
应用稳定同位素技术研究江苏近海食物网营养结构的季节性变化
2022-02-18 12:57陆亚楠张瑞张虎张硕高世科
海洋学报 2022年2期
陆亚楠,张瑞,张虎,张硕, ,高世科*
(1.上海海洋大学 海洋科学学院,上海 201306;2.江苏省海洋水产研究所,江苏 南通 226007;3.长江口水生生物资源监测与保护联合实验室,上海 200922)
1 引言
理解水生生态系统中的食物网结构及生物群落构成是生态学领域的一个重要方向[1-2]。研究食物网中结构、功能及营养相互作用,有助于了解生态系统生物多样性、稳定性和功能[3-5]。几十年来,δ13C、δ15N 稳定同位素技术多被用于分析海域生态环境中的能量流动和物质交替变化,在构建海域食物网、确定营养生态位和鱼类食性特征方面取得了重大进展,也为研究海洋生物营养结构和动力学方面提供了重要见解[6-7]。稳定同位素比率(δ13C/δ15N)可以代表生物个体在食物网中的所有营养途径[8],而同位素生态位反映了消费者-资源之间的营养相互作用,可用于反映生物营养结构的时空分布特征[9-10]。此外,基于稳定同位素值量化的各项生态指标不仅能够用于描述物种和群落水平上的生态位宽度和重叠程度,还能评估不同因素对物种生态位宽度的影响,从而表征食物网的多样性和复杂性[11-13]。
江苏近海的渔业资源丰富,是我国重要的渔场。近年来,随着沿海商业渔业的发展,江苏近海局部地区海域污染状况愈发严重[14]。不断发展的渔业装备技术和生产水平加剧了渔业过度捕捞,使得鱼类种群数量持续下跌,自然生境遭到破坏,对海洋生物的生存环境造成了严重威胁[15-16]。因此,急需开展对该地区主要鱼类群落营养结构时空变化规律的调查,以便及时制定相应政策和管理措施。
本文利用稳定同位素技术研究了江苏近海春、夏、秋3 季主要海洋生物的δ13C、δ15N 和营养级特征,对比分析3 个季节食物网的营养结构和变动规律,结合食物网量化指标,从物种和群落水平上反映渔业资源现状及季节变动情况,以期为进一步评估江苏近海渔业生态系统的营养结构变化奠定基础,为保护沿海海域生态系统以及渔业资源的养护和管理提供一定科学依据和参考基础。
2 材料与方法
2.1 样品采集
本研究分别于2017 年5 月7-24 日(春季)、8 月3-23 日(夏季)和10 月4-19 日(秋季)在江苏近岸海域(31.794°~35.583°N,119.301°~122.243°E)通过拖网渔船进行样品采集。3 个季节分别设置站点23 个、17 个和30 个(图1)。样品的采集和处理方法遵照《海洋监测规范第6 部分:海洋生物调查》(GB/T 12763.6-2007)[17],浮游动物样品用Ⅰ型浮游生物网定点垂直拖网采集,垂直拖网平均深度为10 m,在海水中清养1~2 h 排空胃含物,然后置于1 L 聚乙烯瓶中。其他生物均采用单船有翼单囊底拖网(规格40 m×94 m或40 m×49.3 m),平均拖速为3.0 kn,每网拖行近1 h。将生物样品鉴定到种(未能鉴定到种的做好挂牌登记),测量体长、体重,分类冷冻保存后带回实验室。
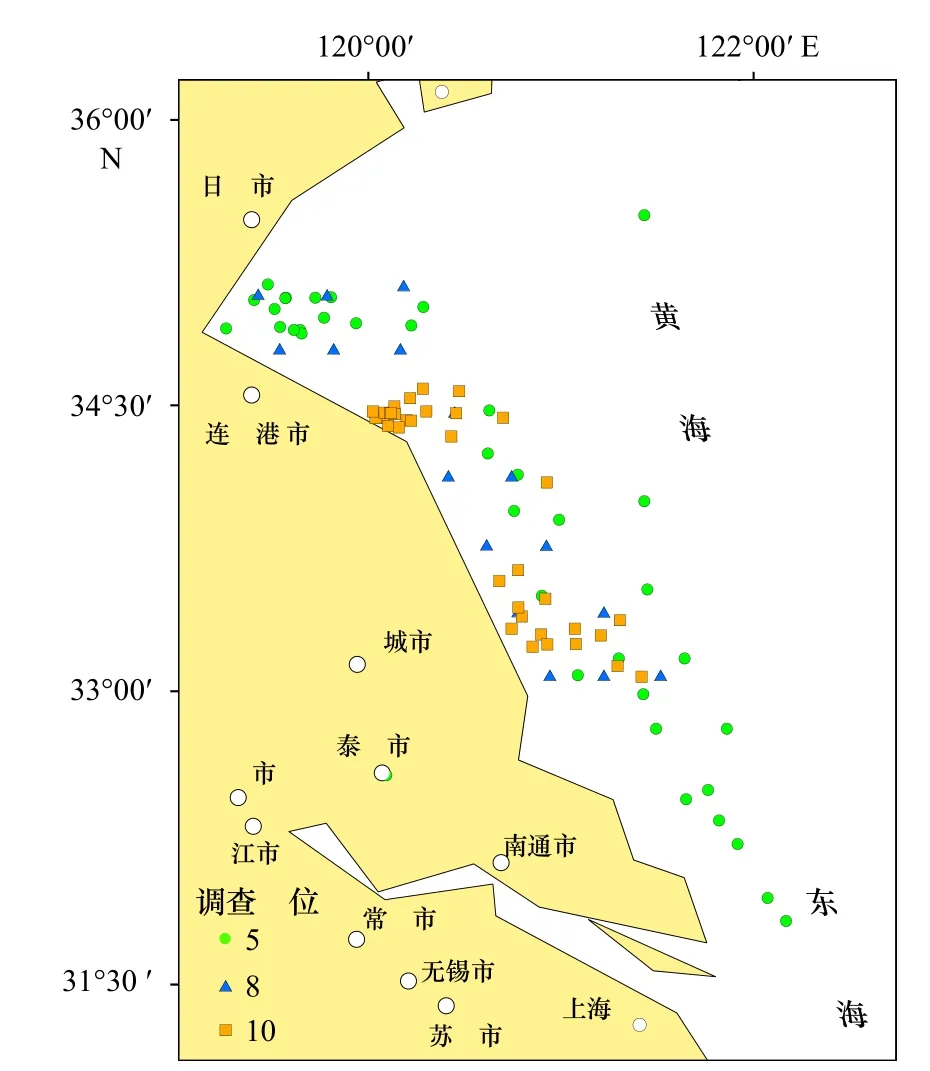
图1 江苏近海采样点位置Fig.1 Locations of sampling sites in the coastal waters of Jiangsu Province
2.2 稳定同位素分析样品预处理
浮游动物用少量蒸馏水反复清洗,去除混杂在其中的杂质,然后置于清洁培养皿中挑选。其他生物每个物种选取3 个大小相近的个体(对于采样数量少于3 个的种类,则选取全部个体),鱼类取第一背鳍下方肌肉、虾类取腹部肌肉、蟹类取第一鳌足肌肉、头足类取腕部肌肉、腹足类去壳取肌肉、双壳类取闭壳肌、中国毛虾(Acetes chinensis)因个体小,选取10 尾相近个体混合、马尾藻(Sargassum)取其叶片。所有样品用去离子水洗净后放入离心管,-20℃冷冻保存。后续分析前,用冷冻干燥机(Christ 1-4α)干燥32 h,并用混合型球磨仪(Retsch MM440)研磨成粉。
研究发现,对脂肪含量不同的组织样品进行δ13C 分析时,可能产生较大偏差[18]。因此,本研究预先使用三氯甲烷和甲醇(体积比为2∶1)对肌肉组织样品进行脱脂处理,然后使用锡杯包被1.0 mg 送入稳定同位素质谱仪(IsoPrime 100,Isoprime Corporation,Cheadle,UK)进行分析,稳定同位素测定在上海海洋大学摄食生态学实验室进行。
2.3 稳定同位素分析
碳、氮稳定同位素值分别用国际通用的标准物质VPDB 和大气氮作为参考标准。结果分别用δ13C和δ15N 表示,其计算公式为
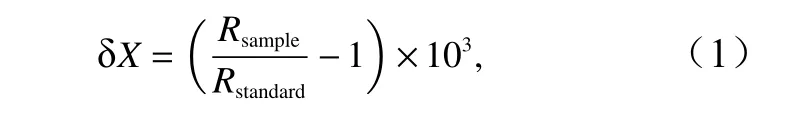
式中,X为13C 值或者15N 值;R为13C 值和12C 值或者15N值和14N 值的比值;Rsample为样品的R值;Rstandard为标准样品的R值。为保证实验结果的精度和准确度,每10 个样品,放入3 个标准品。根据氮稳定同位素比值可以计算生物的营养级(Trophic Level,TL),公式为
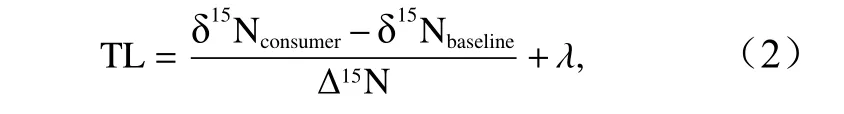
式中,δ15Nconsumer为消费者的氮稳定同位素比值;δ15Nbaseline为基线生物的氮稳定同位素比值;λ表示基线生物所处的营养级,本文选择春季采集沟纹鬘螺(Phalium strigatum)作为基线生物,其营养级定为2,原因是沟纹鬘螺的δ15N 值与毛蚶(Scapharca subcrenata)、紫贻贝(Mytilus edulis)比较接 近[19-20];δ15Nbaseline为沟纹鬘螺的氮稳定同位素比值的平均值;Δ15N表示营养级之间氮稳定同位素的富集度,这里取2.5‰和3.4‰的平均值,其中2.5‰是蔡德陵等[19]通过喂养实验得到的鳀鱼(Engraulis japonicus)与饵料间的氮稳定同位素富集值,而3.4‰出自Post[21]的研究结果。
数据分析及制图采用Origin 2020 和R 语言中的SIBER 程序包。根据Layman 等[11]提出的碳氮稳定同位素二维坐标系代表群落营养结构的方法,选取NR、CR、TA、CD、NND、SDNND 6 种参数来反映生物群落营养结构的季节变化情况。其中NR 为食物网中δ15N 值的范围,表示消费者营养层次;CR 为δ13C 的最大差值,表示食物来源的多样性;TA 为食物网营养结构总面积,代表生物多样性水平;CD 为平均离心距离,代表群落整体的多样性水平;NND 为平均最邻近距离;SDNND 为平均最邻近距离标准差,反映营养冗余程度。考虑到TA 可能因个别数据产生较大偏差,使用贝叶斯标准椭圆面积(SEA)可以减少样本误差,若样本数小于30,则比较标准椭圆校正后的面积(SEAC)[22-23]。
2.4 数据处理
通过ArcGIS 10.3 软件绘制调查海域站位图;根据聚类分析结果,运用OriginPro 2020 软件构建连续营养谱;数据处理和表格制作通过EXCEL 2020 完成;利用SPSS 25.0 软件进行显著性t检验。
3 结果
3.1 春、夏、秋3 季生物的碳、氮稳定同位素特征
春季共采集到生物样品31 种,其中鱼类16 种、甲壳类5 种、头足类3 种、底栖动物6 种、浮游生物1 种(附表A1)。其δ13C 值范围为-24.47‰(马尾藻)~-14.50 ‰(黄魟Dasyatis bennettii);δ15N值范围为6.86‰(马尾藻)~(14.98±0.49)‰(中国花鲈Lateolabrax maculates)(附表A1,图2)。夏季共采集到生物样品32 种,其中鱼类17 种,甲壳类7 种,头足类3 种,腹足类2 种,双壳类3 种。δ13C 值的范围为-19.86‰(江珧Atrina pectinata)~-14.44‰(脊尾白虾Exopalaemon carinicauda);δ15N值范围为5.79‰(江珧)~(12.54±3.50)‰(赤鼻棱鳀Thryssa kammalensis)。秋季共采集到生物样品24 种,其中鱼类11 种,甲壳类6 种,头足类3 种,腹足类1 种,双壳类2 种以及浮游动物1 种。δ13C 值的范围为(-22.17±4.90)‰(鲻Mugil cephalus)~(-14.21±0.36)‰(短吻舌鳎Cynoglossus abbreviatus);δ15N 值范围为6.30‰(浮游动物)~(14.60±0.51)‰(中国花鲈)。总体上,鱼类δ13C 值的跨度最广,最低的是鲻,为(-22.17±4.90)‰,最高的是短吻舌鳎,为(-14.21±0.36)‰,δ15N 值范围为(8.43±0.38)‰(棘头梅童鱼Collichthys lucidus)~(14.79±0.19)‰(中国花鲈)。
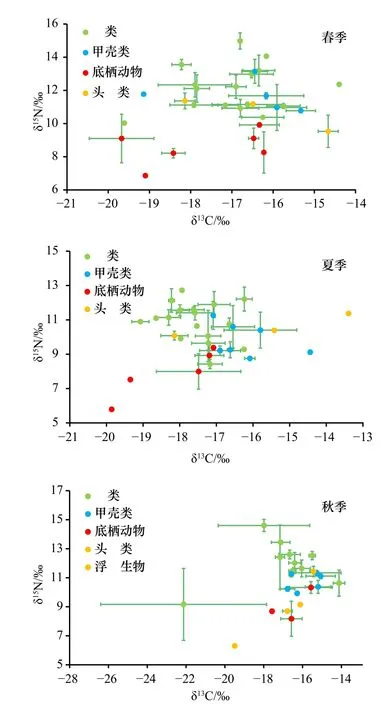
图2 春、夏、秋3 季江苏近海生物δ13C、δ15N 值的双坐标图Fig.2 The biplot of δ13C,δ15N of organisms from the coastal waters of Jiangsu Province in spring,summer and autumn
3.2 江苏近海营养结构的季节性变化特征
配对样本t检验的结果表明(表1),2017 年春、夏、秋3 个季节江苏近海海洋生物的δ13C 值和δ15N值均没有显著性差异(p>0.05)。
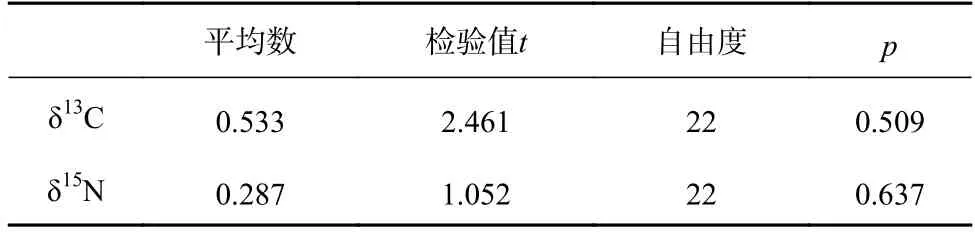
表1 春、夏、秋3 季生物的δ13C、δ15N 值差异性分析Table 1 Difference analysis in δ13C and δ15N values for organisms in spring,summer and autumn
由于鲻的δ13C 值与其他消费者存在明显差别,对营养结构影响过大,为了更准确地反映群落营养结构的季节变化,本研究选择除鲻和浮游动物外的所有消费者进行空间生态位的计算,结果见图3,参数见表2。不同类群对生物资源利用情况不同。春季,甲壳类对生态空间利用率较其他类群高,而底栖动物由于整体营养级较低,与其他类群生态空间重叠不明显;夏季,生物的生态空间利用情况较春季一致,但重叠程度不如春季明显;秋季,不同类群生物的生态空间利用率较小,头足类、甲壳类和底栖动物的生态空间重叠程度较为明显。此外,不同季节相同类群生物对生态空间利用情况也不同。总体上,3 个季节鱼类的生态空间利用率最高,最低为底栖动物。
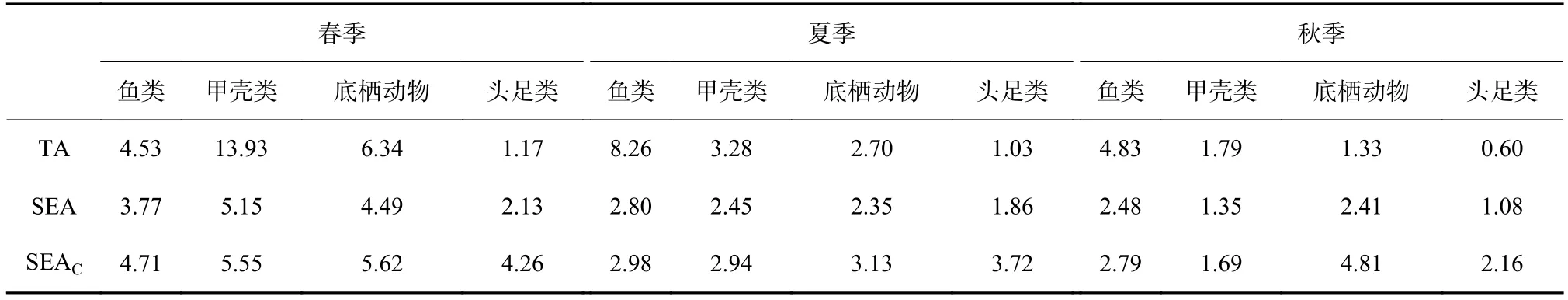
表2 春、夏、秋3 季生物的TA、SEA 和SEAC 值Table 2 TA,SEA and SEAC for organisms in spring,summer and autumn

图3 春、夏、秋3 季江苏近海生物营养框架Fig.3 Stable isotope bio-plotting of the trophic structure in the coastal waters of Jiangsu Province in spring,summer and autumn
通过R 语言对春、夏、秋3 个季节营养结构的6 种生态位指标进行计算,结果见图4。结果显示,春季的营养级长度最长,其次为夏季、秋季;在生态位多样化程度和生物对生态空间利用方面由大到小依次为春季、夏季、秋季;平均营养级多样性和营养冗余程度也均呈现出春季最高,秋季最低。
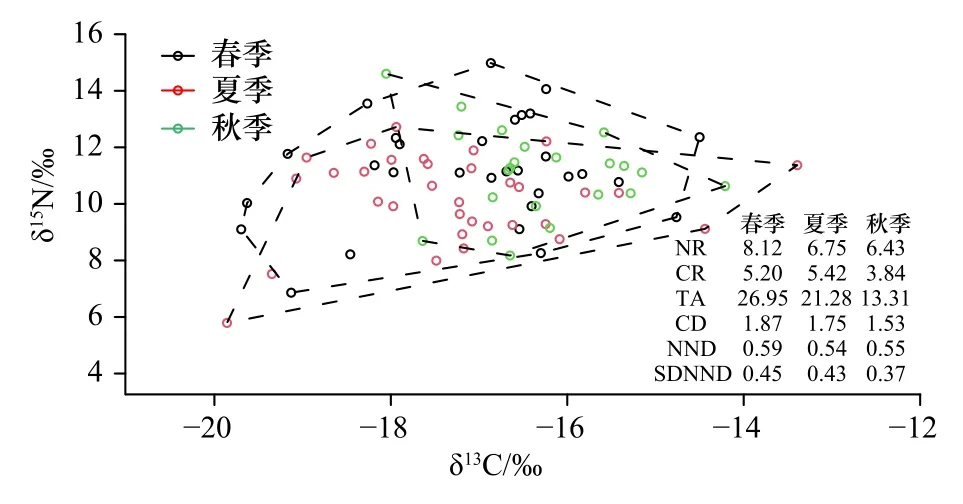
图4 春、夏、秋季江苏近海生物群落稳定同位素参数值Fig.4 Community-wide metrics based on stable isotopic ratios for organisms from coastal waters of Jiangsu Province in spring,summer and autumn
3.3 春、夏、秋3 季江苏近海生物的营养级特征
春季江苏近海海洋生物的营养级范围为1.52(江珧)~4.28±0.17(中国花鲈),营养层次长度为4 级(图5)。其中,底栖动物属于第1~2 营养级,营养级最高的是脉红螺(Ostrea denselamellosa)(2.56±0.04);头足类属于第2 营养级,火枪乌贼(Loligo beka)的营养级达到3.05±0.16;甲壳类属于第2~3 营养级,口虾蛄(Oratosquilla oratoria)营养级最高,为3.66±0.25,其次是中国毛虾(3.19)和日本蟳(Charybdis japonica,3.16±0.06);鱼类属于第4 营养级,营养级范围为2.6(日本鳀)~4.29±0.17(中国花鲈)。而夏季的营养层次长度较短,为3 级,营养级范围为1.25(江珧)~3.52(长蛇鲻,Saurida elongata)。其中,多数底栖动物属于第2 营养级,仅有毛蚶和江珧为1.84 和1.25。甲壳类中,仅有鹰爪虾(Trachypenaeus curvirostris)属于第3 营养级(3.11±0.28),其余均为第2 营养级。鱼类属于2~3 营养级,范围从2.06±0.10(棘头梅童鱼)~3.52(长蛇鲻)。秋季的营养层次长度亦为4 级,营养级范围是1.34(浮游动物)~4.15±0.14(中国花鲈)。其中,浮游动物属于第1 营养级,多数底栖动物、头足类和甲壳类属于第2~3 营养级,只有毛蚶为1.97±0.41。鱼类中仅有中国花鲈营养级最高,为4.15±0.14,其余鱼类均属于2~3 营养级。
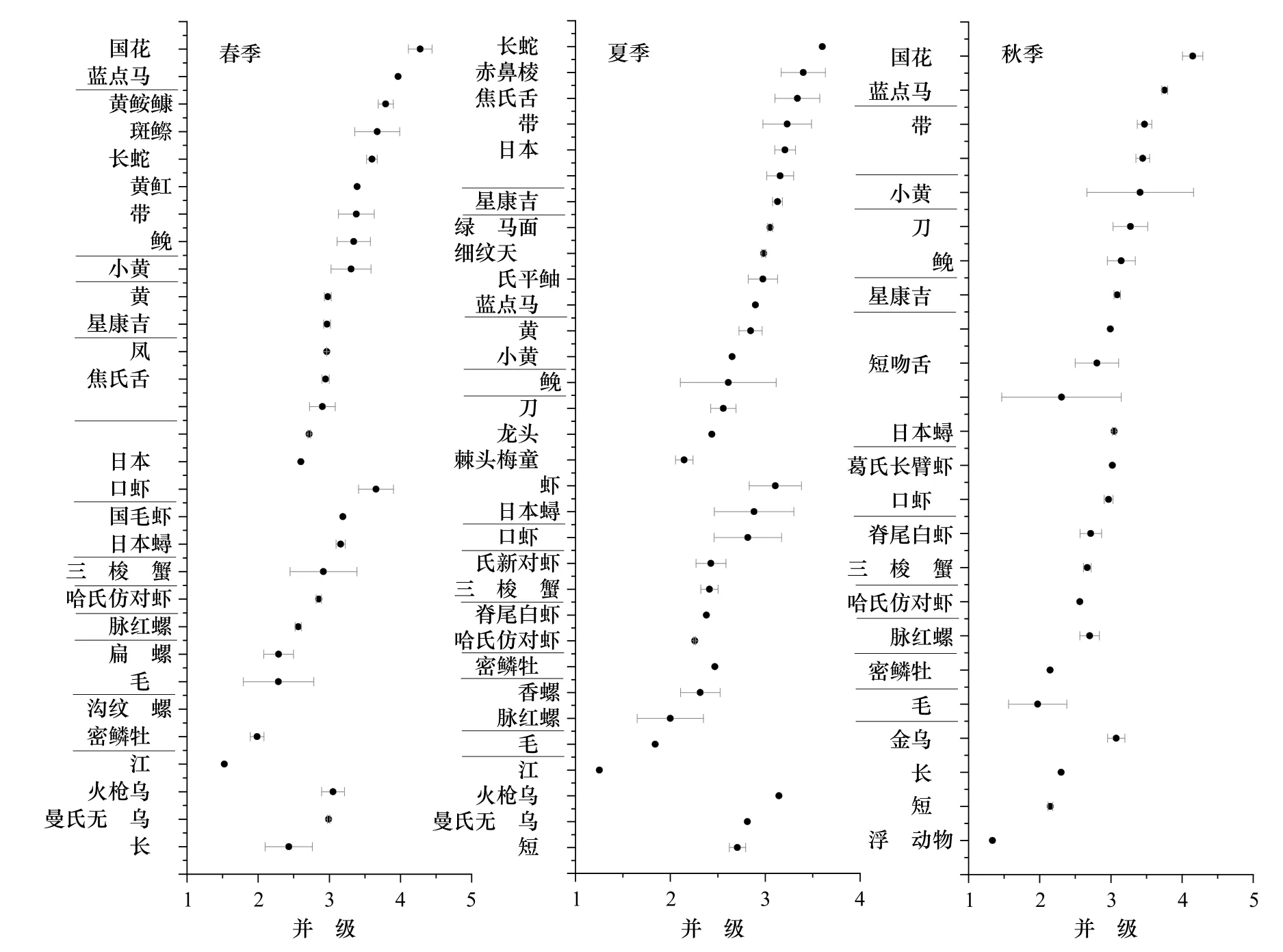
图5 春、夏、秋3 季江苏近海生物营养级对比Fig.5 The comparison of trophic level of organisms from coastal waters of Jiangsu Province in spring,summer and autumn—代表3 个季节中共同出现的物种——represents the species occured in three seasons
4 讨论
4.1 春、夏、秋3 季江苏近海生物的δ13C、δ15N 特征
本研究基于渔业资源调查采集生物样品,应用稳定同位素技术构建江苏近海的食物网营养结构。消费者的δ13C 值可以指示其有机质和能量来源。本研究不同季节食物网中,生物的δ13C 值没有明显差异(p>0.05),这与高世科等[24]的研究中春季吕泗渔场主要生物的δ13C 值一致,说明各季节的有机质来源可能差异不大。春、秋季鲻的δ13C 值跨度较大,远低于其他消费者,说明其在食物网中可能存在个体特殊化现象[25],即在食物网中,同一物种的不同个体始终对不同的生产力来源进行摄取。生物的δ15N 值可以指示其营养级,在江苏近海食物网中,不同季节生物的δ15N值也没有明显差异(p>0.05),说明该食物网营养结构稳定,各季节的营养层次基本一致。此外,本研究各个季节采样站位分布不均匀。这种空间差异性可能会影响结果,可能原因是不同区域受人类活动影响和水体富营养化程度不同[26]。所以在今后的采样设计中应该尽可能选择相同区域的生物样品进行对比分析。
4.2 江苏近海食物网营养结构的季节性变化
3 个季节中,春季鱼类与甲壳类、头足类的生态位重叠程度较高,说明这些消费者可能在生活环境和食物溯源上存在一定相似性(图3)。江苏近海拥有多个重要的过路渔场(如吕泗渔场、海州湾渔场等),是多种经济洄游性鱼类的必经之地[27-28]。此外,春季饵料缺乏,鱼类需要大量摄食以保证生存和繁殖[29]。在一定程度上,这可能是导致春季鱼类食性较为广泛的原因。春季底栖动物的δ13C 值跨度范围较大,亦可说明春季的食物来源分布广泛。并且,秋季底栖动物的生态位与头足类部分重叠,而春、夏季不与任何类群重叠(图3)。这不仅与种类、生物量有关,还与生物体大小相关[30]。研究表明,随着生物体的增长,δ13C 和δ15N 会不断富集,而摄食器官大小和饵料选择的变化也会引起同位素的季节性变化[30-31]。本研究还显示,夏季鱼类的TA 值为8.26,SEA 为2.80,说明夏季鱼类种类多样性较高,但生态位分化程度低,种间竞争不激烈,同一现象还出现于春季的甲壳类(TA:13.93,SEA:5.15)(表2)。这种生态位的空间差异可能是内外因素共同导致,比如食性偏好,亦或是区域间复杂的环境因素[32-33],因此还需要深入研究。
位于营养结构边界的生物,对于维系该食物网的稳定具有重要意义[20]。本研究中,处于营养结构端点位置的生物包括顶级捕食者(如中国花鲈、鮸(Miichthys miiuy)),浮游食性的消费者(如赤鼻棱鳀、中国毛虾)和底栖消费者(如黄魟、脊尾白虾等),与纪炜炜等[34]的研究结果一致。江苏近海3 个季节生物群落的稳定同位素参数存在差异。春季NR 和TA 值最高,说明春季消费者的营养层次和群落多样性更高。当饵料充足时,摄食生物对特定食物会有选择性,食源多样性CR 相应就会减少[34]。夏季CR 值高于春、秋两季,说明夏季消费者的食物来源更加丰富,食源多样性更高。春季和夏季的CD 值差别很小,说明食物网营养多样性在春夏季比较稳定。秋季NR、CR、TA 值最低,说明秋季食物网结构单一,群落多样性水平最低。在海洋生物群落中,饵料生物数量的变动引起消费者的竞争关系发生改变。若饵料生物数量减少,消费者对于食物的激烈竞争导致生态位重叠,群落营养冗余程度增加[20]。本研究中,3 个季节NND 值十分接近,说明各季节营养冗余度差别不大。SDNND值在秋季略低,说明秋季食物网中营养生态位分布较广。鉴于本研究只从分析季节性特征来反映江苏近海海域的营养结构特征,将来应该从方法比较以及时间尺度上来进一步阐明该海域的营养结构变化。生物营养级的差异取决于多个因素,比如生物体规格大小、Δ15N 和基线生物的选取[34-35]。本研究3 个季节同种生物的营养级差异均不显著(p>0.05)(图5)。除了银鲳(Pampus argenteus)、星康吉鳗(Conger myriaster)和密鳞牡蛎(Ostrea denselamellosa),同种生物的营养级最低值均出现在夏季,这与夏季捕获的生物规格较小有关(附表A1)。此外,为了统一化比较,本研究3 个季节的所有个体都被视为具有相同的基线生物,但基线生物也存在季节性差异。如季节性水温变化会影响浮游植物的光合作用及其对氮元素的吸收和分馏,进而影响作为基线生物的浮游动物的δ15N 值[35]。因此,在估算时空尺度上消费者的营养状况时,应考虑不同或多个基线的选择。
5 结论
本文利用稳定同位素技术研究江苏近海不同季节主要海洋生物的δ13C、δ15N 值特征。结果表明,各季节的食物网结构稳定,一些消费者的营养生态位出现不同程度的重叠现象。其中,春季的食物来源较为广泛,生物营养层次和群落多样性更高;夏季消费者的食物来源更加丰富,但生态位分化程度较低;秋季的食物网结构比较单一,但营养生态位分布较广。本研究结果为进一步评估江苏近海渔业生态系统的营养结构奠定基础,也为江苏近海渔业环境保护和渔业资源的恢复与管理提供一定科学依据和参考基础。
附录
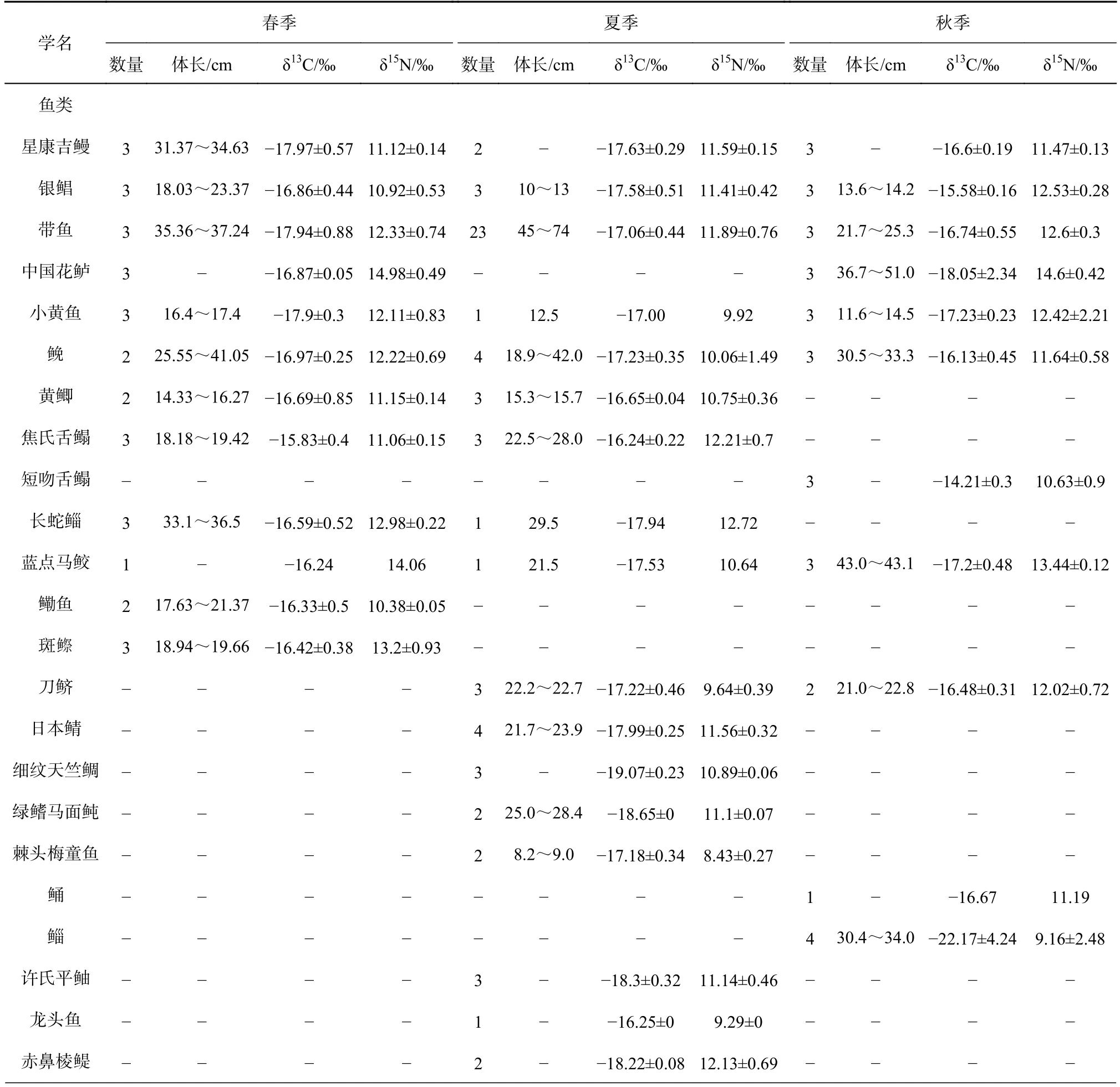
表A1 春、夏、秋3 季江苏近海主要生物数量、体长及碳、氮稳定同位素Table A1 The number,mean length and the δ13C and δ15N values of organisms in the coastal waters of Jiangsu Province in spring,summer,autumn
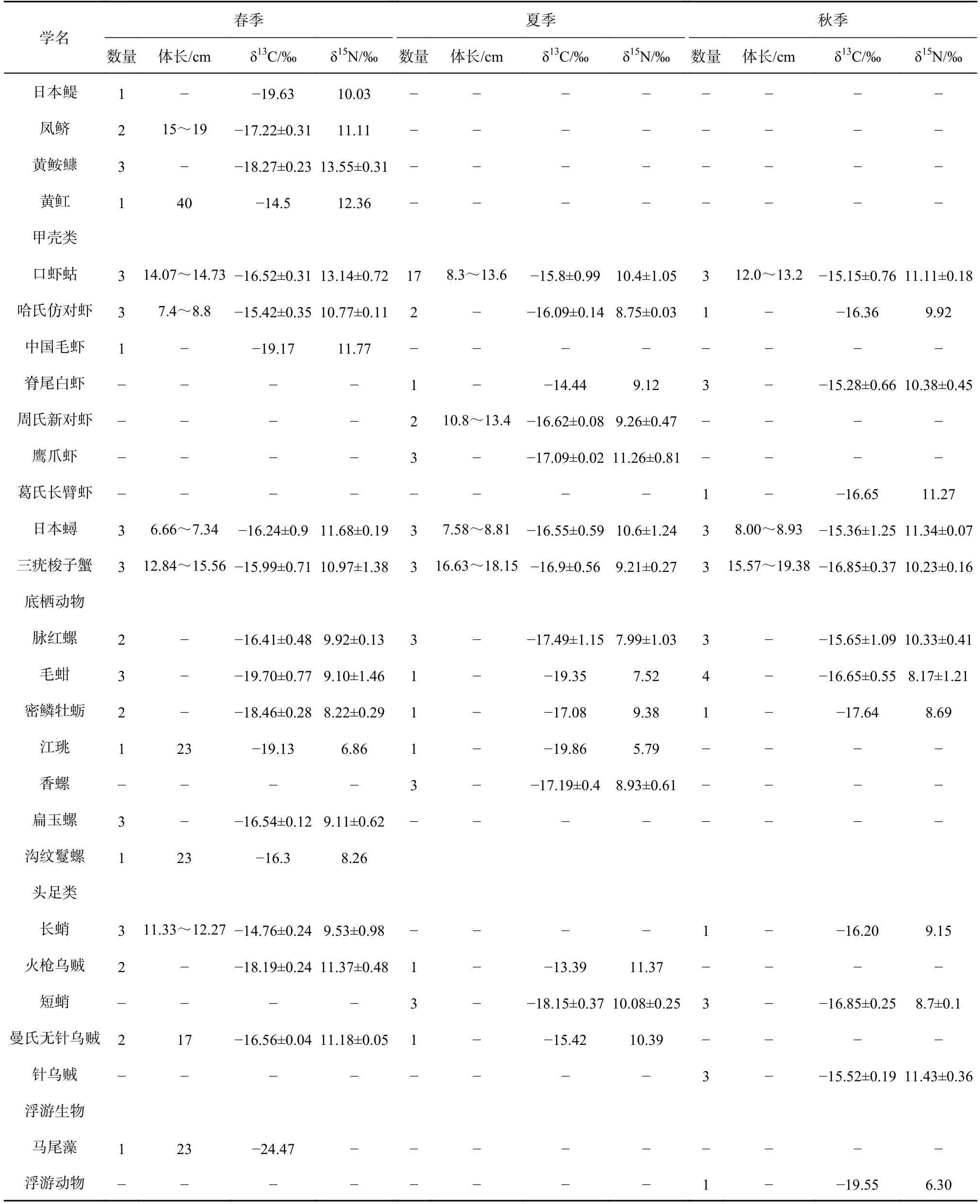
续表 A1
猜你喜欢
儿童时代·幸福宝宝(2020年9期)2020-09-08
快乐作文(1.2年级)(2019年3期)2019-09-10
英语文摘(2019年5期)2019-07-13
幼儿画刊(2018年10期)2018-10-27
分析化学(2017年5期)2017-06-21
探索科学(2017年5期)2017-06-21
小雪花·成长指南(2016年2期)2016-03-16
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
同位素(2014年2期)2014-04-16
