
塔克拉玛干沙漠微生物研究进展
2021-11-20 05:55刘光琇张昺林章高森
微生物学杂志 2021年5期
李 娟,刘光琇,张昺林,张 威,陈 拓,章高森
(1.中国科学院 西北生态环境资源研究院,甘肃 兰州 730000;2.中国科学院大学,北京 100049;3.甘肃省极端环境微生物资源与工程重点实验室,甘肃 兰州 730000)
塔克拉玛干沙漠是中国最大的沙漠,也是世界第二大流动沙漠,由北边的天山和南边的昆仑山和阿尔金山环绕而成[1-2]。面积约为337 000 km2,85%的沙子是流沙,气候温暖、干燥并且多风。年平均温度9.9~12.6 ℃,年平均降雨量17.4~66.3 mm,年平均蒸发量2 500~3 400 mm。塔克拉玛干沙漠沙丘具有多种类型,并且移动性较强[3-4]。与其他沙漠相比,塔克拉玛干沙漠腹地的沙子类型主要以细沙和极细沙为主,粒径0.063~0.250 mm,不同沙丘类型的沙子粒径也有差异[5]。沙漠中有两种风向,沙漠东部主要受东北风影响,西部主要受西北风影响[6]。由于环境极端严酷,塔克拉玛干沙漠中能够生存的动植物极少,尤其在沙漠腹地。沙漠周边能存活的植被主要为胡杨、柽柳、芦苇、骆驼刺、沙拐枣、罗布麻和管花肉苁蓉等极端耐旱的植物[7-8];已发现的动物有野生双峰驼、沙狐、赤狐、草原斑猫、塔里木马鹿和野猪等[9]。因此,微生物是塔克拉玛干沙漠生态系统的主要组成部分。近年来已有不少关于塔克拉玛干沙漠微生物的研究。本文从塔克拉玛干沙漠微生物的群落组成、新种资源和菌株应用方面进行综述,并对塔克拉玛干沙漠微生物的深入研究提出展望,以期为后续的研究提供参考。
1 塔克拉玛干沙漠不同区域微生物群落特征
1.1 塔克拉玛干沙漠腹地的微生物群落组成
Yu等[10]从塔克拉玛干沙漠的表层样品中,使用T2M和PTYG两种培养基分离沙漠菌株,结果表明:T2M培养基分离到8个属,分别是节杆菌属(Arthrobacter)、异常球菌属(Deinococcus)、微枝形杆菌属(Microvirga)、类诺卡氏菌属(Nocardioides)、庞氏杆菌属(Pontibacter)、副球菌属(Paracoccus)、考克氏菌属(Kocuria)和Rufibacter。其中庞氏杆菌属为优势属。PTYG培养基上分离到6个属,分别是土壤球菌属(Agrococcus)、纤维菌属(Cellulomonas)、诺尔士菌属(Knoellia)、溶杆菌属(Lysobacter)、微枝形杆菌属和动性微菌属(Planomicrobium)。其中优势属为土壤球菌属和诺尔士菌属[10]。另外,龚春霞[11]、卡丽比努尔·艾依提[12]还从塔克拉玛干沙漠中分离到一些藻类,主要是绿藻和蓝藻,绿藻有Tetranephris、衣藻(Chlamydomonas)、咸胞藻(Brachiomonas)、拟绿囊藻(Chlorosarcinopsis)、月牙藻(Pseudokirchneriella)和小球藻(Chlorella),蓝藻有束丝藻(Aphanizomenon)、念珠藻(Nostoc)和细鞘丝藻(Leptolyngbya)[11-12]。蓝藻可进行光合作用,这对于寡营养环境尤其重要,因为光合作用参与了关键的地球化学循环,如碳源和氮源的利用以及胁迫反应[13-15]。
石下生微生物在塔克拉玛干沙漠腹地也常有发现,这些微生物可以通过岩石的遮蔽降低对热的应激且防止紫外线辐射[16]。石下生微生物能够定殖的岩石大多是透明或半透明的,这为光能自养的石下微生物提供了能源[17]。末端限制性片段长度多态性(T-RFLP)、变性梯度凝胶电泳(DGGE)指纹图谱和16S rRNA基因测序[18-20]等研究方法的结果表明,塔克拉玛干沙漠的石下生微生物中的生产者主要是蓝藻菌门,其中拟甲色球藻(Chroococcidiopsis)最常见。异养细菌优势菌来自于酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)和异常球菌-栖热菌门(Deinococcus-Thermus)[20]。Pointing等[18]通过用培养基分离菌株发现了异常球菌属。其他沙漠石下生微生物的报道已有很多,如莫哈维沙漠[21]、内盖夫沙漠[22]和纳米布沙漠[23]。研究表明,石下生微生物的多样性与液体可利用水成正相关,而与温度和降雨量无关[18]。
An等[19]将采集于塔克拉玛干沙漠沙丘顶部的样品,使用16S rRNA基因测序方法,对微生物群落组成进行探究。测序结果表明,共有413个属,15个门,优势菌门是拟杆菌门、变形菌门、放线菌门和厚壁菌门(Firmicutes)。研究表明沙子土壤的C/N比与细菌群落显著相关,而pH与细菌群落结构没有相关性[19]。
1.2 塔克拉玛干沙漠周边的微生物群落组成
徐玲花[24]从塔克拉玛干沙漠北部的胡杨林土壤分离的菌株属于放线菌门、厚壁菌门、拟杆菌门和变形菌门的11个属,这些菌株具有固氮作用。王子旋等[25]在植物根部(红柳、胡杨、沙棘和骆驼刺)土样中共分离出10种芽胞杆菌属(Bacillus)菌株,分别是萎缩芽胞杆菌(Bacillusatrophaeus)(3.15%)、枯草芽胞杆菌(Bacillussubtilis)(15.74%)、地衣芽胞杆菌(Bacilluslicheniformis)(2.66%)、巨大芽胞杆菌(Bacillusmegaterium)(14.78%)、蜡样芽胞杆菌(Bacillusscereus)(1.69%)、嗜碱芽胞杆菌(Bacillusalcalophilus)(8.28%)、Bacillusfirmus(4.84%)、简单芽胞杆菌(Bacillussimplex)(0.24%)、球形芽胞杆菌(Bacillussphaericus)(0.24%)和短小芽胞杆菌(Bacilluspumilus)(48.43%)。董艳萍等[26]在南缘的植物根际土壤中分离出放线菌24个属,分别为拟诺卡氏菌属(Nocardiopsis)、链霉菌属(Streptomyces)、考克氏菌属、分支杆菌属(Mycobacterium)、类诺卡氏菌属、节杆菌属、土壤球菌属、假诺卡氏菌属(Pseudonocardia)、乔治菌属(Georgenia)、糖单孢菌属(Saccharomonospora)、微球菌属(Micrococcus)、诺卡氏属(Nocardia)、小球菌属(Microbacterium)、产丝菌属(Myceligenerans)、迪茨氏菌属(Dietzia)、短状杆菌属(Brachybacterium)、原小单胞菌属(Promicromonospora)、红球菌属(Rhodococcus)、棒状杆菌属(Corynebacterium)、Nonomurae、小单胞菌属(Micromonospora)、Jiangella、马杜拉放线菌属(Actinomadura)和疣孢菌属(Verrucosispora),优势属为拟诺卡氏菌属和链霉菌属。在很多沙漠中放线菌门都是优势菌门,比如阿塔卡玛和纳米布沙漠[27-28]。放线菌在干旱环境中作为优势门是不足为奇的,因为它们具有产孢能力、广泛的代谢和降解能力,具有次生代谢物合成和多重紫外修复机制[29-31]。
徐玲花[24]采用构建克隆文库和T-RFLP两种方法对塔克拉玛干沙漠的无植被沙漠土壤以及胡杨林根际土壤中固氮微生物nifH的基因多样性进行测定,表明固氮微生物主要是α-变形菌纲(Alpha-proteobacteria)。沙漠中微生物可以通过基因转移从而进行固氮,比如拟杆菌门的Pontibacter通过从α-变形菌纲的菌株获得nifH基因,从而进行固氮。
任敏[32]在塔克拉玛干沙漠的北缘和南缘共采集了18份样本,采用宏基因测序方法测定微生物的群落结构,结果表明,微生物群落主要以细菌为主,占微生物群落的89.72%,真菌、古菌和病毒分别占0.71%、7.36%和2.21%。优势菌门包括变形菌门(36.50%~37.98%)、放线菌门(30.44%~37.5%)、厚壁菌门(5.53%~6.63%)。另外,拟杆菌门、异常球菌-栖热菌门、螺旋体门(Spriochaeats)和浮霉菌门(Planctomycetes)也是优势菌门。古菌和真菌的优势菌门分别是广古菌门(Euryarchaeota)和子囊菌门(Ascomycota)。优势菌纲是放线菌纲(Actinomycetia)、α-变形菌纲、β-变形菌纲(Beta-proteobacteria)、γ-变形菌纲(Gamma-proteobacteria)、δ-变形菌纲(Deltaproteobacteria)、拟杆菌纲(Bacteroidetes)、嗜盐菌纲(Halobacteria)、芽胞杆菌纲(Bacillibacteria)、梭菌纲(Clostrida)和异常菌纲(Deinococci)。优势属为链霉菌属、棒状杆菌属、分支杆菌属、拟诺卡氏菌属、假单胞菌属(Pseudomonas)、戈登氏菌属(Gordonia)、伯克霍尔德菌属(Burkholderia)、拟无枝酸菌属(Amycolatopsis)、红球菌属和放线菌属(Actinomyces)。
1.3 塔克拉玛干沙漠气溶胶中的微生物种类和数量
亚洲沙尘暴每年能将大约(4~8)×1011kg的沙子带到世界其他地区[33]。沙子携带的微生物不仅能改变大气中微生物组成,而且也可能对环境和人类健康带来危害[34]。已有很多研究报道微生物可以通过空气进行长距离运输[35]。尽管沙子中的细菌仅能被风运输1 000 km,但粉尘中的细菌可以被运输5 000 km[36]。粉尘中的颗粒可以穿透人体肺部,导致肺部与粉尘相关的潜在过敏原和病原体增加[37],所以研究气溶胶中携带的微生物具有重要意义。
由于塔克拉玛干沙漠中85%的沙子为流沙,风力作用很容易使其形成沙尘暴。不论是沙漠腹地还是周边,可培养方法分离塔克拉玛干沙漠气溶胶中的微生物主要为放线菌门、厚壁菌门、拟杆菌门和变形菌门,优势属为链霉菌属、芽胞杆菌属(Bacillus)、节杆菌属、动性球菌属(Planococcus)、副球菌属、假单胞菌属、Massilis、鞘氨醇单胞菌属(Sphingomonas)和地杆菌属(Pedobacter)[38]。真菌属有链格孢属(Alternaria)、枝孢属(Cladosporium)、亚隔孢壳属(Didymella)、埃里格孢属(Embellisia)、镰刀菌属(Fusarium)、赤霉菌属(Gibberella)、Lewia、青霉菌属(Penicillium)、单格孢属(Ulocladium)、Tetracladium、派伦霉属(Peyronellaea)、栓菌属(Trametes)和单端孢霉属(Trichothecium)[38-39]。与沙尘暴前期分离的微生物多样性相比,沙尘暴中期和后期分离到的多样性更高[41-42]。选取气溶胶的位置不同,分离到的微生物种类差异也很大,如从塔克拉玛干沙漠的东北、东南方向和腹地5个沙尘源区分离到18个细菌属[40],而从各个方向的周边及腹地共23个地区,分离到47个细菌属[38]。沙漠周边地区微生物的多样性明显高于腹地[38,41],这可能是沙漠周边植被较腹地丰富的原因。
研究塔克拉玛干沙漠气溶胶中的微生物,不论是Illumina测序还是T-RFLP方法,都表明主要的细菌来自于放线菌门、厚壁菌门、拟杆菌门和变形菌门,这与可培养方法结果一致[41]。其中优势属为微杆菌属(Microbacterium)、不动杆菌属(Acinetobacter)、短波单胞菌属(Brevundimonas)、柄杆菌属(Caulobacter)、叶杆菌属(Phyllobacterium)、假单胞菌属、庞氏杆菌属和根瘤菌属(Rhizobium)。真菌主要归属于子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和壶菌门(Chytridiomycota),优势属为Lactifluus、伊萨酵母属(Issatchenkia)、Microidium和Alternaris[40]。
可培养方法可以分离出很多种类的微生物菌株,然而用这种分离方法存在很多的局限性,比如培养基、生长温度、pH、接种量、培养时间等的不同都会造成微生物个数与种类的差异[42]。此外可培养方法分离到的微生物菌株数量有限,已有实验数据表明,99%的微生物菌株都是难培养的[43]。与可培养方法相比,不可培养方法会获得更多的微生物种类信息,比如宏基因测序方法,不仅能检测全部微生物的群落组成,还能检测微生物的群落功能[44]。不可培养方法不能获得菌株,因此可培养方法是获得新的微生物菌种资源的一种有效办法。
2 塔克拉玛干沙漠发现的微生物新种
由于塔克拉玛干沙漠的环境特殊性,在沙漠中发现了大量微生物新种[45],鉴定并正式命名的新种模式菌株已有17个,其中3株归属于拟杆菌门,2株归属于变形菌门,8株归属于放线菌门,2株归属于异常球菌-栖热菌门,1株归属于厚壁菌门,另外1株归属于真核生物的子囊菌门[46](表1)。
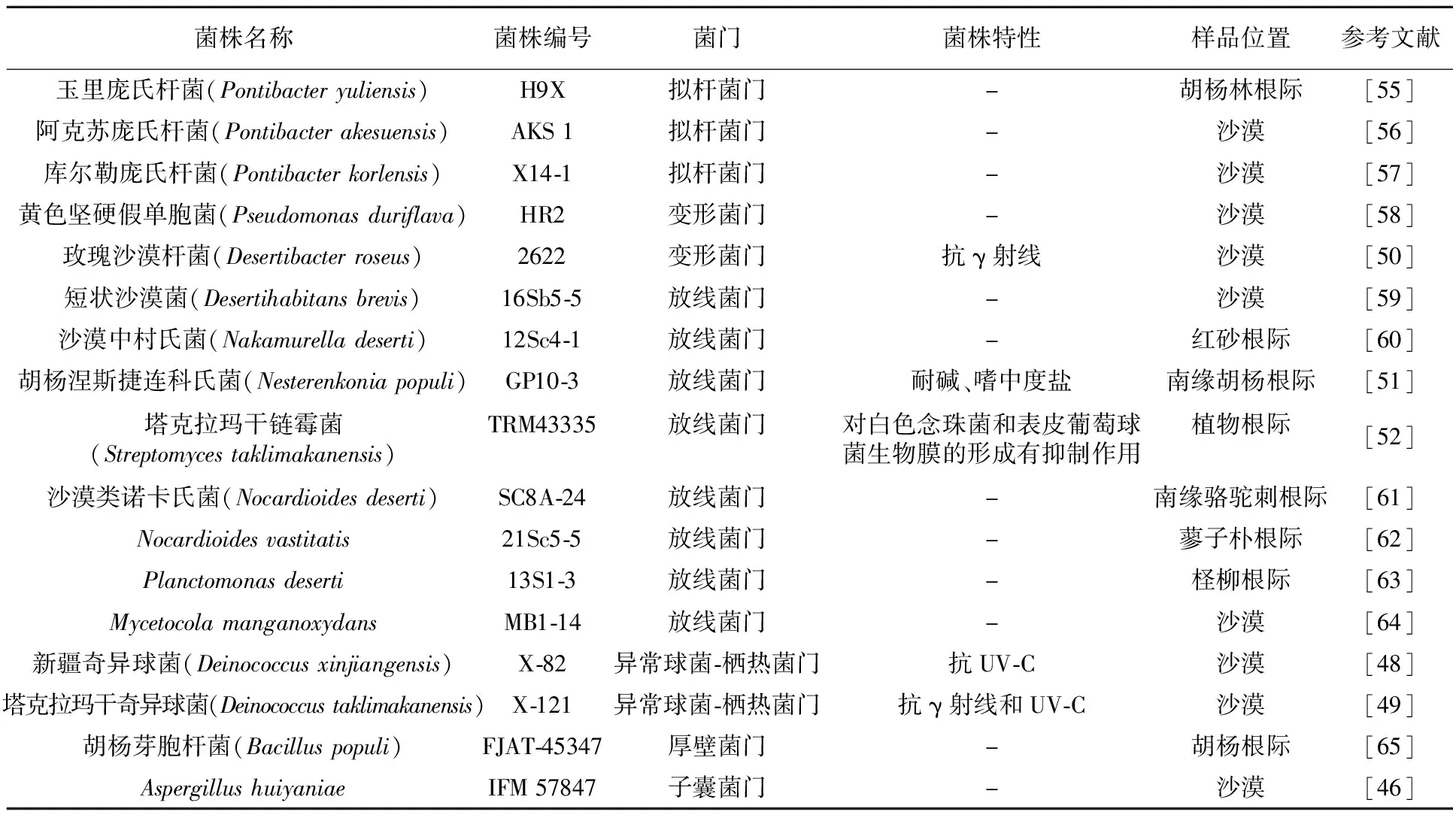
表1 从塔克拉玛干沙漠分离出的微生物新种
从塔克拉玛干沙漠中分离到的新种中大多具有抗辐射能力。异常球菌-栖热菌门的新种菌株,大多数具有抗UV或抗γ射线能力[47]。分离于塔克拉玛干沙漠的新疆奇异球菌(Deinococcusxinjiangensis)具有抗UV的能力,但对γ射线抗性较差[48]。而塔克拉玛干奇异球菌(Deinococcustaklimakanensis)不仅能抗γ射线,而且对UV也具有抗性[49],说明Deinococcus属的不同种具有不同的抗辐射能力,这可能是由于每个菌株都具有独特的抗辐射机制。另外,Liu等[50]从塔克拉玛干沙漠分离到变形菌门的玫瑰沙漠杆菌(Desertibacterroseus)也具有抗γ射线的能力。
除了从塔克拉玛干沙漠分离到抗辐射的新种外,还分离到具有其他功能的新种菌株。如Liu等[51]分离自沙漠南缘胡杨根际的胡杨涅斯捷连科氏菌(Nesterenkoniapopuli)(放线菌门)可耐受碱性环境,并具有中度嗜盐特性。塔克拉玛干链霉菌(Streptomycestaklimakanensis)的次级代谢产物对念珠菌和表皮葡萄球菌生物膜的形成有抑制作用[52]。由于链霉菌能够产生多种具有生物活性的次级代谢产物,如抗肿瘤、抗细菌、抗真菌等,一直是科学家研究的热门种质资源[53],这对于生物、医药和农业等行业都非常重要[54]。
3 沙漠微生物的应用
塔克拉玛干沙漠不仅孕育着丰富的微生物新种资源,而且还有很多具有应用价值的微生物菌株。已有报道[66]从塔克拉玛干沙漠中分离到56株对多环芳烃(PAHs)具有降解能力的菌株,其中50%的菌株能够在两种以上的PAHs培养基上生长,如Arthrobacter、Paenibacillus、Bacillus、Rhodococcus、Sphinomonas和Rhizobium,其中Rhodococcussp.PAH-E-64具有较强的萘降解能力。Luo等[67]分离的MicrovirgaaerilataLM菌株具有降解铅(Pb,浓度1 mmol/L)的能力。罗雪松[66]从该沙漠也分离到一些耐受锰离子的菌株,不同菌株耐受锰离子的能力不同,在80 mmol/L锰离子浓度下能生长的菌株,95%以上都是芽胞杆菌属。
抗辐射菌株广泛分离于各种环境,如核污染地区、沙漠和海洋等[68-70]。沙漠受到极强的太阳辐射,为抗辐射菌株提供了有利的生存条件。研究人员已从塔克拉玛干沙漠分离到很多抗辐射菌株[67,71],通过不同辐照强度可以筛选出抗辐射能力不同的菌株。Luo等[67]经5 000 Gy的γ射线辐照后筛选到1株MicrovirgaaerilataLM菌株。Yu等[10]经3 000 Gy的γ射线辐照后筛选出52株菌株,属于拟杆菌门、变形菌门、异常球菌-栖热菌门、厚壁菌门和放线菌门,这些菌株还具有清除自由基的能力。
沙漠微生物的次级代谢产物具有多种应用价值。Jiang等[71]分离自塔克拉玛干沙漠的菌株黑色短梗霉(Aureobasdiummelanogenum)XJ5-1在生长过程中产生大量黑色素,具有耐紫外线辐射、耐H2O2(200.0 mmol/L)、耐热(40 ℃)、耐盐(200.0 g/L)和耐干旱的能力。菌株XJ5-1的黑色素生物合成是由PKS1基因编码的,黑色素在应激反应中起到重要的作用,但在(NH4)2SO4的存在下,其黑色素生物合成和负责黑色素生物合成的PKS基因的表达明显受到抑制[72]。另外,Jiang等[73]发现菌株黑色短梗霉13-2能高产胞外多糖(EPS),摇瓶培养5 d,EPS产量为(73.25±2.3)g/L,在10 L发酵罐中发酵120 h后,EPS产量为(78.05±3.5)g/L。塔克拉玛干沙漠蕴含着丰富的放线菌资源,该地区具有发现新的放线菌物种及分离出具有抑菌活性菌株的潜力[26]。陈川等[74]对分离自塔克拉玛干沙漠的81株放线菌的次级代谢产物进行铜绿假单胞菌(Pseudomonasaeruginosa)抑菌活性检测,发现具有抗性的主要是链霉菌属,其中对拟诺卡氏菌属的发酵液经过乙酸乙酯萃取、纯化以及质谱检测,表明抗铜绿假单胞菌的化合物是十六元大环内酯类化合物-蔷薇霉素。另外Luo等[75]还分离出抗β-内酰胺酶的菌株寡养单胞菌(Stenotrophomonas)(γ-变形菌门)和生根瘤菌(Mesorhizobium)(α-变形菌门)。从其他沙漠中分离的微生物次级代谢产物的应用研究也有很多报道,如Pervee等[76]分离自沙特阿拉伯沙漠的林氏念珠藻(Nostoclinckia)次级代谢产物粗提物对植物病害真菌黄曲霉(Aspergillusflavus)、黑曲霉(Aspergillusniger)、尖孢镰刀菌(Fusariumoxysporum)、黄青霉(Penicilliumchrysogenum)和枝孢霉属(Cladosporium)有抑菌活性。Schulz等[77]从阿塔卡玛沙漠中分离到的链霉菌可产生具有抗菌、抗肿瘤活性的袢霉素和22-十五环大内酯(22-membered macrolactones)[78-79],以及能够对细菌和皮肤真菌具有抑菌活性的新氨基苯醌类。
4 展 望
随着现代分子技术的发展,更多的沙漠微生物被人们熟知。近年来高通量测序技术[80]、质谱分析学[81]和生物信息学[82]都加快了人们对沙漠微生物群落结构和功能的认知。目前人们已通过测序技术和可培养方法对塔克拉玛干沙漠的细菌、真菌多样性进行研究[19,24,41],但是由于不同测序手段的差异和可培养技术的落后,作为中国最大的沙漠,对其微生物的研究还很少,尤其是有关古菌和病毒的研究。沙漠微生物的研究比其他生态系统困难,一方面可能是测序技术有待提高,另一方面是提取沙漠土壤DNA的难度较大。
对于塔克拉玛干沙漠微生物的研究,尚需关注以下几个方面:①细菌、真菌、古菌和病毒构成了整个沙漠微生物系统,应该去研究它们之间的相互联系和相互影响。②对于沙漠植物与微生物的关系还不是很清楚。例如对于菌根菌与植物的生长是否有直接联系尚不清楚。理解贫瘠环境中营养循环的相互作用,有助于理解沙漠生物多样性对沙漠生态系统功能的影响。③塔克拉玛干沙漠微生物多样性的研究仅仅是基于数量不多的几个采样点来推导和评估整个沙漠的微生物情况。未来的研究应该建立在更多采样点和不同深度以及更大空间和时间尺度上,系统了解塔克拉玛干沙漠的微生物群落组成和功能。④塔克拉玛干沙漠是发掘具有潜在应用价值菌株的宝藏地区,完善可培养方法并分离更多有用的菌株资源,是未来应该重点考虑的研究方向。
因此,全面了解塔克拉玛干沙漠微生物的组成和功能非常必要。尽管宏基因组可以研究微生物的组成和功能,但由于DNA浓度要求较高并且测序费用比较昂贵,完全实现还是比较困难。研究沙漠微生物,对于了解和保护沙漠生态系统、预防沙漠化以及沙漠微生物资源的应用都具有重要意义。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
中国记者(2022年8期)2022-09-15
石油沥青(2022年4期)2022-09-03
中国生物防治学报(2022年3期)2022-07-09
微生物学杂志(2021年2期)2021-07-01
微生物学杂志(2020年2期)2020-12-31
生态学报(2019年11期)2019-07-08
科学导报(2019年3期)2019-06-11
中亚信息(2018年2期)2018-04-26
食品工业科技(2014年23期)2014-03-11
