
马铃薯ERECTA 基因克隆和生物信息学分析
2021-11-15 07:03陈如意李玉双郭志平
湖北农业科学 2021年19期
张 乐,陈如意,李玉双,郭志平
(丽水学院,a.医学院;b.生态学院;c.医务室,浙江 丽水 323000)
ERECTA 是一种富含亮氨酸重复序列的类受体蛋白激酶,广泛分布于各种植物的器官和组织[1]。拟南芥(Arabidopsis thaliana)ERECTA基因参与花药发育,调控细胞增殖和分化[2],ERECTA基因还与植株抗逆性相关,其过表达能够提高植株对高温干旱的耐受性[3,4]。目前,仅在少数经济作物上有EREC⁃TA基因相关的研究[5-7]。
马铃薯(Solanum tuberosum)在世界各地广泛种植,因其具有产量高、适应性强、营养丰富等特点,已成为世界第三大淀粉原料作物和第四大粮食作物[8]。马铃薯除菜用外,近年来,利用马铃薯生产薯片、薯条、淀粉等的产量不断增加。为满足不同需求,育种工作者根据不同区域的气候特点和生产用途,在利用传统方法积极开展马铃薯种质资源研究和杂交育种的同时,积极探索利用现代生物技术在分子水平开展种质资源研究和分子标记育种,目前已取得了一定的成果[9]。本研究通过RT-PCR 技术克隆马铃薯ERECTA基因cDNA 序列,并对其进行生物信息学分析,为进一步开展该基因的功能研究和马铃薯分子育种提供参考。
1 材料与方法
1.1 材料
供试材料为浙江省丽水学院生态学院实验室保存的克新19 马铃薯(种质来自黑龙江省农业科学研究院克山分院)。
1.2 方法
1.2.1 马铃薯总RNA 提取与反转录 总RNA 的提取参照李飞等[10]的方法,取新鲜马铃薯幼苗嫩叶置于预冷的研钵中,同时加入Trizol 试剂,充分研磨并振荡混匀,放置5 min,离心10 min 后取上清液;添加氯仿混匀,室温下放置3 min,离心15 min 取上清液;换至新离心管中,按1 mL Trizol 加0.5 mL 异丙醇的比例配比,静置后离心10 min,弃上清液;按1 mL Trizol 液加入1 mL 以上的75% 乙醇清洗沉淀,涡旋振荡混匀,离心5 min,沉淀在室温下干燥5~10 min,然后将总RNA 溶于DEPC 水。 以提取得到的总RNA 为模板,采用PrimeScriptTMRT reagent 试剂盒进行反转录。
1.2.2 马铃薯ERECTA基因cDNA 克隆 在NCBI数据库中获取马铃薯ERECTA基因cDNA 序列(NCBI登陆号:XM_006353788),设计上游引物:5′-TCAAAGGAGCTGAGTTAGTGCA-3′;下游引物:5′-TCAGCATCAGTGGTCCCAAA-3′。PCR 扩增程序为94 ℃预变性5 min;94℃变性30 s,55.7 ℃退火30 s,72 ℃延伸3 min,共35个循环,最后72 ℃延伸10 min。琼脂糖凝胶电泳检查PCR 扩增产物,割胶回收后送生工生物工程(上海)股份有限公司检测。
1.2.3 生物信息学分析 利用ORF finder(https://www.ncbi.nlm.nih.gov/orffinder/)将马铃薯ERECTA基因的cDNA 翻译成氨基酸序列。使用ProtParam tool(https://web.expasy.org/protparam/)预测等电点、分子质量等理化性质,并通过ProP 1.0 Server(http://www.cbs.dtu.dk/services/ProP-1.0/)分析马铃薯ERECTA蛋白中有无信号肽、前导肽。利用MEGA6.0 构建系统进化树,1 000 次Bootstrap。 用SWISS-MODLE(https://swissmodel.expasy.org/interactive)对马铃薯ERECTA 蛋白进行三维建模。
2 结果与分析
2.1 马铃薯ERECTA 基因cDNA 克隆
利用Trizol 试剂从马铃薯幼苗嫩叶中提取到了总RNA,经Nanodrop 和琼脂糖凝胶电泳确定总RNA浓度和质量(图1A)。随后反转录获得cDNA 第一链,并利用Taq酶体外扩增ERECTA基因cDNA,电泳检测条带大小为3 281 bp,与预期一致(图1B)。
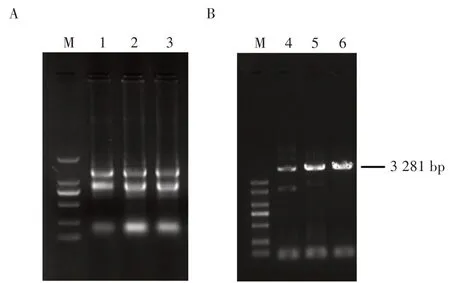
图1 马铃薯ERECTA 基因cDNA 克隆
2.2 马铃薯ERECTA 蛋白分子特征
马铃薯ERECTA基因cDNA 序列包括一个2 973 bp 的开放阅读框,可编码990 个氨基酸。预测分析马铃薯ERECTA 蛋白理化性质,结果显示该蛋白的分子质量为108.95 kDa,理论等电点为5.85。马铃薯ERECTA 蛋白具有一个信号肽,信号肽切割位点位于32 位和33 位氨基酸之间;马铃薯ERECTA不含前导肽(图2)。

图2 马铃薯ERECTA 蛋白信号肽和前导肽预测
2.3 ERECTA 氨基酸序列对比
多序列比对结果表明,马铃薯ERECTA 蛋白的氨基酸序列中含有10 个保守基序,其中8 个为富含亮氨酸的LRR 基序,它们由长度为8~45 的氨基酸残基组成,通常折叠成弧形或马蹄形;ERECTA 有1 个Pkinase 结构域,该结构域是具有催化功能的蛋白激酶结构域(图3)。
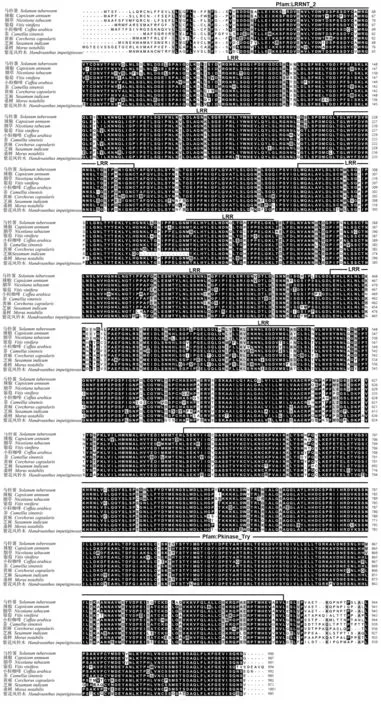
图3 马铃薯ERECTA 及其同源序列的多重比对
2.4 同源性分析
序列同源性分析表明,马铃薯ERECTA与辣椒(Capsicum annuum)ERECTA的 一 致 性 最 高(94.04%),其次是与烟草(Nicotiana tabacum)ERECTA的一致性(93.65%)较高,与黄花风铃木(Handroanthus impetiginosus)ERECTA 的一致性最低(85.03%)。这与多序列比对的结果一致(表1)。

表1 马铃薯ERECTA 与其他植物ERECTA 一致性分析
2.5 ERECTA 蛋白的系统进化树分析
为了分析马铃薯ERECTA 与其他植物ERECTA之间的亲缘关系,利用MEGA 软件构建了系统进化树。结果表明,马铃薯ERECTA 与智利番茄、芝麻ERECTA 聚为一簇,进而和小粒咖啡聚为一簇,说明马铃薯ERECTA 在进化上的保守性(图4)。
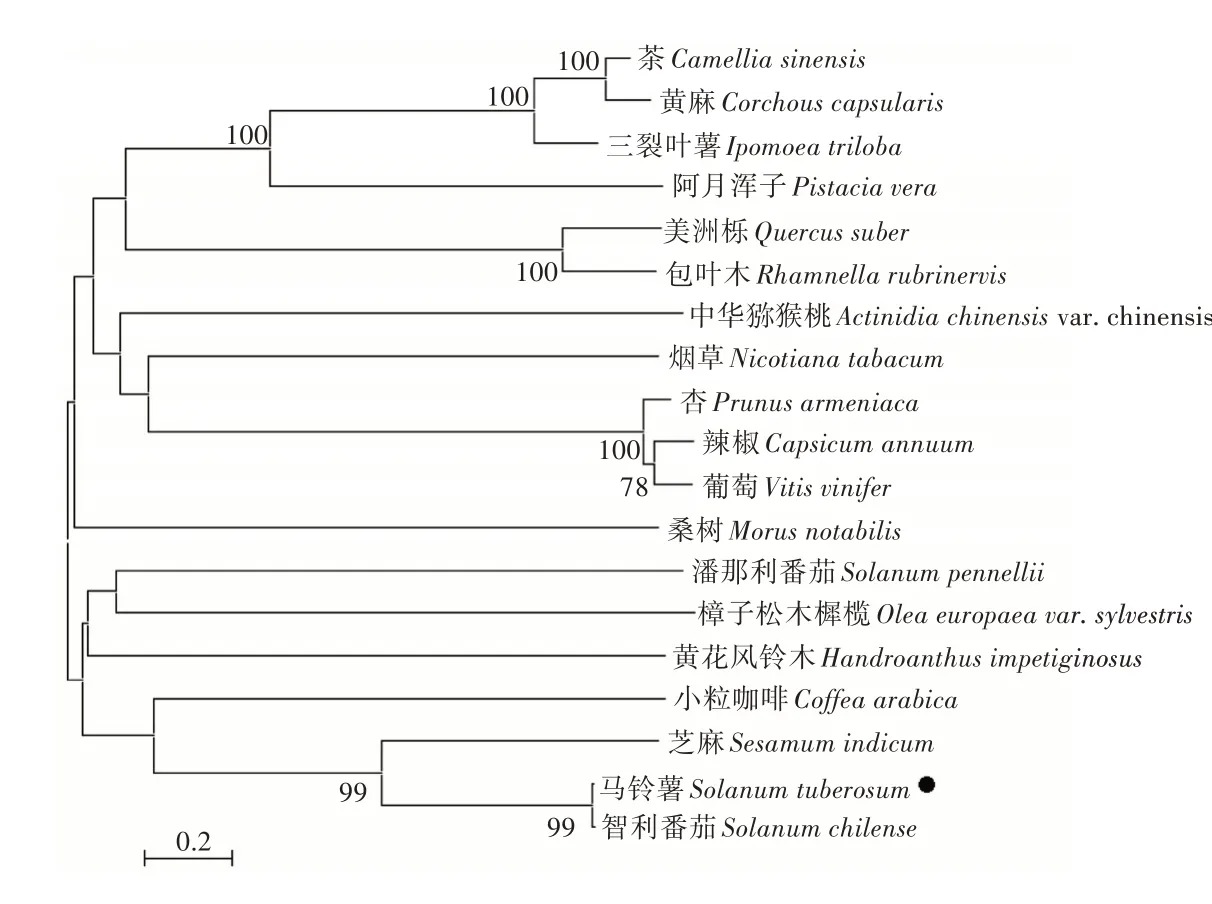
图4 马铃薯ERECTA 与其他植物ERECTA 的系统进化树分析
2.6 马铃薯ERECTA 蛋白质结构分析
马铃薯ERECTA 的二级结构预测结果表明,该蛋白质含34.9% 的α-螺旋,50.9% 的无规则卷曲,14.2% 的β-折叠。三级结构预测结果表明,马铃薯ERECTA 包含8 个富含亮氨酸的LRR 基序(图5)。
3 讨论
ERECTA基因对植株生长发育有多个方面作用,如控制植株的矮化过程[11]。对拟南芥的研究表明,ERETCA 通过调控分生组织的细胞分裂周期实现植株矮化[12,13]。ERECTA基因家族还可以促进细胞增殖和抑制气孔分化,从而促进植物叶片的伸长[14]。相关研究表明,ERETCA基因与植物抗逆性也密切相关,能够提高植物对高温干旱的耐受性[15]。例如高粱(Sorghum bicolor)ERECTA1和ERECTA2基因均在高粱茎和叶中表达,而且其表达水平均随着干旱胁迫程度的加深而逐渐提高[15]。因此,进一步研究ERECTA基因在马铃薯生长发育中的作用,对马铃薯种质研究和育种有指导意义。
马铃薯ERECTA蛋白分子的质量为108.95 kDa,这与Shen 等[3]研究的拟南芥ERECTA 分子质量有所不同,但与辣椒ERECTA 的分子质量相近。通过多序列比对和系统发育分析表明,马铃薯与辣椒等的亲缘关系较近,但与拟南芥的关系较远。研究同时表明,马铃薯ERECTA 与拟南芥ERECTA 在结构和功能上有所差异[7]。ERECTA 为编码富含亮氨酸重复序列的类受体蛋白激酶(LRR-RLK),LRR-RLKs是普遍存在于植物中的信号受体亚家族,它们调控多种信号通路[15]。马铃薯ERECTA 蛋白同样拥有8个LRR 保守基序以及1 个激酶构域,这些保守度较高的基序在其他植物ERECTA 蛋白中也存在[15,16]。马铃薯ERECTA基因的克隆和生物信息学分析,为后续的功能研究和分子育种奠定了基础。
猜你喜欢
新医学(2023年10期)2023-12-09
南方医科大学学报(2022年3期)2022-04-13
天津市教科院学报(2021年5期)2021-11-10
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
生物学通报(2021年9期)2021-07-01
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
淮阴工学院学报(2014年5期)2014-09-10
华南师范大学学报(自然科学版)(2011年2期)2011-11-20
