
苦木碱F通过miR-331-3p调控宫颈癌HeLa细胞增殖及凋亡的机制研究
2022-01-25 07:45邓贵华肖大立
陕西中医 2022年1期
张 琳,邓贵华,杨 亮,肖大立
(1.广东省妇幼保健院药学部,广东 广州 510010;2.广东省妇幼保健院医务科,广东 广州 510010)
宫颈癌是我国中青年女性第二大常见恶性肿瘤,近些年的发病率和病死率均呈显著上升趋势[1-2]。目前,手术及放化疗是宫颈癌的主要治疗方法,但是其有效性却不尽人意。因此,找到更安全、更有效的方法是当前治疗宫颈癌亟待解决的问题。苦木碱F(Picrasidine F)是一种二聚体β-卡巴林生物碱,从苦木(Picrasmaquassioides)的根茎中分离出来。越来越多的研究报道苦木中的生物碱类成分具有抗肿瘤的作用,如苦木生物碱类成分抑制肝癌细胞的增殖[3]。但是苦木生物碱类成分抗肿瘤的分子机制仍有待深入研究。微小核糖核酸(microRNAs,miRNA)是内源性单链小RNA,可通过翻译后修饰在人类肿瘤的发展和进展中发挥重要作用[4-5]。已有研究报道miRNA与宫颈癌细胞的增殖、凋亡、侵袭密切相关,被认为是宫颈癌诊断、治疗和预后评估的新靶点[6-7]。既往研究发现,miR-331-3p对调控宫颈癌细胞增殖有显著影响,可能具有抗癌作用[8]。而且近年来研究报道显示miRNA广泛参与中药的抗肿瘤作用过程中,如青藤碱通过调控miR-324-5p抑制乳腺癌细胞侵袭和迁移[9]。然而,miR-331-3p是否参与苦木碱F对宫颈癌的作用尚不清楚。因此,本研究旨在从miR-331-3p水平深入探讨苦木碱F对宫颈癌细胞增殖和凋亡中的作用及其可能的分子机制,为临床治疗宫颈癌提供新的靶点。

与苦木碱 F 0 μg/ml组比较,*P<0.01;与苦木碱 F 6.25 μg/ml组比较,#P<0.01;与苦木碱 F 12.5 μg/ml组比较,$P<0.01
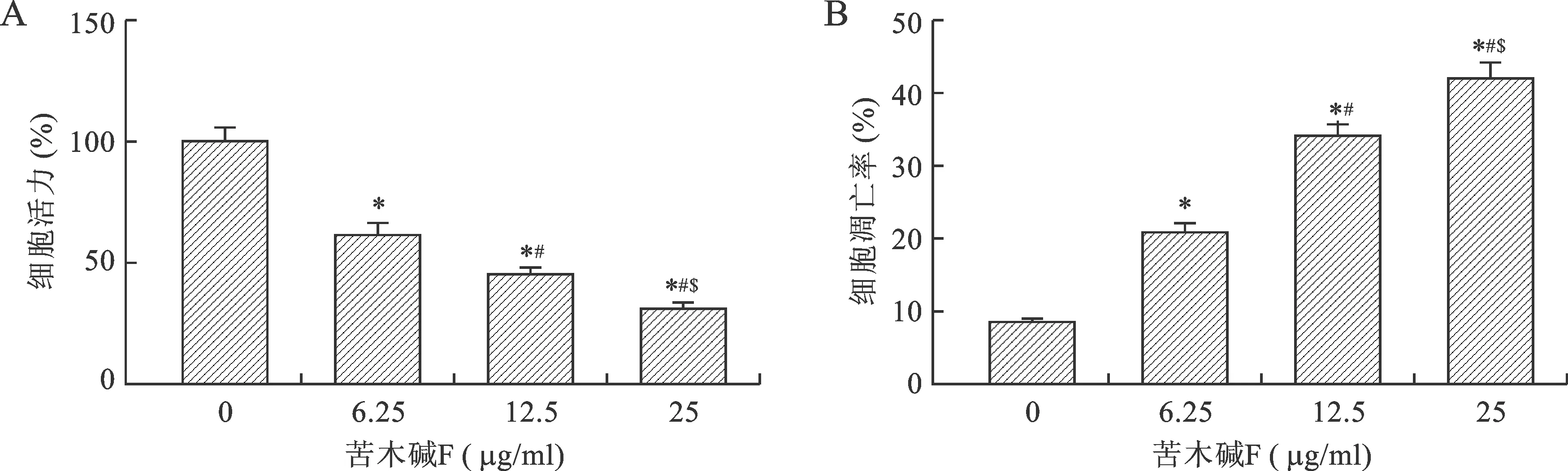
与苦木碱F 0 μg/ml组比较,*P<0.01;与苦木碱F 6.25 μg/ml组比较,#P<0.01;与苦木碱F 12.5 μg/ml组比较,$P<0.01
1 材料与方法
1.1 主要仪器和试剂 主要试剂:人宫颈癌HeLa细胞购自中国科学院上海生物化学与细胞生物学研究所细胞库;RPMI 1640培养基购自美国Gibco公司;MTT细胞检测试剂盒购自上海碧云天生物有限公司;Annexin V-FITC/PI 细胞凋亡检测试剂盒购自北京索莱宝科技有限公司;Trizol试剂、Lipofectamine 2000转染试剂购自Invitrogen公司;PrimeScript RT-PCR试剂盒购自Takara公司;SYBR Green试剂盒购自Takara公司;引物均由Takara公司设计合成;BCA蛋白含量测定试剂盒购自美国Thermo Scientific公司;β-catenin、Bcl-2、Bax、β-actin抗体购自美国Abcam公司;双荧光素酶报告基因载体pGL3以及双荧光素酶报告基因检测试剂盒均购自美国Promega公司;miR-331-3p mimic、NC mimic、miR-331-3p inhibitor、NC inhibitor、蛋白酶体活化复合物3(Proteasome activator complex subunit 3,PSME3)-siRNA、NC-siRNA购自华大基因。
主要仪器:实时荧光定量PCR仪购自德国Eppendorf公司;Thermo Varioskan LUX多功能酶标仪购自美国的Thermo Fisher Scientific公司;CO2培养箱购自美国SHELDON;倒置普通光学显微镜购自日本OLYMPUS公司;BD FACS Calibur流式细胞仪购自美国BD公司;免疫印记系统和凝胶成像分析系统购自美国Bio-Rad公司。
1.2 实验方法
1.2.1 药物配备:本研究根据前人研究方法[10]从苦木Picrasmaquassioides Bennet的根中分离得到β-卡巴林生物碱成分苦木碱F。经1H-NMR和HPLC分析,该化合物的纯度>98%。
1.2.2 HeLa细胞培养:人宫颈癌细胞株HeLa接种于RPMI 1640培养基(含10%胎牛血清、100 IU/ml青霉素和100 μg/ml链霉素)中,置于37 ℃、5% CO2细胞培养箱内进行培养。每3 d进行1/2换液,待细胞融合到70%左右进行转染。选取对数生长期的HeLa细胞接种到96孔板中,使用Lipofectamine 2000TM试剂将miR-331-3p mimic、NC mimic、miR-331-3p inhibitor、NC inhibitor、PSME3-siRNA转染到HeLa细胞中,37 ℃孵育24 h,转染后收集各组细胞用于后续实验。
1.2.3 MTT实验:转染24 h后,将HeLa细胞以每孔1×104个细胞接种于96孔板中。分别于24、48、72 h收取细胞,于每个孔中加入25 μl 1 mg/ml MTT溶液。继续培养4 h后,吸掉培养基上清液,向每个孔中加入100 μl DMSO。10 min后待晶体充分溶解后,用酶标仪测量490 nm处的吸光度。
1.2.4 RT-qPCR:根据Trizol试剂说明书从细胞中提取总RNA,然后使用Prime Script RT试剂盒将RNA反转录为cDNA。当检测miRNA表达时,使用Taq Man microRNA逆转录试剂盒根据其说明书将RNA反转录为cDNA。以cDNA为模板,使用SYBR Green进行PCR扩增。反应条件如下:预变性95 ℃30 s;95 ℃变性5 s;在60 ℃退火30 s,共35个循环。使用2-△△Ct方法计算相对基因表达。采用GAPDH和U6分别作为mRNA和miRNA的内参。基因引物序列如下:PSME3 正向:5’-TCTGACATGAATCTCCCAGTCC-3’,反向:5’-CTCATCCAACCTTCGCTTCTTAT-3’;GAPDH 正向:5’-CAGCAAGAGCACAAGAGGAA-3’,反向:5’-ATGGTACATGACAAGGTGCGG-3’;miR-331-3p 正向:5’-GCGCTAGGTATGGTCCCAG-3’,反向:5’-GTG CAG GGT CCG AGG T-3’;U6 正向:5’-GCT TCG GCA GCA CAT ATACTA AAA T-3’,反向:5’-CGC TTC ACG AAT TTG CGTGTC AT-3’。
1.2.5 流式细胞术:转染到细胞后,各组细胞用不含EDTA的0.25%胰蛋白酶消化,制备单细胞悬液,PBS洗涤3次。将细胞重悬于预冷的1×Binding Buffer中,反复吹打细胞,并用移液器充分混合使细胞密度约为1×106个/ml。加入5 μl Annexin V-FITC,轻轻混合,室温避光反应15 min。细胞重悬于0.5 ml预冷的1×Binding Buffer中。加入碘化丙啶(PI)10 μl,流式细胞仪检测细胞凋亡率。实验重复3次,取平均值。
1.2.6 双荧光素酶报告基因系统:将野生和突变的PSME3的3’UTRs(PSME3 3’UTR-WT或3’UTR-MUT)分别插入pGL3报告基因载体的下游。然后将带有miR-331-3p mimic和PSME3-pGL3报告基因的载体共转染到细胞中,在37 ℃、5% CO2细胞培养箱内培养48 h。然后收集细胞,利用双荧光素酶报告基因进行裂解和分析,将F/R作为各种细胞的相对荧光素酶活性值。
1.2.7 Western blot:收集各组转染48 h的细胞,加入RIPA细胞裂解液,冰上提取细胞总蛋白。然后,采用BCA蛋白检测试剂盒检测蛋白浓度。将蛋白质样品在沸水浴中加热变性。将等量变性后的蛋白样品加到上样孔,进行SDS-PAGE凝胶电泳。蛋白质分离后,转移到PVDF膜上进行电转移。将转化的膜用5%脱脂牛奶封闭液室温封闭1 h。稀释一抗(1∶1000)并在4 ℃下孵育过夜。然后稀释二抗(1∶3000)于室温下孵育30 min,然后用TBST洗涤3次。用化学发光法检测蛋白,采用Image J分析软件计算各组细胞蛋白相对表达量。

2 结 果
2.1 苦木碱F对宫颈癌HeLa细胞增殖和凋亡的影响 通过6.25、12.5、25 μg/ml苦木碱F处理宫颈癌HeLa细胞后,检测苦木碱F对HeLa细胞增殖和凋亡的影响,结果显示与对照组比较,6.25、12.5、25 μg/ml苦木碱F组的细胞活力均显著降低,细胞凋亡率均显著升高,随着浓度的增加,细胞存活率逐渐降低,提示苦木碱F呈浓度依赖性抑制HeLa细胞的存活(P<0.01),见图1。
2.2 苦木碱F上调HeLa细胞miR-331-3p的表达 通过使用6.25、12.5、25 μg/ml苦木碱F处理HeLa细胞24 h后,RT-qPCR实验结果显示,与对照组相比,6.25、12.5、25 μg/ml苦木碱F组中HeLa细胞miR-331-3p水平均显著升高,并且呈浓度依赖性,差异有统计学意义(P<0.01),见图2。结合上述结果得知25 μg/ml苦木碱F更加显著抑制HeLa细胞增殖,促进细胞凋亡,并且上调miR-331-3p的表达,因此进一步选择25 μg/ml苦木碱F用于后续实验。
2.3 苦木碱F通过调控miR-331-3p对HeLa细胞增殖和凋亡的影响 结果显示,与NC mimic组比较,miR-331-3p mimic组中miR-331-3p的水平较显著升高,细胞存活率降低,细胞凋亡率升高,Bcl-2蛋白表达水平降低,Bax蛋白表达水平升高(P<0.05);而miR-331-3p inhibitor+苦木碱F组细胞存活率较NC inhibitor+苦木碱F组显著升高,细胞凋亡率降低,Bcl-2蛋白表达水平升高,Bax蛋白表达水平降低(P<0.05),见图3。以上结果表明过表达miR-331-3p可以抑制HeLa细胞增殖,促进细胞凋亡,而miR-331-3p inhibitor可以逆转苦木碱F对宫颈癌HeLa细胞增殖和凋亡的作用。
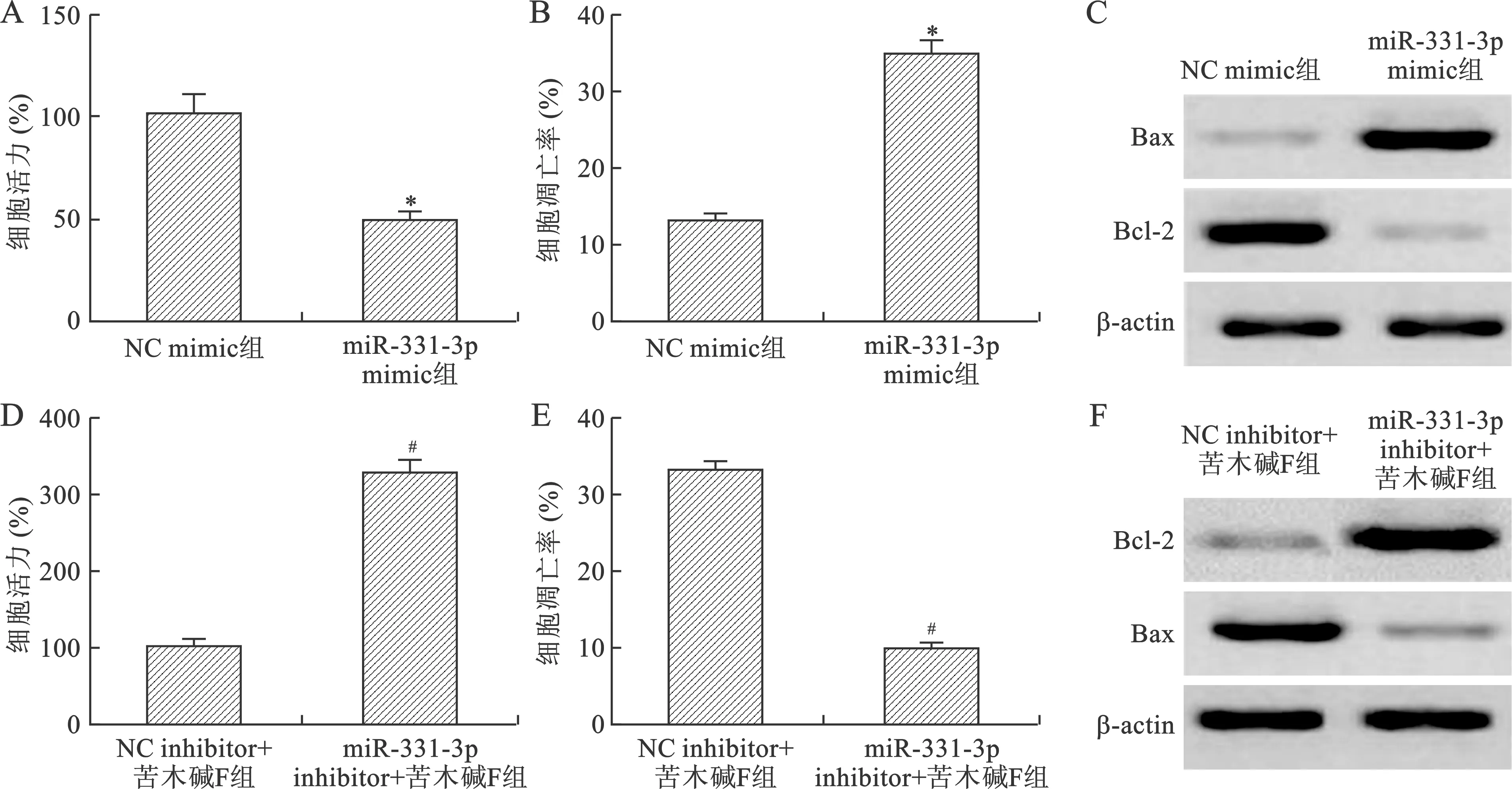
与NC mimic组比较,*P<0.05;与NC inhibitor组比较,#P<0.05
2.4 miR-331-3p靶向PSME3并抑制其表达 通过Targetscan数据库预测miR-331-3p与PSME3-3’UTR的靶向结合位点。进一步通过双荧光素酶报告基因系统验证,结果显示与NC mimic+WT-PSME3-3’UTR组相比,miR-331-3p mimic+WT-PSME3-3’UTR组荧光素酶活性显著降低,差异有统计学意义(P<0.05);与NC mimic+MUT-PSME3-3’UTR组相比,miR-331-3p mimic+MUT-PSME3-3’UTR组荧光素酶活性差异无统计学意义(P>0.05);与NC mimic组比较,miR-331-3p mimic组中PSME3 mRNA水平显著降低;与NC inhibitor组比较,miR-331-3p inhibitor组中PSME3 mRNA水平显著升高(P<0.05),见图4。
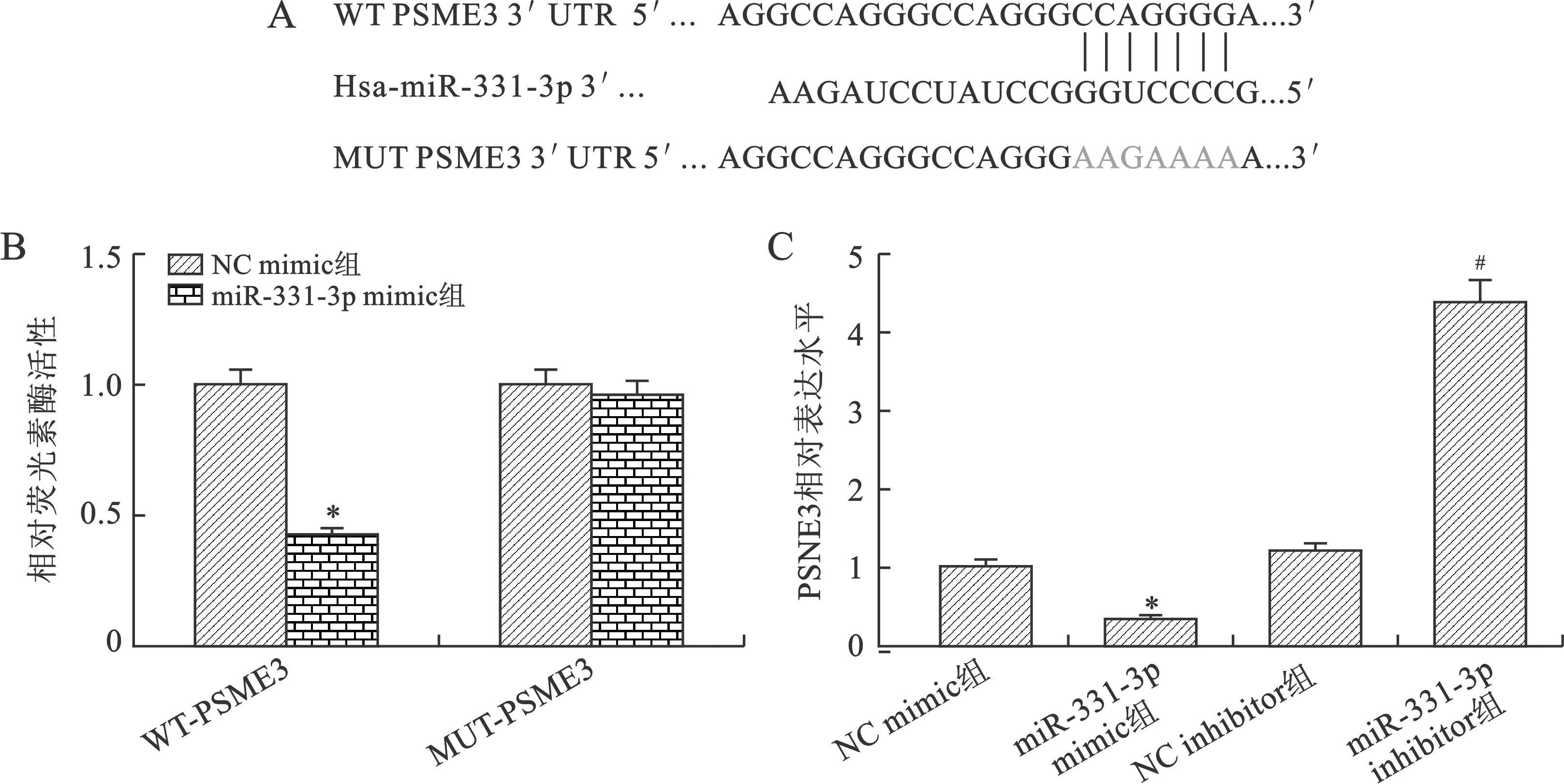
与NC mimic组比较,*P<0.05;与NC inhibitor组比较,#P<0.05
2.5 PSME3在苦木碱F介导miR-331-3p影响HeLa细胞增殖和凋亡中的潜在作用 与苦木碱F+NC inhibitor+NC-siRNA组比较,苦木碱F+miR-331-3p inhibitor+NC-siRNA组中细胞存活率升高,细胞凋亡率降低,β-catenin和Bcl-2蛋白表达水平升高,Bax蛋白表达降低;与苦木碱F+miR-331-3p inhibitor+NC-siRNA组比较,苦木碱F+miR-331-3p inhibitor+PSME3-siRNA组中细胞存活率降低,细胞凋亡率升高,β-catenin和Bcl-2蛋白表达水平降低,Bax蛋白表达水平升高(P<0.05),见图5。
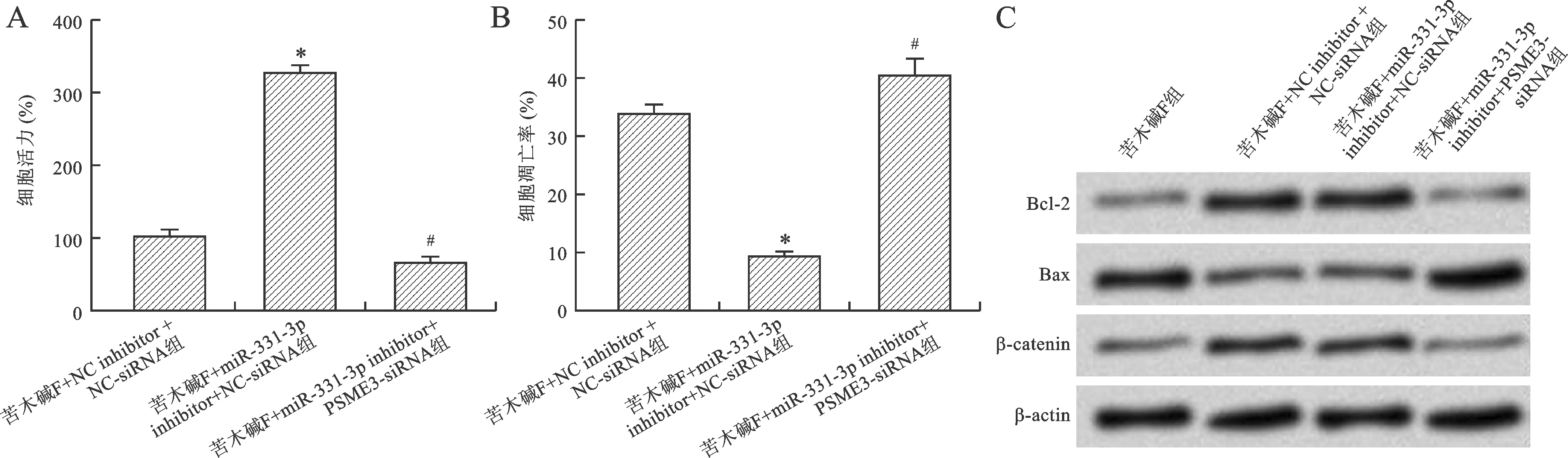
与苦木碱F+NC inhibitor+NC-siRNA组比较,*P<0.05;与苦木碱F+miR-331-3p inhibitor+NC-siRNA组比较,#P<0.05
3 讨 论
宫颈癌是一种常见的妇科恶性肿瘤,是世界各国特别是发展中国家妇女癌症相关死亡的主要原因。因此,需要筛选宫颈癌发生发展的有效治疗药物,探讨其分子机制。本研究探讨苦木碱F对宫颈癌细胞的抗肿瘤作用。我们发现苦木碱F能抑制HeLa细胞的增殖,促进细胞凋亡。在HeLa细胞中,苦木碱F可以上调miR-331-3p的表达水平。此外,在HeLa细胞中,低表达miR-331-3p可逆转苦木碱F对HeLa细胞增殖和凋亡的调控作用。苦木碱F通过miR-331-3p/PSME3调控HeLa细胞的增殖及凋亡。苦木碱F是一种从苦木根茎中分离得到的β-卡巴林生物碱成分。苦木中的生物碱类成分具有一定的抗肿瘤活性。Yamashita等[11]报道一种苦木生物碱成分能显著抑制乳腺癌细胞的增殖。石国华等[12]研究发现3个二聚β-卡巴林生物碱在抑制胃癌MKN-28以及黑色素瘤B-16细胞中起着重要作用。与这些研究结果相似,本实验的结果显示苦木碱F能抑制HeLa细胞增殖,促进细胞凋亡。表明苦木碱F对宫颈癌的治疗可能有一定的抗肿瘤作用。
近年来,越来越多的证据表明miRNAs在宫颈癌中异常表达参与调控肿瘤的生长与转移,如miR-203、miR-41、miR-221[13-15],提示miRNAs可能作为新的预后标志物或宫颈癌患者的前瞻性药物治疗靶点。研究报道,miR-331-3p在多种癌症中具有重要作用[16-17]。在宫颈癌中,有研究报道miR-331-3p抑制细胞增殖,诱导G2/M期阻滞和凋亡[8]。本研究发现苦木碱F使miR-331-3p表达水平升高。miR-331-3p过表达能抑制HeLa细胞增殖,促进凋亡,这与之前的研究结果一致。miR-331-3p抑制剂可逆转苦木碱F对HeLa细胞增殖抑制和凋亡促进作用。这些结果表明苦木碱F抗肿瘤作用可能是通过上调miR-331-3p介导的。国内外大量研究表明,miRNA在转录后通过负向调控靶基因的稳定性或翻译效率来调控基因表达,从而在生物体中发挥致癌或抑癌作用[18-19]。研究[20]发现miR-331-3p通过靶向人表皮生长因子受体2参与结直肠癌细胞生长调控。研究报道miR-331-3p通过靶向髓系/淋巴/混合系白血病易位基因10在肺癌细胞增殖、上皮-间充质表型转换介导的转移和侵袭中发挥抑制作用[21]。本实验的研究结果显示PSME3是miR-331-3p的下游靶基因,miR-331-3p过表达可抑制PSME3的表达,表明PSME3可能参与宫颈癌的发生和发展。
PSME3是一种蛋白酶体成分,大量研究强调了PSME3在癌变中的重要性。在乳腺癌中,沉默PSME3会抑制细胞活力诱导细胞周期阻滞[22]。在子宫内膜癌中,PSME3敲低可抑制细胞增殖、迁移和侵袭能力[23]。与上述结果一致的是,本实验发现苦木碱F处理抑制宫颈癌细胞增殖,转染miR-331-3p抑制剂削弱了这一作用,而敲除PSME3再一次逆转了miR-331-3p抑制剂对HeLa细胞的作用,表明苦木碱F通过miR-331-3p/PSME3抑制HeLa细胞的增殖能力,促进细胞凋亡。Wnt/β-catenin通路调控细胞过程,如增殖,侵袭和分化等。已有研究表明,激活Wnt/β-catenin通路可促进宫颈癌细胞增殖,抑制细胞凋亡[24]。因此,通过分析Wnt/β-catenin信号通路,探讨miR-331-3p抑制剂对苦木碱F诱导HeLa细胞的作用机制。结果显示,miR-331-3p抑制剂能削弱苦木碱F对Wnt/β-catenin信号通路的抑制作用,而沉默PSME3逆转了这一结果。表明苦木碱F是通过上调HeLa细胞中的miR-331-3p表达来抑制Wnt/β-catenin通路的激活,提示Wnt/β-catenin通路参与宫颈癌细胞中苦木碱F的抗肿瘤活性。
综上所述,苦木碱F通过miR-331-3p/PSME3途径抑制HeLa细胞增殖,促进细胞凋亡。本研究表明苦木碱F对宫颈癌具有一定的抗肿瘤作用,为临床治疗宫颈癌提供了新的思路。苦木碱F对宫颈癌的潜在作用仍需进一步研究。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
西安交通大学学报(医学版)(2021年3期)2021-05-15
中日友好医院学报(2021年1期)2021-04-14
昆明医科大学学报(2021年2期)2021-03-29
广东蚕业(2021年1期)2021-03-18
临床超声医学杂志(2020年12期)2020-12-20
山东医药(2020年9期)2020-05-20
数码世界(2018年1期)2018-12-23
热带农业科学(2015年9期)2015-10-14
中国民族民间医药·下半月(2014年4期)2014-09-26
