
基于数据库探讨NEDD8在头颈鳞癌中的表达及其预后价值
2021-11-17 08:55顾姗姗邓红霞沈志森
浙江医学 2021年20期
顾姗姗 邓红霞 沈志森
头颈鳞癌(head and neck squamous cell carcinoma,HNSCC)占头颈部肿瘤的90%以上,是第六大常见的恶性肿瘤,全球每年的新发病例和死亡病例分别超过89万和45万[1-2]。尽管近年来外科手术及放化疗等多种治疗手段有了很大进步,但HNSCC的5年生存率仍仅约50%,超过70%的患者会出现不同程度的复发及淋巴结转移[3-4]。HNSCC是一种异质性上皮肿瘤,与抽烟、饮酒有密切相关性[5]。HNSCC的发病过程较为复杂,涉及到多个分子与细胞的相互作用;且由于其涉及部位多,目前尚缺乏理想的特异性生物标志物。筛选HNSCC相关基因有望为其诊治提供新思路,为其诊断和预后提供候选的生物标志物和治疗靶点。
神经前体细胞表达下调因子8(neural precursor cellexpressed developmentally down regulated 8,NEDD8)是一种类泛素蛋白,最早发现于小鼠大脑中,其含有81个氨基酸,与泛素分子结构最为接近,同源性达到60%[6-7]。NEDD8参与蛋白类泛素化修饰,在调节相关转录因子活性、信号转导和肿瘤发生等生命活动中起重要作用。既往研究表明,NEDD8在肺癌、肝癌、子宫内膜癌等多种肿瘤中异常表达,并与肿瘤的发生、发展及预后判断密切相关[8]。但NEDD8基因在HNSCC中的潜在生物标志物价值尚不明确。微阵列和RNA测序技术的进步为生物医学研究提供了大量数据。本研究通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)和多种公共数据库探索HNSCC患者NEDD8基因的表达、突变和生物学功能情况,预测其在HNSCC中的预后价值,以期为HNSCC预后判断和治疗新靶点选择提供参考。
1 材料和方法
1.1 检索Oncomine数据库 Oncomine(www.oncomine.org)数据库包含715个基因表达数据集和86 733个样本,是全球最大的癌基因芯片数据库和数据挖掘平台[9]。本研究利用该数据库比较NEDD8在HNSCC组织和正常组织中的表达;以改变倍数阈值>1.5,基因排序为前10%,P<0.05为差异有统计学意义。
1.2 检索UALCAN数据库 UALCAN数据库(http://ualcan.path.uab.edu/)是一个TCGA数据库在线分析和挖掘的网站,其基于TCGA基因表达数据提供基因在不同肿瘤亚群中的表达,如性别、年龄、肿瘤分期等,也提供基因生存分析、相关性分析和DNA启动子区甲基化数据分析[10]。本研究通过UALCAN数据库分析HNSCC中NEDD8的表达及其与HNSCC临床病理参数的关系,以P<0.05为差异有统计学意义。
1.3 检索GEPIA数据库 基因表达谱数据动态分析GEPIA 数据库(http://gepia.cancer-pku.cn/)是北京大学开发的用于癌症和正常基因表达谱分析的公共数据库,包括9 736个肿瘤样本和8 587个正常样本的RNA测序表达数据[11]。本研究通过GEPIA数据库分析NEDD8在HNSCC中的生存预后评估价值。
1.4 Kaplan-Meier Plotter分析 采用Kaplan-Meier Plotter(http://kmplot.com/analysis/)在线分析不同NEDD8基因表达水平HNSCC患者的总生存期(overall survival,OS)和无复发生存期(relapse-free survival,RFS)[12]。分别根据NEDD8基因中位表达水平将HNSCC患者分为高表达和低表达两组,采用Kaplan-Meier法绘制生存曲线评估OS和RFS。
1.5 检索cBioPortal数据库 cBioPortal数据库(http://www.cbioportal.org)是一个可视化和分析多维癌症基因组数据的综合性开放平台,目前包含245项癌症研究[13]。本研究利用该平台中488例HNSCC患者的基因表达信息,分析NEDD8基因突变率和患者生存预后情况。
1.6 检索LinkedOmics数据库LinkedOmics(http://linkedomics.org)是一个含有TCGA 32种癌症多组学数据的公共平台,包括 LinkFinder、LinkInterpreter、LinkCompare 3个分析模块[14]。本研究利用该平台的LinkFinder模块研究TCGA HNSC队列(520例)中NEDD8相关的差异表达基因,使用Pearson相关分析进行统计学处理。然后使用LinkInterpreter模块对差异基因进行基因本体论(gene ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG) 通路富集分析。假阳性发现率(false discovery rate,FDR)<0.05,模拟1 000次。
2 结果
2.1 NEDD8 mRNA在HNSCC组织中的表达情况 Oncomine数据库分析结果显示,NEDD8 mRNA在乳腺癌、头颈部肿瘤、淋巴瘤等肿瘤中高表达,而在前列腺癌和白血病等肿瘤中低表达,差异具有统计学意义(P<0.05),见图1(插页)。同时,在HNSCC 6个分析结果中,NEDD8 mRNA均是高表达(均P< 0.05),见表1。

图1 Oncomine数据库常见肿瘤组织中神经前体细胞表达下调因子8(NEDD8)mRNA的表达情况

表1 Oncomine数据库NEDD8 mRNA在HNSCC中表达的差异
通过UALCAN数据库对520例TCGA中HNSCC样本验证发现NEDD8 mRNA的表达水平显著高于正常组织(P<0.05),见图2a。在基于疾病分期、肿瘤分级、淋巴转移分期、患者性别、年龄、种族的亚组分析中,HNSCC患者的NEDD8 mRNA表达上调(P<0.05),见图2b-g。此外,TP53突变的HNSCC患者较TP53不突变患者有着较高的NEDD8 mRNA表达量(P<0.05),见图2h。因此,NEDD8可能是HNSCC潜在的生物标志物。
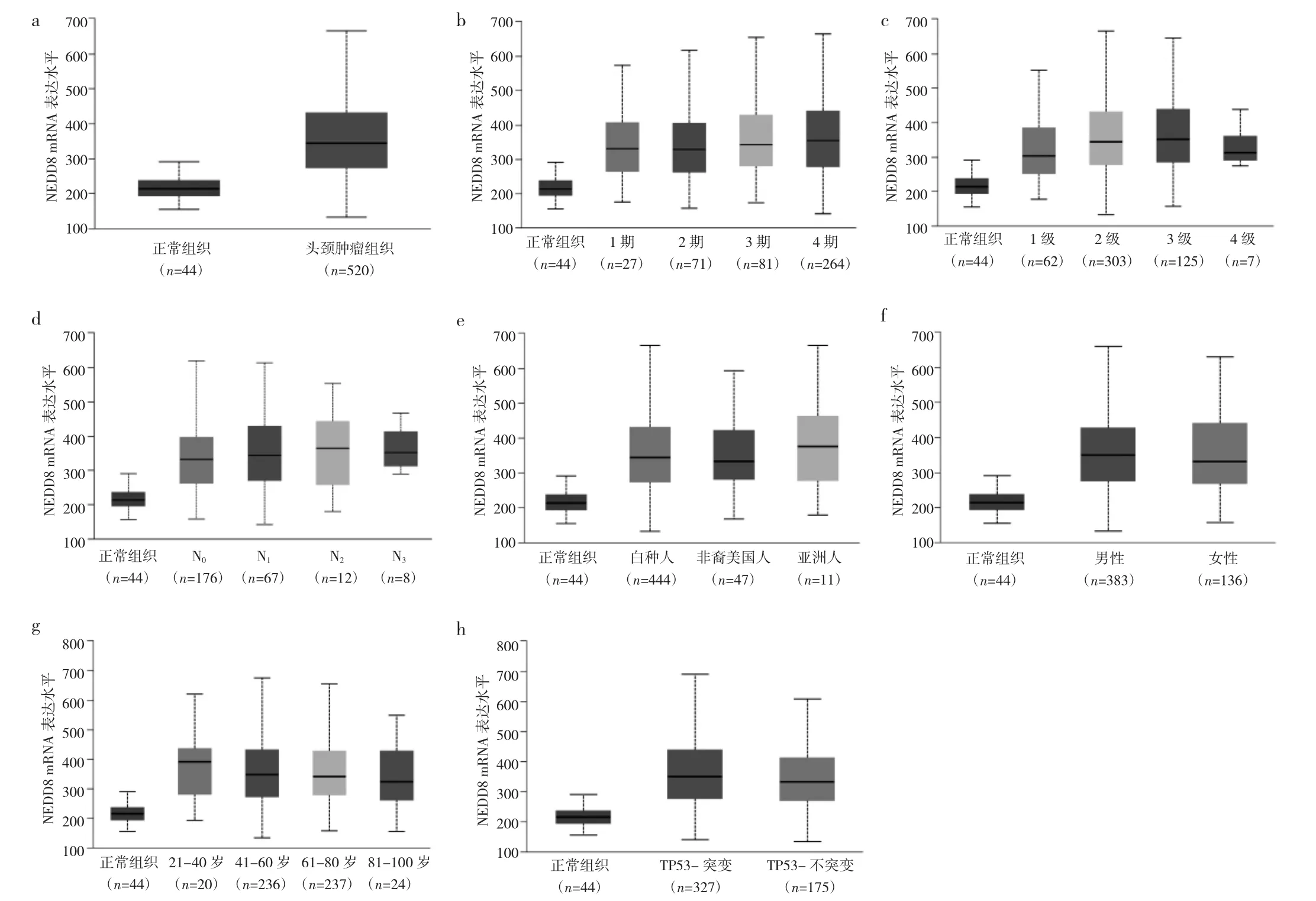
图2 神经前体细胞表达下调因子8(NEDD8)mRNA在头颈鳞癌(HNSCC)组织中的差异表达
2.2 NEDD8 mRNA表达与HNSCC患者预后的关系分析 采用GEPIA数据库和Kaplan-Meier Plotter双重验证NEDD8 mRNA表达与HNSCC患者预后的关系。GEPIA数据库显示,高表达NEDD8 mRNA的HNSCC患者OS偏短[P<0.05,见图3a(插页)],无病生存期更短[P<0.05,见图 3b(插页)]。Kaplan-Meier Plotter在线分析结果显示,同样NEDD8 mRNA高表达组的患者OS显著短于低表达组[P<0.05,见图 3c(插页)],而 NEDD8 mRNA表达与HNSCC患者的RFS未见无明显相关[P<0.05,见图 3d(插页)]。这提示 NEDD8 mRNA 表达与HNSCC的预后相关,高表达的患者预后较差。

图3 不同神经前体细胞表达下调因子8(NEDD8)mRNA表达水平头颈鳞癌(HNSCC)患者生存曲线比较(a、b:GEPIA 数据库分析;c、d:Kaplan-Meier Plotter在线分析)
2.3 NEDD8突变与HNSCC预后的关系分析 基于cBioPortal数据库分析HNSCC测序数据发现,488例HNSCC患者中NEDD8基因发生改变28例,突变率为6%,主要表现为mRNA上调、扩增、深度缺失等,见图4a(插页)。然而,生存曲线图和log-rank检验显示,NEDD8基因突变与HNSCC患者OS[见图4b(插页)]、无进展生存期[见图 4c(插页)]或无病生存期[见图 4d(插页)]均无关(均P>0.05)。

图4 神经前体细胞表达下调因子8(NEDD8)在头颈鳞癌(HNSCC)中的突变和生存分析
2.4 HNSCC中NEDD8共表达基因的GO和KEGG通路分析 使用LinkedOmics数据库分析TCGA中520例HNSCC患者的mRNA测序数据。火山图中用t检验分析NEDD8与HNSCC差异表达基因之间的联系,见图5a(插页)。热图分别显示50个与NEDD8显著正相关或负相关的基因(P<0.05),见图 5b、c(插页)。其中NEDD8 表达与 MRPL52(r=0.8666)、RPL36AL(r=0.8184)、DAD1(r=0.8041)表达均呈正相关(均 P<0.05),见图5d、e、f(插页)。GO通路分析显示与NEDD8相关的差异表达基因主要参与翻译延伸、线粒体呼吸链复合体组装和蛋白折叠等,见图5g(插页);KEGG通路分析显示,其在核糖体、蛋白运输和嘧啶代谢等方面富集,见图5h(插页)。此外,GO和KEGG通路分析得出了与NEDD8相关且最重要的前4个激酶、miRNA和转录因子靶网络,结果表明对激酶靶点、miRNA靶点和转录因子靶点基因集富集得分贡献最大的基因成员数分别是丝裂原活化蛋白激酶1(mitogen-activated protein kinase 1,MAPK1)、GTGCCTT、MIR-506、TTGTTT_V$FOXO4_01,见表2。

图5 LinkOmics数据库分析头颈鳞癌(HNSCC)中神经前体细胞表达下调因子8(NEDD8)共表达基因(GO为基因本体论;KEGG为京都基因与基因组百科全书)

表2 HNSCC中NEDD8的激酶、miRNA和转录因子靶点
3 讨论
NEDD8最早于1992年在小鼠脑组织中被发现,通过类泛素化过程与底物蛋白共价结合,调控其生物性质。类泛素化修饰最具特征的底物是cullin家族蛋白,它是CRLs E3泛素复合体的骨架蛋白[19]。NEDD8也可与许多非 cullin 蛋白底物结合,如 p53、p73、BCA3、VHL,趋化因子受体CXCR5等[20-21]。因此,NEDD8在细胞定位、调节相关转录因子活性和增加蛋白稳定性等方面起重要作用。研究表明NEDD8的过表达及类泛素化的异常激活与人类多种肿瘤、炎症和自身免疫性疾病的发生、发展相关。值得注意的是,在肺腺癌和鳞状细胞癌中,NEDD8通路的大多数酶都被过度激活,且与患者不良预后呈显著相关[22]。但是,目前关于NEDD8在HNSCC中的作用还未明确。因此,本研究利用生物信息学分析NEDD8在HNSCC中的表达、患者临床病理特征和预后价值,以期为今后HNSCC的调控机制和精准治疗研究提供指导。
本研究通过Oncomine数据库挖掘NEDD8在常见肿瘤中的表达情况,发现其在HNSCC中高表达。再次用UALCAN数据库验证发现NEDD8的表达量显著高于正常组织,且与患者临床肿瘤分期、病理肿瘤分级、淋巴结转移分期等显著相关。生存曲线结果表明高表达NEDD8的HNSCC患者OS显著缩短,HNSCC患者预后不良,提示NEDD8可能是一个促癌基因。此外,进一步检索488例HNSCC组织RNA测序结果发现,NEDD8基因在HNSCC中的变异率较低,且变异类型以mRNA上调为主,但其变异与否与患者预后无明显相关性。通过LinkedOmics数据库探索头颈鳞癌中NEDD8共表达基因,并用GSEA进行富集分析,GO分析显示与NEDD8相关的差异表达基因主要参与翻译延伸、线粒体呼吸链复合体组装和蛋白折叠等;KEGG通路分析显示,其在核糖体、蛋白运输和嘧啶代谢等方面富集。随后,本研究用富集分析目标基因集,确定了重要的目标激酶、miRNAs和转录因子网络。本研究结果发现NEDD8在HNSCC中与MAPK1、HCK、MAPK3和ATM激酶相关,这些激酶调控肿瘤信号通路、细胞增殖和细胞周期。研究表明,过表达miR-506通过下调转录抑制因子SNAI2来抑制胃癌细胞的增殖和迁移[23],这为临床了解癌症的发生、发展提供了思路。
综上所述,本研究通过挖掘常见数据库,初步明确了NEDD8基因在HNSCC的表达、变异情况及其预后诊断价值。本研究的结论基于转录水平得出,后续笔者团队将进一步在蛋白水平上对NEDD8的表达及功能进行验证,探究NEDD8调控头颈鳞癌发生、发展的作用机制,为头颈鳞癌的预后判断和治疗新靶点提供理论参考。
猜你喜欢
西北林学院学报(2022年5期)2022-10-04
今日中国·西班牙文版(2022年5期)2022-06-06
云南医药(2021年3期)2021-07-21
现代临床医学(2021年2期)2021-03-29
人人健康(2020年8期)2020-09-02
中等数学(2020年1期)2020-08-24
文化创新比较研究(2020年8期)2020-01-02
科学导报(2019年7期)2019-09-23
医学研究杂志(2015年9期)2015-07-01
都市丽人(2015年4期)2015-03-20
