
饲料脂肪水平对大规格松浦镜鲤越冬前后体质量损失、体成分及脂代谢基因表达的影响
2022-02-11 02:49李晋南张圆圆王连生
中国饲料 2022年1期
范 泽, 李晋南, 张圆圆, 吴 迪, 王连生
(中国水产科学研究院黑龙江水产研究所,黑龙江省水生动物病害与免疫重点实验室,黑龙江哈尔滨150070)
越冬期是鱼类养殖过程中一个特殊的阶段,尤其是在我国“三北”(东北、华北、西北)地区。 在越冬过程中,养殖鱼类通过改变生化、生理和分子活动状态来应对越冬产生的食物匮乏和低温冷应激 (Song 等,2019;Long 等,2013;Shoup 和 Wahl,2008)。 鲤鱼作为池塘养殖的主要品种,其在“三北” 地区的年产量约占池塘养殖总量的50% ~60%(中国渔业统计年鉴,2020),因此深入研究越冬对鲤鱼养殖过程中的影响是实现鲤鱼健康养殖的基础且必要工作。
越冬期间,鱼类基础代谢在水温降低至4 ℃以下时显著下降,摄食完全停止,仅仅依赖于体内储存的营养物质产生能量来维持基本生命活动(Xu等,2020;于淼等,2015;Fullerton 等,2011)。研究表明, 脂肪是鲤鱼长期饥饿过程中主要的供能物质(于淼等,2015)。 养殖实践中往往在鲤鱼越冬前投喂高脂饲料,进而提高体内能量物质的储备确保安全越冬,但同时也造成了越冬期结束后鲤鱼较高的死亡率及较弱的体质(王钊,2017;NRC,2011)。 饲料脂肪的适宜添加水平与饲料营养配比、 生长阶段、养殖环境等多种因素相关,目前,鲤鱼越冬前投喂何种水平的脂肪饲料鲜见研究报道。
松浦镜鲤于2008 年通过全国水产原种和良种审定委员会审定,在十余年的推广过程中,养殖规模和分布地域逐年扩大, 取得了良好的经济效益和社会效益,已成为我国鲤主要的养殖品种(石连玉等,2016)。在课题组先前的研究发现,大规格松浦镜鲤[(247.00±16.67)g]对饲料脂肪最适添加水平为6%。 本试验在前期研究基础上探讨饲料脂肪水平对大规格松浦镜鲤越冬前后生长、 体成分及脂代谢基因表达的影响, 探究饲料脂肪水平与鲤鱼越冬的关系, 为鲤鱼在北方越冬期及越冬后的健康养殖提供科学依据, 为开发利于鲤鱼越冬的饲料奠定实践基础。
1 材料与方法
1.1 试验饲料 以鱼粉、豆粕为蛋白质源,豆油为脂肪源, 通过设置不同水平的豆油及次粉配制3%、6%、9%、12%、15%和 18% 6 组脂肪水平的试验饲料,其组成及营养水平见表1。 饲料制作在哈尔滨市合禾饲料有限责任公司完成, 首先将所有干性原料经粉碎机后,过60 目筛,按逐级放大原则使用混匀机搅拌混匀, 后依据饲料配方比例加入豆油, 在混匀机中充分混合后使用饲料制粒机制成直径为3.00 mm 的沉性颗粒饲料。
1.2 试验鱼及饲养管理 在进行完大规格松浦镜鲤对饲料脂肪适宜需要量的研究试验后(2019年10 月底),在每一个网箱中保留规格齐整、质量为(609.70±49.93)g 的 20 尾大规格鲤鱼进行越冬试验,共 18 个网箱(2.0 m×2.0 m×2.0 m),越冬期间不投喂任何食物,只进行水温检测。越冬试验时间为 2019 年 10 月 31 日至 2020 年 4 月 28 日。越冬期间水温3 ~ 8 ℃。
1.3 样本采集 越冬周期结束后对试验鱼进行计数并对所有试验鱼进行称重, 以计算存活率及增重率;从每箱中随机取3 尾鱼,对其体长进行测量,以计算肥满度;试验鱼于冰盘中迅速解剖取其肝胰脏及肌肉, 样本转入-20 ℃超低温冰箱中保存备用。
1.4 指标测定及方法
1.4.1 生长指标计算
存活率/%=Nt/N0×100;
增重率/%=(mt-m0)/m0×100;
肥满度/(g/cm3)=mt/Lt3;
肝体比/%=mh/mt×100;
式中:t 为试验天数,d;Nt为终末存活尾数;N0为初始放养尾数;m0为平均初始体质量,g;mt为平均终末体质量,g;Lt为平均终末鱼体长,cm;mh为肝胰脏质量,g。
1.4.2 肌肉成分的测定 粗脂肪含量采用索氏抽提法(GB/T6433-1994)进行测定,水分及灰分分别采用常压恒温烘干法 (GB6435 -1986),550 ℃灼烧法进行测定(GB/T6438-2007)。
1.4.3 脂代谢相关基因的相对表达量测定 越冬周期结束后各组试验鱼饥饿24 h 后分别投喂各组饲料,在2 h 后每组随机取3 尾鱼,取其肝胰脏于液氮中研磨。利用Trizol 法提取各组织总RNA,参照TaKaRa PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time) (Code No:RR047A)说明书,将总RNA 反转录为cDNA。
根据Genbank 中现有的鲤鱼脂肪酸合成酶(FAS2,HM802556.1)、 脂 蛋 白 脂 酶 (LPL2,FJ436077)、 肉毒碱棕榈酰转移酶-1 (CPT-1,JF728839)、 固 醇 调节 元件 结 合蛋 白 (SREBP,KJ162572)、乙酰辅酶 A 羧化酶(ACC,GU908475)及β-actin 保守序列设计引物(表2)。 所有引物均由生工生物工程(上海)有限股份公司合成。

表2 试验所用引物
实时荧光定量PCR 反应根据SYBRRPremix Ex Taq TM (Tli RNaseH Plus)(Code:RR420A)试剂盒说明书进行, 采用SYBR Green 染色法,在Applied Biosystems 7500 Real Time PCR System实时PCR 检测系统下进行。 为了避免扩增批次间存在差异, 目的基因与其对应的内参基因同时扩增,每个模板均做三个重复,得到三个Ct 值,取其平均值作为最终 Ct 值, 后运用比较 Ct 法(△△Ct) 对所得数据进行分析比较进而得到各模板中各基因的相对表达量,计算公式为:相对表达量=2-△△Ct=2-[(Ct 处理-Ct 内参)-(Ct 对照-Ct 内参)]。
1.5 数据统计 所有数据均用Excel 2007 及SPSS 22 进行分析处理。利用Excel 软件制作各指标的标准曲线, 并利用标准曲线计算得出各样品中对应指标的活性含量。 所有数据均以“平均值±标准差”表示,并利用SPSS 19.0 统计软件进行单因素方差分析(One-way ANOVA),若差异达到显著水平(P < 0.05),则进行Duncan‘s 法多重比较。
2 结果
2.1 饲料不同脂肪水平对松浦镜鲤越冬后鱼体质量及存活率的影响 由表3 可以看出, 越冬前的8 周养殖试验中,6%脂肪组的均重 (668.59 g)显著高于 3%脂肪组 (587.69 g)、15%脂肪组(583.92 g)和 18%脂肪组(556.07 g)(P < 0.05),其增重率 (175.53%) 显著高于其余各组 (P <0.05)。 越冬后,各脂肪水平组鲤鱼体质量呈现一定比例下降, 其中6%脂肪组的越冬后体质量(613.77 g)显著高于 3%脂肪组(506.05 g)、12%脂肪组 (525.04 g) 和 18%脂肪组 (494.87 g)(P <0.05),其体质量损失率(8.25%)显著低于3%脂肪组(13.90%)和 18%脂肪组(11.04%)(P < 0.05)。越冬存活率在各组间无显著差异 (P > 0.05),6%脂肪组最高,为100.00%。

表3 饲料不同脂肪水平对松浦镜鲤越冬后体质量及存活率的影响
2.2 饲料不同脂肪水平对松浦镜鲤越冬后形体指标的影响 由表4 可以看出, 各脂肪组的肥满度和肝体比均表现出一定比例的下降。越冬前后,各组间肥满度均无显著差异(P > 0.05),肥满度下降比以6%脂肪组(7.74%)为最低,以18%脂肪组(16.95%) 为最高。 越冬前,18%脂肪组的肝体比(2.44%) 显著高于除15%脂肪组外的其余各组(P < 0.05), 而越冬后,18%脂肪组的肝体比(2.44%) 显著高于除9%和15%脂肪组外的其余各组(P < 0.05),各组间肝体比下降比无显著差异(P > 0.05),以 6%脂肪组(19.69%)为最低。
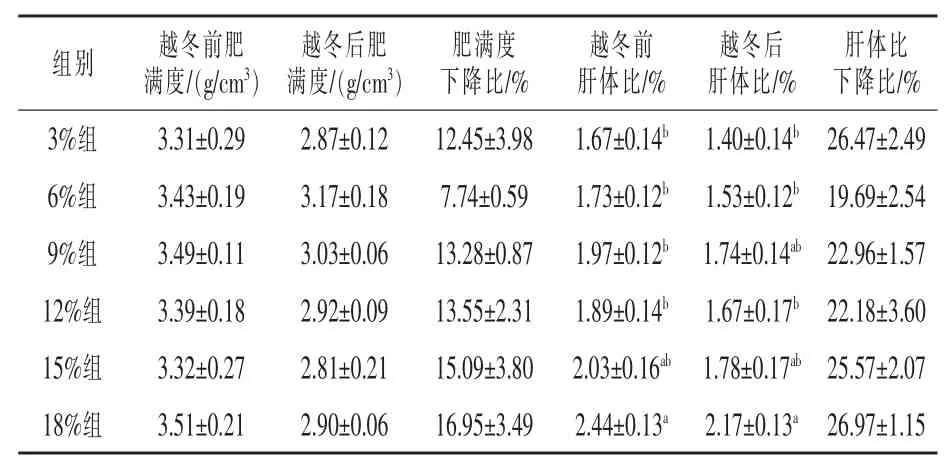
表4 饲料不同脂肪水平对松浦镜鲤越冬前后形体指标的影响
2.3 饲料不同脂肪水平对松浦镜鲤越冬后肌肉和肝胰脏营养成分的影响 由表5 可以看出,各脂肪水平组越冬后肌肉粗蛋白质呈现一定比例的下降, 其中6%脂肪组越冬后肌肉粗蛋白质含量(13.36%)最高,显著高于15%组(12.28%)和18%组(11.28%),且其下降率为最低(13.08%),显著低于 3%组 (32.32%) 和 18%组 (20.58%)(P <0.05)。 各脂肪水平组越冬后肌肉粗脂肪呈现一定比例的下降, 以3%脂肪组的肌肉粗脂肪含量(0.80%)为最低,显著低于其余各组(P < 0.05),而15%组(1.00%)和18%组(1.01%)的肌肉粗脂肪含量显著低于6%、9%和12%三组 (P < 0.05)。6%组(36.06%)和9%组(37.05%)的肌肉粗脂肪下降比显著低于其余各组 (P < 0.05)。 各脂肪水平组越冬后肝胰脏粗脂肪呈现一定比例的下降,越冬后以18%脂肪组的肝胰脏脂肪含量(7.73%)显著高于除15%组外的其余各组 (P < 0.05),其下降比 (30.33%) 亦显著高于其余各组 (P <0.05)。 各脂肪组越冬后肌肉水分呈现一定比例的增加,以3%脂肪组的肌肉水分含量(84.12%)为最高,显著高于 6%组(78.58%)(P < 0.05),其增加比(8.72%)亦显著高于其余各组(P < 0.05),水分增加比最低的为3%脂肪组(0.53%),显著低于除9%组外的其余各组(P < 0.05)。
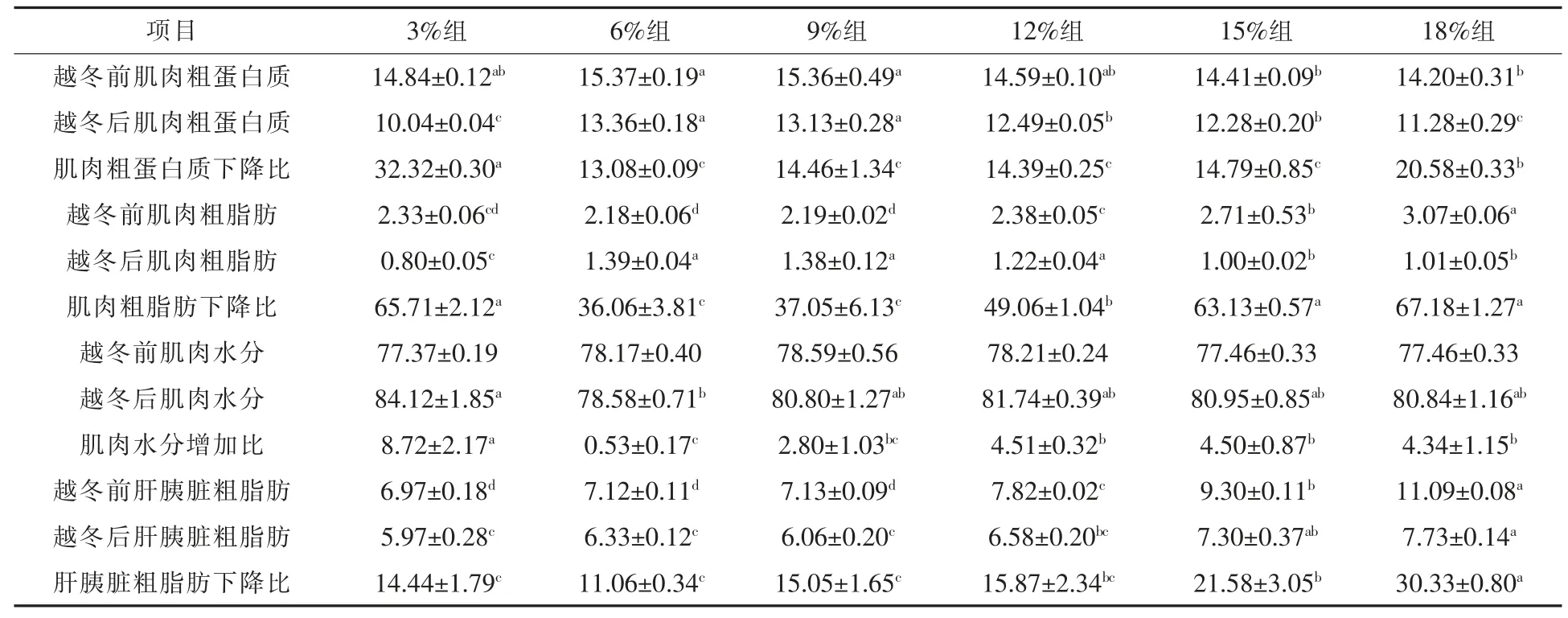
表5 饲料不同脂肪水平对松浦镜鲤越冬前后肌肉和肝胰脏营养成分的影响 %
2.4 饲料不同脂肪水平对松浦镜鲤越冬后脂代谢相关基因表达的影响 由表6 可以看出, 脂肪合成相关的FAS、ACC1 及SREBP 的基因表达量均以6%组为最高, 其中FAS 表达量显著高于其余各组(P < 0.05),ACC1 的基因表达量显著高于3%、15%及 18%组(P < 0.05),SREBP 的基因表达量与其他各组无显著差异(P > 0.05)。脂肪分解相关的CPT1 的基因表达量3%组和6%组显著低于其余各组(P < 0.05),而LPL 基因表达量在各组间无差异(P > 0.05)。
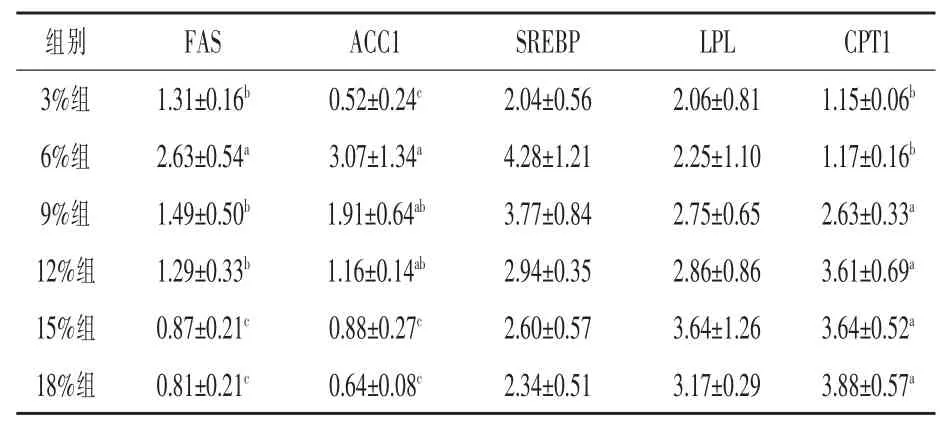
表6 饲料不同脂肪水平对松浦镜鲤越冬后脂代谢相关基因表达的影响
3 讨论
在德国镜鲤选育系基础上进一步培育出的松浦镜鲤,在鳞被、生长及抗逆抗病方面都有着较为显著的提高(于淼,2015)。 于淼等(2015)从形态学、 组织学及生化组成等三方面综合分析了松浦镜鲤在越冬期间发生的变化。 本试验从体质量损失、形态学指标、肌肉及肝胰脏营养成分组成及脂代谢相关基因表达初步探讨越冬前饲料脂肪供给水平对鲤鱼越冬的潜在影响。
本试验结果表明, 越冬前饲料脂肪水平为6%时可有效降低大规格松浦镜鲤 [(609.70±49.93)g]越冬后体质量损失率,而在前期的试验中发现, 同规格松浦镜鲤的适宜脂肪需求量也为6%。 武文一(2020)研究发现,越冬前饲料强化适宜蛋白(31%)和适宜脂肪(4%)水平饲料可有效降低草鱼越冬后体重损失率, 而当饲料脂肪水平为8%时反而会提高体重损失率。 说明无论是养殖阶段还是越冬期过高的脂肪摄入均会带来潜在的脂质过氧化应激,进而提升体质量损失率。养殖实践中肥满度、 肝体比常作为评估鱼体生理生化或营养状况的参考指数 (贾明亮,2010;Froese,2006)。 于淼 (2015) 发现, 越冬后松浦镜鲤[(248.87±15.75)g]的肥满度降低比达到 23.26%,肝体比下降达到29.59%。 而在本试验中,越冬后各脂肪组的肥满度下降比为7.74% ~ 16.95%,肝体比下降比为19.69% ~ 26.07%,均以6%脂肪组为最低,因此从生物学性状来看,越冬前饲料脂肪水平为6%可使松浦镜鲤能量沉积状况处于较好状态(黄春红等,2015)。 这种差异可能主要源于:(1) 本研究中松浦镜鲤越冬前体质量较于淼(2016)试验中的松浦镜鲤重400 余克,规格差异影响了二者越冬后的形态学指标, 即鱼规格大且肥满度高,则越冬效果好(Fullerton 等,2011)。 (2)越冬前饲料脂肪供给会影响鱼类越冬后形态学指标。武文一(2020)发现相较于4%脂肪组,8%脂肪组草鱼会获得更低的肥满度和肝体比。 结合本试验的结果来看, 过高或过低的脂肪水平都会影响机体形态学指标, 进而反映鱼体生理生化或营养状况的异常。
鱼类在越冬期间面对的是低温冷应激 (4 ℃以下)和食物匮乏的双重压力,仅仅依赖于体内储存的营养物质维持基本生命活动。 因此鱼体营养成分的变化能够较好地反映出内源营养在越冬期的利用规律(Li 等,2018)。 于淼(2015)研究脂肪的消耗贯穿整个越冬饥饿期, 蛋白质在越冬饥饿后期才进行消耗, 且越冬后肌肉蛋白质含量与体质量损失率成显著负相关, 说明越冬后肌肉蛋白质水平能体现越冬期体重损失程度, 这种营养物质利用的顺序规律与草鱼相似 (Gong 等,2017)。本试验同样发现, 各脂肪组肌肉及肝胰脏粗脂肪含量、肌肉蛋白质含量均出现一定比例的下降,下降比分别为36.06% ~ 67.18%、11.06% ~ 30.33%及13.08% ~ 32.32%, 均以6%脂肪组为最低,肌肉及肝胰脏粗脂肪下降比均以18%脂肪组为最高, 肌肉粗蛋白质含量下降比以3%脂肪组为最高, 这表明越冬前不适宜的饲料脂肪供给非但不能降低营养物质的消耗和体重损失率的下降,反而可能会促进更多的营养物质的消耗和动员,进而影响鱼体对越冬环境的营养物质消耗策略 (石桂城等,2012)。6%脂肪组在越冬后保持了较高的肌肉蛋白质和粗脂肪含量, 消耗了更少的能量物质,更利于维持体质,便于越冬后机体代谢能力的恢复(武文一,2020)。 与于淼(2015)对松浦镜鲤[(248.87±15.75)g]的研究结果相比,本试验中各脂肪组松浦镜鲤的肌肉粗蛋白质含量下降比均较高, 这表明规格差异可能是影响越冬过程中松浦镜鲤营养物质利用的关键因素。此外,越冬期间各试验组松浦镜鲤肌肉水分含量明显增加, 原因可能是肌肉中蛋白质和脂肪分解代谢后释放出的空间会被体液所填充, 致使肌肉水分含量升高(Wu等,2020;Kojima 等,2015)。
鱼类生长是机体合成代谢不断积累的结果,低温冷应激除了通过限制鱼体摄食和消化吸收功能, 减少合成代谢所需的营养原料而降低鱼类的生长速度外(鲁栋梁,2019),还会影响机体内源营养物质的合成能力(辛芳等,2016)。鲁栋梁(2019)研究发现, 脂肪分解代谢激活剂Fenofibrate 的使用促进了斑马鱼脂肪分解代谢的效率, 提高了斑马鱼的短期冷应激耐受;与之相反,脂肪分解代谢抑制剂Mildronate 的使用抑制了斑马鱼脂肪分解的效率并降低了斑马鱼的短期冷应激耐受能力,表明脂肪的分解利用是鱼类应对短期冷应激的主要供能途径。对草鱼的研究发现,越冬后脂肪合成基因表达受到影响,如脂肪酸合成酶(FAS),乙酰辅酶A 羧化酶 (ACC), 硬脂酰辅酶 A 去饱和酶(SCD),长链酰基 CoA 合成酶(ACSL);同时脂质分解基因表达水平显著上调, 如激素敏感脂酶(HSL),乙酰 CoA 乙酰基转移酶(ACAT)等。 表明草鱼在越冬期间严格限制脂质合成同时加速脂质分解供能 (武文一,2020;Zaarur 等,2019;Mar-queze 等,2018)。 在本试验中,越冬后 6%脂肪组松浦镜鲤的FAS、ACC1 和SREBP 基因表达量更高,CPT1 和LPL 基因表达量更低,鱼体的脂肪动员模式较为合理, 表明越冬前适宜的饲料脂肪供给能够显著增强机体脂肪合成能力, 有助于机体在越冬期间自身内源脂肪的合成, 降低蛋白质用于供能的比例, 这与肌肉粗蛋白质及粗脂肪含量下降结果相吻合。此外,与鲁栋梁(2019)的短期低温冷应激试验不同,越冬后出池水温逐渐上升,鲤能摄取底泥中残留的营养物质, 机体所需脂肪合成的营养原料逐步充足(于淼,2015),进而增强脂肪合成能力。 但饲料脂肪调控越冬期松浦镜鲤的肝胰脏脂代谢的具体作用机制仍需进一步研究。
4 结论
在整个越冬期, 饲料脂肪水平为6%时能够使体质量损失率及肥满度降低比最小化, 在越冬后保持较高的肌肉蛋白质和粗脂肪含量, 消耗更少的能量物质, 使鱼体自身内源脂肪合成能力增强而分解能力减弱,更利于维持体质,便于越冬后饥饿后机体代谢能力的恢复。
猜你喜欢
当代水产(2022年7期)2022-09-20
思维与智慧·下半月(2022年5期)2022-05-17
动漫界·幼教365(中班)(2022年3期)2022-03-11
莫愁·智慧女性(2022年2期)2022-03-01
环球时报(2021-03-09)2021-03-09
河北渔业(2020年6期)2020-06-29
课外生活(小学1-3年级)(2019年12期)2019-09-10
绿色科技(2016年20期)2016-12-27
幸福·婚姻版(2016年5期)2016-05-19
湖北农业科学(2014年21期)2014-12-22
