
不同盐度对生物絮团、对虾生长以及酶活性的影响
2020-05-25 03:10:52秦海鹏杨世平廖栩峥胡世康赵吉臣何子豪孙成波
水产科学 2020年3期
秦海鹏,杨世平,王 博,廖栩峥,胡世康,赵吉臣,何子豪,孙成波
( 广东海洋大学 水产学院,广东 湛江 524088 )
目前,水产养殖业正在走向高密度集约化、工厂化的养殖模式,在此模式下,养殖水体水质严重恶化,危害养殖对虾的健康,同时高密度养殖病害暴发日益频繁,养殖废水的排放对生态环境也造成了严重污染[1]。生物絮凝技术由以色列专家首先应用于水产养殖业之后,已经在调控水质、切断水产病害传播等方面展现出了巨大优势,已成为行业研究的热点[2],生物絮体的核心是菌胶团细菌和丝状细菌,它们通过分泌的胞内和胞外聚合产物等, 将异养细菌、硝化细菌、真菌、藻类、原生动物、有机聚合体、胶体、阳离子和碎屑等黏附在一起,形成结构多样化的絮状物[3-4]。生物絮团可以通过微生物的同化作用和硝化作用有效去除水体中的氨氮、亚硝态氮,实现水质的净化[5-6]。
凡纳滨对虾(Litopenaeusvannamei)为广盐性海水虾类,近年来在淡水、半咸水和海水中都进行了大规模养殖。目前,关于盐度对虾类生物絮团养殖影响的研究主要集中于盐度突变方面[7],而盐度对凡纳滨对虾影响的研究主要集中于凡纳滨对虾生存、生长、能量收支和渗透压调节等方面[8-11]。已有研究表明,生物絮团水体中氮元素的循环受到盐度的影响;生物絮团中的生物类群和组成结构,也受到盐度的影响,从而影响到对虾养殖水体的水质、对虾生长和代谢等[12]。但盐度对凡纳滨对虾生物絮团长周期养殖的影响尚未见相关报道。笔者在不同盐度下进行凡纳滨对虾的生物絮团养殖试验,旨在探究不同盐度对凡纳滨对虾生理因子的影响,以及生物絮团在不同盐度下氮元素转化的规律,探究生物絮团养殖凡纳滨对虾的适宜盐度,为凡纳滨对虾生物絮团海水养殖模式的推广提供参考。
1 材料与方法
1.1 试验地点
广东海洋大学水产学院东海岛海洋生物研究基地。
1.2 试验材料
试验在设有防逃网的0.3 m3的蓝色塑料桶中进行,用曝气处理后的自来水稀释消毒海水来调节盐度,用200目的筛绢网将生物絮团自养殖池中捞取至调配好盐度的海水中。凡纳滨对虾在不同盐度组内经过7 d的适应期,体长(7.7±0.63) cm、体质量(6.15±1.15) g。
1.3 试验方法
试验设置5个盐度梯度:10、15、20、25、30,每个梯度3个平行,饲养密度为500 尾/m3,生物絮团初始量为20 mL/L,试验周期30 d。试验过程中24 h连续充气。试验从第15 d起每日添加5 mg/L的碳酸氢钠来调节pH和总碱度。
1.4 水质和生长指标
试验期间,每5 d测定一次总碱度、pH、硝态氮、亚硝态氮、氨氮。在试验开始和试验结束时测定对虾的体长和体质量。
氨氮的测定采用靛酚蓝分光光度法;硝态氮的测定采用锌—镉还原分光光度法;亚硝态氮的测定采用盐酸萘乙二胺分光光度法;总碱度的测定采用酸碱滴定法[13]。
1.5 消化和免疫指标测定
试验结束时进行取样,取样时随机挑选处于蜕壳间期的对虾。取其肝胰脏、肌肉组织分别放入样品管中,准确称量质量后迅速投入液氮冷冻,再转移到冰箱-80 ℃保存,以上操作均在冰盘中进行。取保存的组织样品,加入9倍体积预冷的对虾生理盐水,在冰浴中研磨匀浆,然后4 ℃,5000 r/min离心10 min,取上清液用于对虾消化和非特异性免疫指标的测定。所有酶活力及蛋白含量的测定均采用南京建成生物研究所研制的检测试剂盒,按照操作步骤进行操作。
1.6 数据分析
所有数据采用Excel 2016软件统计分析,用SPSS 19.0软件在0.05水平上进行显著性差异分析。
2 结果与分析
2.1 不同盐度下凡纳滨对虾生长指标的差异
凡纳滨对虾在20盐度组盐度下体长的增长率最大,为20.00%,净增体长1.54 cm,15盐度组体长增长率最小,为15.58%,净增体长1.20 cm,达15.58%。各盐度下终末体长差异不显著(P>0.05)。20盐度组和25盐度组的对虾质量的增长率最大,为70.73%,净增质量4.35 g,15盐度组的体质量增长率最小,体质量增长3.09 g,达50.24%,各盐度间差异不显著(图1)(P>0.05)。
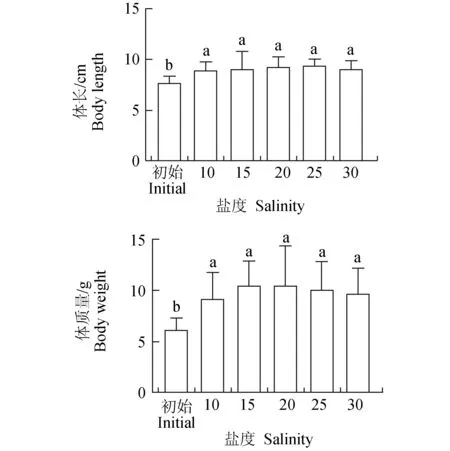
图1 不同盐度下凡纳滨对虾终末体长和体质量的差异
2.2 不同盐度下生物絮团沉降量的变化
盐度越高生物絮团生长越快,30盐度组的生物絮团沉降量在第17 d达到最大值,为200 mL/L,随后逐渐降至43 mL/L,其他各盐度组呈相同变化趋势(图2)。第30 d各盐度絮团沉降量差异不显著(P>0.05)。

图2 不同盐度下生物絮团沉降量的变化
2.3 不同盐度下pH与总碱度的变化
试验各盐度组的pH持续降低,盐度越低,pH降低越快,15盐度组pH最低为7.36,15盐度组在第11 d与20盐度组、25盐度组和30盐度组差异显著(P<0.05),20盐度组与25盐度组、30盐度组差异显著(P<0.05)(图3)。各盐度组的总碱度持续降低,盐度越低,总碱度降低越快,15盐度组总碱度最低,达43 mg/L(以CaCO3计)(图4),10、15盐度组在第11 d与20、25、30盐度组差异显著(P<0.05)。

图3 不同盐度下pH的变化
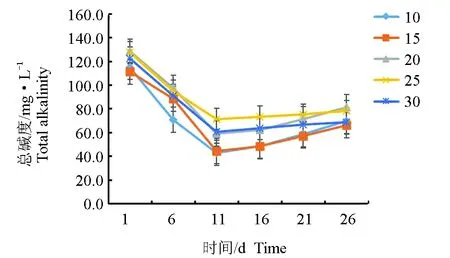
图4 不同盐度下总碱度的变化
2.4 不同盐度下氨氮的变化
氨氮在30盐度组积累的最快,在第6 d达到最大质量浓度8.62 mg/L后,在全部试验周期中持续降低,在第21 d降至0 mg/L,10盐度组积累的最慢,也在第6 d达到最大质量浓度1.71 mg/L,随后在第21 d降至0 mg/L,其他盐度组呈相同趋势变化(图5)。在第6 d各盐度组之间氨氮质量浓度差异不显著(P>0.05)。
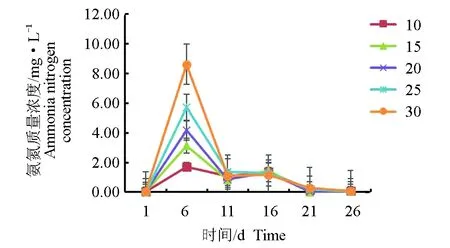
图5 不同盐度下氨氮质量浓度的变化
2.5 不同盐度下亚硝态氮的变化
亚硝态氮在10盐度组积累的最快,在第6 d达到最大质量浓度9.18 mg/L,随后在全部试验周期中持续降低,在第16 d降至0 mg/L,30盐度组积累的最慢,也在第6 d达到最大质量浓度5.79 mg/L,随后在第16 d降至0 mg/L,其他各盐度组呈相同变化趋势(图6)。30盐度组的亚硝态氮浓度质量在第6 d显著高于10、15、20盐度组,25盐度组亚硝态氮质量浓度显著高于10盐度组和15盐度组,10、15、20盐度组之间差异不显著(P>0.05)。
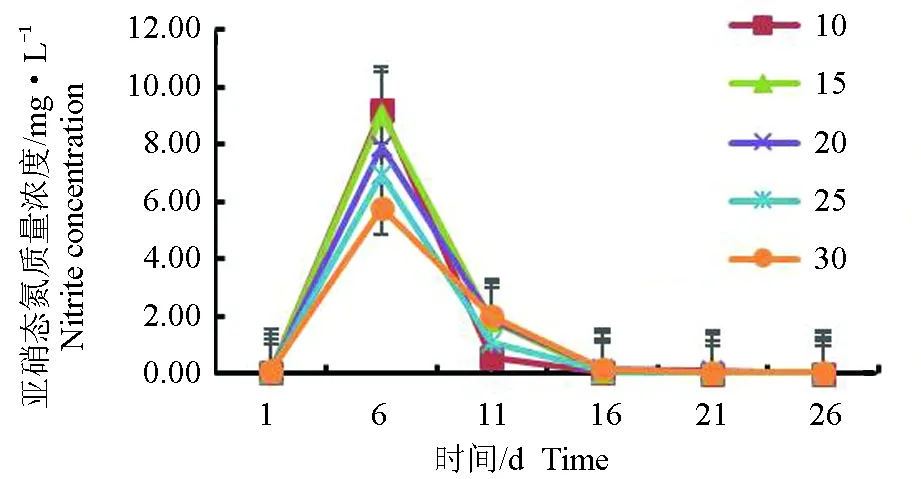
图6 不同盐度下亚硝态氮质量浓度的变化
2.6 不同盐度下硝态氮的变化
试验由第1 d开始到第16 d硝态氮在不同盐度中呈上升的趋势,试验前6 d硝态氮的累积为盐度越低硝态氮质量浓度越高,10盐度组在第6 d硝态氮质量浓度达到最大值,为15.75 mg/L,第16 d盐度越高硝态氮质量浓度越高,30盐度组在第30 d硝态氮质量浓度达到最大值,为58.39 mg/L,随后开始降低,最低达28.63 mg/L(图7)。

图7 不同盐度下硝态氮质量浓度的变化
2.7 盐度对凡纳滨对虾消化指标的影响
低盐度与高盐度肝胰脏脂肪酶活性较高,25、30盐度组的脂肪酶活性最高,达1.21 U/mg,显著高于15、20盐度组的活性,10盐度组脂肪酶活性也高于15、20盐度组的活性,但差异不显著(P>0.05)(图8)。15盐度组的淀粉酶活性显著高于其他盐度组(P<0.05),其他各盐度组间差异不显著(P>0.05)(图9)。
2.8 盐度对凡纳滨对虾超氧化物歧化酶的影响
20、25盐度组肌肉中的超氧化物歧化酶活性显著高于其他盐度组(P<0.05),20盐度组的酶活性最高,此外10盐度组的酶活性也较高,30盐度组的酶活性最低(图10)。肝胰脏中各盐度组酶活性差异不显著(P>0.05),肝胰脏与肌肉中超氧化物歧化酶活性在不同盐度下的规律相似,即先降后升再降。
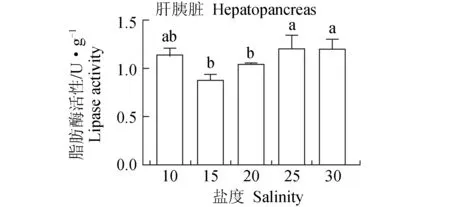
图8 盐度对凡纳滨对虾脂肪酶活性的影响
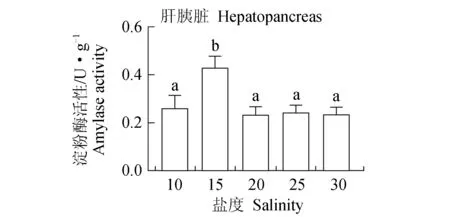
图9 盐度对凡纳滨对虾淀粉酶活性的影响

图10 盐度对凡纳滨对虾超氧化物歧化酶活性的影响
2.9 盐度对凡纳滨对虾过氧化氢酶的影响
30盐度组肌肉中的过氧化氢酶活性显著高于其他各盐度组,其他各盐度组差异不显著(P>0.05)(图11)。肝胰脏与肌肉中过氧化氢酶活性的规律接近,30盐度组的活性也显著高于其他各盐度组,25盐度组的活性低于其他各盐度组。相同盐度下,肌肉中过氧化氢酶的活性低于在肝胰脏中的活性。
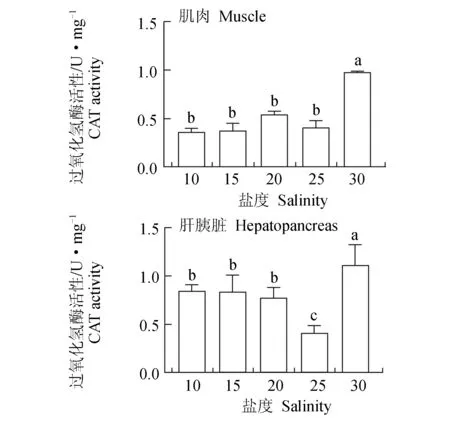
图11 盐度对凡纳滨对虾过氧化氢酶活性的影响
2.10 盐度对凡纳滨对虾碱性磷酸酶的影响
碱性磷酸酶在肌肉中的活性极低,20盐度组的活性高于其他各盐度组,但各组间差异不显著(P>0.05),碱性磷酸酶活性在不同盐度下的规律先升后降(图12)。肝胰脏中,碱性磷酸酶的活性在20盐度组中最高,20、30盐度组活性差异显著(P<0.05),20盐度组与其他各组盐度差异不显著(P>0.05)。肝胰脏中碱性磷酸酶活性在不同盐度下的规律为先降后升再降。
2.11 盐度对凡纳滨对虾酸性磷酸酶的影响
酸性磷酸酶在肌肉中的活性很低,20盐度组的活性高于其他各盐度组且差异显著(P<0.05),30盐度组的活性显著低于其他各盐度组(P<0.05),酸性磷酸酶活性在不同盐度下的规律为先升后降。肝胰脏中,酸性磷酸酶的活性在20盐度组中最高,20盐度组与其他各盐度组活性差异显著(P<0.05),10盐度组显著低于其他各盐度组(P<0.05)(图13)。肝胰脏碱性磷酸酶活性与肌肉的规律相同,即随着盐度升高酶活性先升再降。

图12 盐度对凡纳滨对虾碱性磷酸酶活性的影响
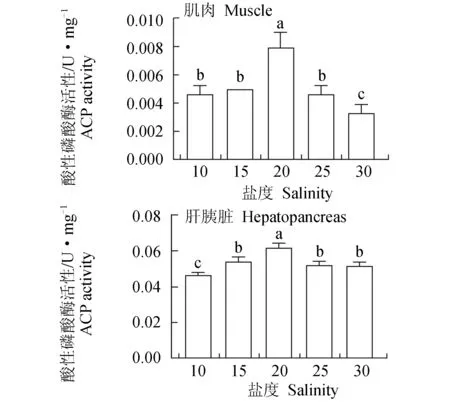
图13 盐度对凡纳滨对虾酸性磷酸酶活性的影响
3 讨 论
3.1 不同盐度下凡纳滨对虾生长指标的差异
盐度是海洋无脊椎动物生存、生长发育的重要影响因子, 盐度变化直接影响动物体内渗透压和离子浓度的调节[14]。已有的研究表明,凡纳滨对虾等广盐性虾类可通过自身的渗透压离子调节来适应水体的不同盐度,水体盐度较高时,虾类排除体内的多余盐分,同时增加对水分的吸收;在盐度较低时又从水体中吸收盐分,并排出多余的水分,来实现自身与环境的渗透压平衡。在这个过程中,虾类要消耗大量的能量,同时虾类的生长也会受到影响[15]。而在处于等渗点的环境时消耗能量少,用于生长的能量比最大[16]。凡纳滨对虾最适宜盐度的研究结果不尽相同,不同试验条件下,凡纳滨对虾生长的最适盐度分别为4~8[17]、2~8[18]、20[9-10]和30[19]。本试验中,盐度15和20的条件下,凡纳滨对虾的生长速度最快。可以得出,在生物絮团模式下凡纳滨对虾的最适生长盐度为15~20。
3.2 不同盐度下pH、总碱度和生物絮团结构的变化
本试验的结果显示,生物絮团对盐度有广泛的适应性,盐度越高生物絮团生长越快。试验所用生物絮团来源于盐度约30的养殖池中,在与原环境盐度相同的30盐度组水体中絮团生长速度最快,但是不同盐度下絮团的演替周期和转化氮元素的周期相同,在时间上保持同步。
生物絮团在生长过程中需要消耗水体中的碱度并产生大量的二氧化碳,使水体pH和总碱度降低,与Ebeling等[20]的研究结果相似。已有研究表明,在生物絮团养殖系统中,碱度降至低于100 mg/L、pH降至低于7.0时会对对虾的健康产生影响[21]。同时,较高的碱度可对水产养殖絮体生物学特性及氨氮转化产生影响[22]。因此在试验过程中需要给水体中持续添加碳酸氢钠或者氧化钙等来维持pH和总碱度。
3.3 不同盐度下生物絮团养殖水体中氮元素的转化
本试验中,盐度越高氨氮累积越快,盐度越低亚硝态氮累积越快。不同盐度下硝态氮的累积趋势类似, 25、30盐度组的硝态氮质量浓度自21 d起开始呈降低的趋势。
生物絮团养殖水体中碳氮比在15以上时,以异养的方式将水体中的三态氮转化为自身的菌体蛋白;在碳氮比低于5时主要依靠硝化作用清除无机氮[23]。本次试验中的生物絮团通过硝化途径来处理水体中的氮元素。生物絮团中的亚硝化细菌将氨氮转化为亚硝态氮,硝化细菌把亚硝态氮转化为硝态氮,伴随着硝态氮质量浓度的增加。这与Avnimelech[5]的研究结果相似,对于试验后期硝态氮质量浓度的降低,初步分析有两种原因:一是反硝化细菌逐渐繁殖增加,将硝态氮进一步转化掉[24];二是生物絮团在试验后期由自养型微生物群落结构向异养型的结构转化,可直接将氮元素转化为自身的有机氮,不需要通过硝化途径的无机氮循环来处理水体中的氮元素[25]。水体中的细菌在繁殖过程中,不仅可以稳定水质,还可以增加凡纳滨对虾肠道菌群的组成丰度[26]。
3.4 不同盐度对凡纳滨对虾酶活性的影响
盐度是对虾养殖中重要的环境因子之一,盐度会直接影响对虾的生长、存活、渗透压调节、免疫和代谢酶活性等[9,27]。试验中淀粉酶的活性随着盐度的升高呈先升后降的趋势,这与侯文杰等[28]的结果相同。同时黄凯等[9]发现,低盐度组的凡纳滨对虾淀粉酶、蛋白酶活性均高于高盐度组,认为可能是低盐度组维持渗透压平衡需要消耗更多的能量,因此需要更高的淀粉酶活性用于消化食物。脂肪酶在盐度25时活性最高,盐度升高或者降低酶活性均随之降低。凡纳滨对虾在盐度为15~25时,超氧化物歧化酶、碱性磷酸酶、酸性磷酸酶活性均维持在较高水平,与沈丽琼等[29]的研究结果相一致。消化酶与免疫酶活性的强弱决定了凡纳滨对虾对营养物质消化吸收的能力以及机体的免疫能力[30],在盐度15~25间较高的酶活性也表明了凡纳滨对虾的适宜盐度,与朱春华[31]的研究结果相一致。
4 结 论
生物絮团是一种零换水的养殖技术,在解决水产养殖环境污染,减少病害的暴发,阻绝病害的传播等方面具有独特的优势,因此开展生物絮团在水产养殖中的应用研究,具有极大的意义。
本试验的结果表明,在生物絮团模式下,凡纳滨对虾的最适生长盐度为15~20。养殖水体中不同盐度下氨氮和亚硝态氮的累计速度有差异,试验前期盐度越高氨氮累积越快,盐度越低亚硝态氮累积越快,此外生物絮团对盐度有广泛的适应性,不同盐度下絮团的演替周期和转化氮元素的周期相同,在时间上保持同步。值得注意的是,在生物絮团模式下,水体中的氮元素通过硝化途径进行转化,会消耗水体中的碱度并产生大量的二氧化碳,使水体pH和总碱度降低,因此在生物絮团养殖模式下,需要添加一定量的碳酸氢钠、氧化钙等药品来调节水质。
猜你喜欢
工程科学学报(2021年11期)2021-11-17 08:54:28
中南大学学报(自然科学版)(2019年7期)2019-08-13 03:17:42
当代水产(2019年1期)2019-05-16 02:42:00
长江科学院院报(2017年3期)2017-03-11 06:14:38
广东海洋大学学报(2015年4期)2016-01-13 08:39:32
应用海洋学学报(2014年1期)2014-11-22 07:17:44
大连大学学报(2014年3期)2014-09-18 09:16:32
金属矿山(2014年7期)2014-03-20 14:19:58
食品工业科技(2014年7期)2014-03-11 18:15:11
金属世界(2013年6期)2013-02-18 16:30:47
