仅为335 ind./h。

图6 调查海域各站位蟹类尾数分布Fig.6 Mantissa distribution of crab at different stations in the investigated sea area
3.4 聚类、排序分析
蟹类群落Cluster 聚类分析结果和NMDS 二维点阵图排序结果如图7 所示,春季蟹类群落Cluster聚类分析显示,各调查站位在35%相似性水平上可被分为4 个群组,第一群组为Z06 站位、Z07 站位、Z15站位、Z12 站位、Z14 站位和Z16 站位,代表性蟹类为双斑蟳、长手隆背蟹及日本英雄蟹,第二群组有Z04 站位、Z05 站位、Z11 站位和Z13 站位,代表性蟹类是双斑蟳和日本蟳,第三群组有Z02 站位、Z03 站位、Z08 站位、Z10 站位和Z09 站位,代表性蟹类是日本英雄蟹和双斑蟳,第四群组仅Z01 站位,该站位仅捕获到细点圆趾蟹1 种,数量仅有15 尾,由此可见,春季以双斑蟳分布最为广泛。

图7 蟹类群落Cluster 聚类分析和非度量多维标度分析排序Fig.7 Cluster analysis and non-metric multi-dimensional scaling ordination of crab community
秋季,各调查站位在50%相似性水平上也可被分为4 个群组,第一群组为Z08 站位和Z09 站位,代表性蟹类是三疣梭子蟹和银光梭子蟹,第二群组为Z01站位和Z10 站位,代表性蟹类是双斑蟳,第三群组为Z04 站位,代表性蟹类是三疣梭子蟹,第四群组有Z02 站位、Z06 站位和Z16 站位等11 个站位,代表性蟹类是双斑蟳和三疣梭子蟹,可见三疣梭子蟹在秋季分布最为广泛。秋季蟹类群落相似性高于春季,两季各群落代表性蟹类均有所不同。通过非度量多维标度分析结果可知,调查海域春季NMDS 胁迫系数为0.09,介于0.05~0.10 之间,说明其结果基本可信;秋季NMDS 胁迫系数为0.14,介于0.10~0.20 之间,说明其结果具有解释意义。
SIMPER 分析显示,春季对相似性贡献率较大的种类为双斑蟳、日本蟳和长手隆背蟹,贡献率分别为47.1%、17.7%、6.3%;秋季对相似性贡献率较大的种为双斑蟳、三疣梭子蟹和日本蟳,贡献率分别为45.7%、27.4%、10.7%。春、秋季蟹类群落相异性为65.5%,对相异性贡献率较大的分歧种为三疣梭子蟹,贡献率为14.2%,其余蟹类相异性贡献率均低于8%。
3.5 环境因子结果
根据同步调查所获得的温度、盐度数据,调查海域春季表层水温分布范围为14.96~18.05℃,平均为16.59℃,秋季表层水温分布范围为21.89~24.18℃,平均为22.89℃,秋季高于春季;调查海域春季底层水温分布范围为14.00~18.44℃,平均为16.87℃,秋季底层水温分布范围为21.42~23.36℃,平均为22.34℃,秋季高于春季;调查海域春季表层盐度分布范围为30.16~32.72,平均为31.51,秋季表层盐度分布范围为32.89~33.98,平均为33.44,春季与秋季相差不大;调查海域春季底层盐度分布范围为31.69~34.16,平均为33.75,秋季底层盐度分布范围为33.15~34.36,平均为33.81,平均底层盐度春季与秋季基本持平。
将调查海域蟹类群落结构与表层水温(ST)、底层水温(BT)、表层盐度(SS)、底层盐度(BS)和水深(D)5 个环境因子用CANOCO5.0 软件进行运算。CCA 排序结果如图8 所示。春季,CCA1 特征值为0.345,CCA2 特征值为0.230,CCA1 和CCA2 与环境因子相关系数分别为0.930 和0.900。蒙特卡洛显著性检验结果显示,两排序轴呈现极显著差异。两个排序轴共解释了主要蟹类组成21.36%的变异。表层水温聚集在CCA2 的负轴,与CCA2 呈较大负相关;底层水温聚集在CCA1 的负轴,与CCA1 呈较大负相关。其中,Z15 站位、Z16 站位和Z06 站位聚集在CCA1 的负轴,与底层水温呈明显负相关,说明水温是影响其分布的最主要因素;Z08 站位聚集在CCA2 的负轴,与表层盐度呈明显的正相关,说明盐度是影响该站位的主要因素。

图8 调查海域站位与环境因子典范对应分析排序Fig.8 Canonical correspondence analysis ranking of stations and environmental factors in the investigated sea area
秋季,CCA1 特征值为0.224,CCA2 特征值为0.136,CCA1 和CCA2 与环境因子相关系数分别为0.836 和0.818。蒙特卡洛显著性检验结果显示,两排序轴接近于显著差异。两个排序轴共解释了主要蟹类组成20.06%的变异。底层水温聚集在CCA1 的正轴,与CCA1 呈较大正相关,表层盐度、底层盐度及表层水温聚集在CCA1 的负轴,与CCA1 呈明显负相关,其中,Z06 站位和Z08 站位聚集在CCA1 的负轴,与表层盐度和底层盐度呈明显负相关,说明盐度对其影响较大;春、秋两季环境因子中水深对站位的影响效果较为一般。CCA 排序图的分隔模式可能反映了主要蟹类对栖息地类型的偏好或者其生态习性的适应性。
4 讨论
4.1 种类组成及优势种分析
根据本次调查结果,在舟山群岛东侧海域春、秋季共有种和两季特有种均为21 种,表明舟山群岛东侧海域种类丰富,地方性种类和季节性种类均占有一定比例,且蟹类区系组成均以广温、广盐性的暖水性种为主,这与俞存根等[1]开展的东海蟹类资源调查结果相符合,主要原因可能与调查海域受台湾暖流和浙江沿岸水交互作用形成的混合水区有关。本次调查获得的经济蟹类主要有三疣梭子蟹、细点圆趾蟹、锈斑蟳、武士蟳、红星梭子蟹(Portunus sanguinolentus)、日本蟳等,经济蟹类重量占蟹类总重量的74.1%,说明该海域可以成为人们捕捞生产蟹类的渔场。本次调查结果中,两季共同优势种仅双斑蟳1 种,一方面因为双斑蟳在东海分布最为广泛,其次双斑蟳有着较长的繁殖期,春、夏、秋季都能捕到抱卵个体,其往往在夏季大量繁殖,秋季时出现渔获高峰。而经济价值较高的三疣梭子蟹仅为秋季的优势种,其IRI 值相比春季大幅增加,主要原因是春季近岸海域水温逐步回升,大量三疣梭子蟹从外海低温深水区洄游到近岸浅水区和岛礁区进行繁殖索饵,经过春、夏季的育肥阶段,加之我国夏、秋季伏季休渔制度的保护,使得三疣梭子蟹在秋季大量出现,成为东海渔区重要的捕捞对象。本次调查中的蟹类资源优势种数量与郑献之等[23]的研究结果相比有明显减少,而与俞存根等[9]的调查结果对比,本调查中细点圆趾蟹成为东海北部附近海域的优势蟹类。优势种及经济种类明显减少,主要原因与过度捕捞和海洋环境污染有关,是否还有其他潜在影响因素,还有待进一步研究。
SIMPER 分析显示,双斑蟳、日本蟳和长手隆背蟹对春季相似性贡献率较大,主要原因是双斑蟳大多分布在黄海和东海海域,以东海分布最为广泛;日本蟳在我国4 大海区均有分布,在东海主要分布在长江口渔场,是东海蟹类的第五大优势种,主要汛期和产量高峰均在秋季;长手隆背蟹主要分布在东海和南海,数量以春、夏季最多,所以本次长手隆背蟹在春季贡献率较高;春季为三疣梭子蟹的产卵繁殖季节,经过春、夏季的索饵育肥,在秋季达到产量高峰,所以三疣梭子蟹在秋季群落结构相似性中有较大贡献。Cluster 聚类分析表明,调查海域秋季群落相似性大于春季(春季为34.1%,秋季为55.2%),也间接说明了秋季种类数和生物量水平高于春季。主要原因是秋季为三疣梭子蟹产量高峰期以及大规模的增殖放流提高了生物量,但是否还存在其他影响因素,仍有待进一步研究。
4.2 数量分布及生物多样性变化
根据本次调查结果可知,春季调查海域蟹类数量为东南部外海占比较大,西部和西南部海域较少,秋季调查海域蟹类数量相比春季有一定西移和北移现象,在调查海域中部或南部近岸较高,东南部蟹类数量较低。其原因可能是每年冬季蟹类向南向东洄游至外海深水区进行越冬,翌年春季随着水温逐步上升,蟹类再次洄游到近岸岛礁区和浅水区进行索饵产卵;夏、秋季台湾暖流到达东海北部海域,并且夏、秋季大陆降水最多,长江冲淡水及杭州湾等陆地径流达到最大,多个水团交互影响,加之夏季海域内浮游动植物丰度显著上升,提高了海域内初级生产力,也吸引了大量蟹类到群岛近岸海域进行索饵产卵活动。生物多样性结果表明,春、秋季均匀度指数(J')平均值相差不大,而秋季丰富度指数(D)、多样性指数(H')均大于春季,其主要原因可能是由于秋季台湾暖流仍然影响东海北部海域,高温、高盐水吸引大量生物前来生殖产卵,也使广温、广盐性种类所占比例有所降低,使生物多样性及丰富度显著提高。以三疣梭子蟹为代表的秋季生物多样性明显大于春季,其原因一方面是春、夏季蟹类到近海海域产卵繁殖,加之夏季我国实施伏季休渔,减少了蟹笼和刺网等网具的作业频率,另一方面浙江省早在2003 年就开始了三疣梭子蟹增殖放流活动,每年投放亲体数量达千万尾,从而提高了群体数量。
4.3 底层温度和表层盐度对蟹类群落的影响
典范对应分析结果表明,底层温度和表层盐度是影响该海域蟹类群落结构的主要环境因子。秋季平均温度和平均盐度均高于春季,究其原因可能为调查海域受台湾暖流高温、高盐水和江浙沿岸水的共同影响,使该海域温度和盐度相对其他区域较高,侯伟芬等[24]在对舟山近岸渔场海域的水温分布研究中也同样证明了秋季水温高于春季。调查海域远离大陆,加之西北部及西部设立有嵊泗列岛自然保护区和东极岛自然风景区,水质环境较好,受人类活动干扰影响较小,因此吸引大多数蟹类在此栖息生存。盐度方面,盐度受陆地径流的混合水台湾暖流的高盐水影响,以调查海域最北部Z01 站位和Z02 站位为代表,春季表层盐度分别为31.71 和31.73,底层盐度为33.47和33.61,秋季表层盐度为33.37 和33.13,底层盐度为33.61 和33.62。该规律同样适用于调查海域最东部Z08 站位及Z09 站位,同时Z08 站位及Z09 站位秋季底层盐度高达34.21 和34.26,明显具备高盐水特征,侯伟芬等[25]在对舟山近岸渔场海域盐度的调查中也指出了由于受春季汛期和夏季台风天气降水较多影响,近岸海域盐度低于远海。水深方面,春、秋两季中除Z04 站位和Z13 站位水深较浅(27 m、28 m)且渔获较少,春季Z09 站位和Z10 站位水深最深且渔获最多,其余站位平均水深均为60 m 上下,各站位水深相差不大,且水深层面与蟹类渔获量分布无明显规律,表现为无论水深多少,都有一定渔获存在;同时通过典范对应分析也可发现,水深距离两排序轴均较远,相关性不明显。该结果与丁朋朋等[26]对浙江南部近海蟹类以及郑基等[27]对浙江披山海域蟹类研究的结果有所不同,原因可能是本调查海域不仅受台湾暖流和浙江沿岸流影响,而且同时受到长江冲淡水及杭州湾入海水的影响,导致海域内盐度有所下降,使盐度对蟹类群落结构影响的相关性随之降低。
4.4 蟹类生物对生态系统的作用
绝大部分蟹类属杂食性和肉食性,少部分蟹类为植物食性,还有一部分蟹类摄取泥沙中的沉积物或水体中的悬浮物,在维持生态系统平衡和促进能量循环及物质流动中发挥着重要作用。蟹类不但可作为黄姑鱼(Nibea albiflora)、褐菖鲉(Sebastiscus marmoratus)等高营养级鱼类的饵料来源,还可捕食包括贝类、浮游生物和小型甲壳类在内的底栖生物,其中三疣梭子蟹和日本蟳还会捕食小型鱼类、枪乌贼和多毛类等,食物范围也较广。此次调查海域岛礁众多,海域地处副热带季风气候区,受大陆径流和台湾暖流等水系共同影响,浮游生物、底栖生物和游泳动物等种类繁多,是多种海洋生物产卵、繁殖、索饵和洄游的重要栖息地。蟹类生物作为水生生物食物网中重要的一环,既是生产者(鱼类饵料生物)又是消费者(捕食底栖生物),对群落结构的稳定性起着至关重要的作用。在鱼类资源严重衰退的今天,包括蟹类在内的甲壳类作为鱼类资源的重要补充,其地位逐渐被人们所重视并成为人们重要的捕捞对象。与此同时,我们还应进一步加强对蟹类资源的保护,摸清蟹类生物繁殖与生长等生态学特性,掌握其种群变动规律,以期实现蟹类资源的可持续利用和生态环境的可持续发展。
致谢:颜文超、韩磊等同志参加海上样品采集和室内种类鉴定,戴冬旭、张文俊、辛艺、孙蓓蓓、刘坤、于南京、张佩怡等同志参加室内种类鉴定和数据处理,谨致谢枕。
猜你喜欢
中国检察官(2022年20期)2022-11-07
中国水产(2021年6期)2021-07-01
河南水产(2020年1期)2020-06-29
水产科学(2020年3期)2020-05-25
农家之友(2020年9期)2020-04-21
浙江海洋大学学报(自然科学版)(2019年5期)2019-03-21
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
现代营销·经营版(2017年3期)2017-05-17
作文周刊·小学五年级版(2016年1期)2016-06-29
