
甘蓝型油菜花粉粒ARTP 诱变及后代表型鉴定
2021-11-15 07:01何晓莹陆文虎师伟策王文敏王敬乔张国建张建昆束正齐
湖北农业科学 2021年19期
何晓莹,陆文虎,师伟策,王文敏,王敬乔,陈 苇,张国建,张建昆,束正齐,俎 峰
(1.云南省农业科学院经济作物研究所,昆明 650205;2.昆明深洪生物科技开发有限公司,昆明 650051;3.云南省农业职业技术学院,昆明 650031)
甘蓝型油菜大约起源于7 500 年前地中海地区甘蓝和白菜的天然杂交与自然加倍[1],其中油用甘蓝型油菜的驯化与栽培仅有700 余年的历史,是一个年轻的油料作物物种[2]。其有限的栽培驯化史与传统的品种选育方法,尤其是20 世纪70 年代以来的双低化育种导致其遗传基础相对狭窄,致使近年来甘蓝型油菜突破性新品种选育困难,迫切需要创造新的优良种质材料,以拓展甘蓝型油菜的遗传基础与优异基因库[3]。
辐射与化学诱变是传统的种质创新手段。其中辐射诱变多以X 射线、γ 射线、60Coγ 射线与宇宙射线作为辐射诱变源[4],化学诱变多以EMS(甲基磺酸乙酯)为诱变剂[5]。前人利用这些传统技术在甘蓝型油菜中诱变获得了高油酸[6,7]、高含油量[8]、长角果[9,10]、大粒[11]与矮杆[12]等优良突变体。但上述技术多具有人体与环境危害性,限制了其在生产上的大规模应用。基于大气压射频辉光放电原理的常压室温等离子体(ARTP)诱变技术是近年来兴起的一种诱变新方法,具有成本低、安全简便、对环境无害、突变率高、突变体遗传稳定等优点[13,14],前期已广泛应用于各类微生物诱变优良菌株选育中[14-17]。近年来该技术逐渐应用到植物种质创新领域,研究报道显示ARTP 诱变技术能够提高金鸡菊种子发芽势与发芽率,增加幼苗叶片数量与叶面积,提升植株花朵的次生代谢产物[18],促进玉米种子萌发与幼苗生长[19],有助于提升甘蔗愈伤组织草甘膦抗性能力[20],但未有ARTP 在油菜种质创新领域的应用研究报道。
本研究以甘蓝型油菜花粉为诱变材料,采用ARTP诱变技术分别辐照0、20、30 和40 min,再将诱变后的花粉授于同一群体中自然产生的不育单株,探究不同辐照时间对诱变花粉授粉当代单角粒数及后代子粒发芽势、发芽率、不育株率及含油量的影响。探索性地开展甘蓝型油菜ARTP 花粉粒诱变试验,以期获得高含油量变异株系,并为今后生产上应用ARTP花粉粒诱变技术探索较为适宜的参数体系及为开展油菜种质创新提供新的方法与思路。
1 材料与方法
1.1 试验材料
甘蓝型油菜Ogu-CMS 恢复系16C 由云南省农业科学院经济作物研究所油菜研究中心提供(16C外源恢复基因不完全符合孟德尔遗传,纯合可育单株自交后代群体中会自然产生约4% 的不育单株-16CS)。2018 年10 月种植16C 于寻甸基地网室;2019 年5 月收获诱变花粉授粉后代-M0代种子;2019 年10 月种植M0代种子于小哨基地网室;2020年2 月田间调查4 个ARTP 辐照时间梯度后代群体不育株率并于2020 年5 月收获M1代种子。
1.2 试验方法
1.2.1 ARTP 诱变 利用ARTP 诱变育种仪(ARTPP,无锡源清天木生物有限公司)使用99.99% 高纯度氦气作为工作气体对油菜花粉粒进行辐照处理。辐照参数设置如下:额定功率,160 W;工作气量,15 SLM;等离子发射源照射样品距离,2 mm;辐照时间分别为0、20、30 和40 min,以CK、ARTP20、ARTP30和ARTP40 表示。
1.2.2 花粉取样与诱变后授粉 2019 年1 月17 日,16C 盛花期选取当日开花的花朵,使用小毛刷仔细刷取花粉到诱变载盘,花粉厚度<1 mm。花粉粒ARTP 诱变后,诱变载盘置于冷藏柜(4 ℃)保存过夜。第2 天中午授粉于寻甸基地网室16C 群体中的不育单株-16CS(不育单株提前3 d 套袋,避免花粉污染)。每个辐照时间梯度花粉授粉5~10 株,2019年5 月收获M0代种子。
1.2.3 M0代种子与植株测定指标及其方法 诱变花粉粒授粉当代植株单角粒数:每个辐照时间梯度选取5 个单株,每个单株选取主花序20 个角果进行单角果粒数统计。
M0代种子发芽势与发芽率:把M0代种子播种在0.2% 琼脂糖培养基上,室温放置,分别统计第3 天和第6 天的萌发种子数,计算发芽势与发芽率。重复3次,每个诱变时间梯度每重复播种100 粒种子。
发芽势=(第3 天萌发种子数/每组试验种子数)×100%
发芽率=(第6 天萌发种子数/每组试验种子数)×100%
M0代植株不育株率调查:M0代植株盛花期,鉴定单株育性。
不育株率=(不育单株数/总单株数)×100%
1.2.4 M1代种子含油量测定 使用近红外(NIRS)品质分析仪(FOSS5000,丹麦福斯公司)对M0代植株开放结实获得的M1代种子进行含油量分析,具体操作参考前人报道[21]。
1.2.5 数据分析 数据采用均值±标准差表示。使用Excel 2016 进行单因素方差分析。
2 结果与分析
2.1 ARTP 辐照油菜花粉对当代授粉植株单角粒数的影响
用ARTP 辐照0、20、30 和40 min 的花粉给16CS植株授粉,单角粒数分别为29.74±0.90、29.13±1.54、23.95±2.02 与19.55±1.13。可见随着ARTP 辐照时间增加,角粒数逐渐下降。其中ARTP20 处理的角粒数与对照(CK)相比基本没有变化;ARTP40 授粉单角粒数最低,较CK 降低34.26%。方差分析结果见表1。由表1 可知,不同ARTP 辐照处理之间单角粒数差异极显著(F值=53.88**),推测ARTP 辐照能够显著降低花粉粒活性和授粉结实能力,并进而影响角粒数。

表1 不同ARTP 辐照时间方差分析
2.2 ARTP 辐照油菜花粉对M0 代子粒发芽率与发芽势的影响
M0代子粒3 重复发芽试验结果见图1。由图1可知,CK、ARTP20、ARTP30 和ARTP40 诱变花粉授粉所得M0代子粒发芽势分别为96.00%±1.00%、37.00%±2.65%、0.00%±0.00% 与13.67%±2.52%;发芽率分别为100%±0.00%、73.67%±3.79%、12.33%±3.06% 与27.67%±2.52%,表明ARTP 辐照显著影响M0代子粒的发芽势与发芽率。ARTP30 的M0代子粒发芽势与发芽率最低,其中发芽势低至0,而发芽率均值也仅有12.33%。但是,ARTP40 的M0代子粒发芽势与发芽率较ARTP30 均有所上升,结合“2.1”结果,推测这可能是由于ARTP40 辐照诱变载盘上的花粉层厚度不均匀所致,虽然ARTP 辐照40 min 极大降低了花粉活力与授粉能力,但部分较厚花粉层底部的花粉粒未得到ARTP 充分辐照,进而导致其后代M0代子粒发芽势与发芽率较ARTP30 有所提升。

图1 M0代子粒发芽势与发芽率
2.3 ARTP 辐照油菜花粉对M0代植株不育株率的影响
分别对CK、ARTP20、ARTP30 与ARTP40 的M0代植株取样192、768、812 和837 株进行育性情况调查,其中不育株数分别为41、191、319 和265 株。ARTP20 的M0代不育株率为24.87%,比CK 不育株率(21.35%)略高;ARTP30 的M0代不育株率最高,达39.29%;ARTP40 的M0代不育株率次之,达31.66%(图2)。由此可见,ARTP 辐照花粉时长≥30 min 显著影响16C 花粉粒外源恢复基因的遗传稳定性。但同时不同辐照时间梯度对不育株率的影响表现出与“2.2”类似的结果,即ARTP40 M0代植株不育株率低于ARTP30 M0代植株不育株率,推测亦是由于ARTP40诱变载盘上的花粉层厚度不均匀所致。
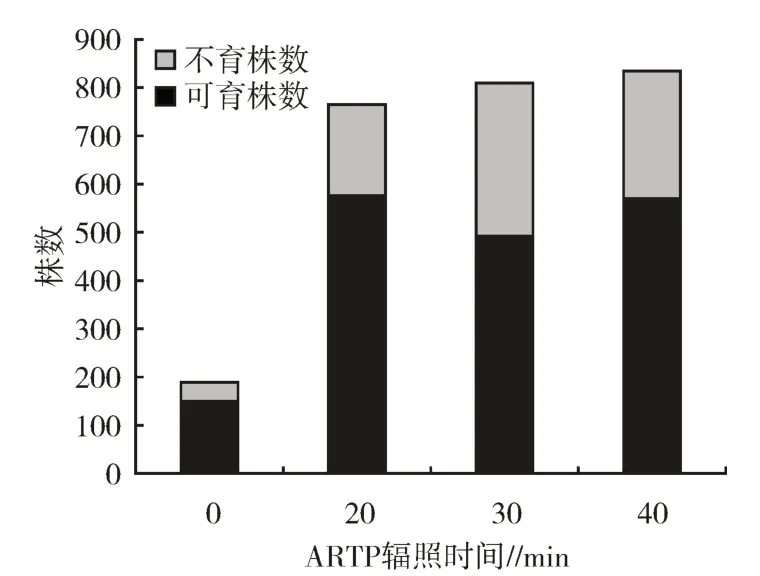
图2 M0代植株不育株率统计
2.4 ARTP 辐照油菜花粉对M1代子粒含油量的影响
随机收获314 株ARTP30 和15 份CK 的M0代植株,利用近红外品质仪对其M1代子粒进行含油量分析。结果显示10 株CK M1代子粒含油量均值为41.39%±0.75%,与历年来16C含油量均值41.17%±1.89%基本一致。而314株ARTP30 M1代子粒中含油量有287株超过42.50%、221株超过45.07%和43株超过50.20%,分别占比91.40%、70.38% 与13.69%;其中ARTP30-315 单株子粒含油量最高,达52.61%,较CK 含油量均值增幅27.11%。由此可见,ARTP30 的M1代油菜子粒含油量较CK 与16C 得到了显著提升。
3 讨论
本研究采用ARTP 诱变技术以4 个时间梯度(0、20、30、40 min)辐照油菜花粉,发现ARTP 辐照30 min 以上的诱变花粉,授粉当代单角粒数及后代M0代子粒发芽势和发芽率均显著下降,M0代植株不育株率显著上升;同时发现ARTP30 M1代子粒含油量较CK 有显著提升(43 份子粒含油量超过50.2%,占比13.69%),其中ARTP30-315 单株子粒含油量最高,达52.61%,较CK 含油量均值增幅27.11%。研究初步明确,利用ARTP-P 诱变仪开展甘蓝型油菜花粉诱变较为适宜的ARTP 辐照时间区间,并获得了大量含油量显著提升的株系,为今后生产上应用ARTP 花粉粒诱变技术及开展油菜种质创新提供了有益的尝试与技术参数借鉴。
较种子、幼胚及愈伤组织而言,花粉粒更适宜作为种质创新的诱变材料[5]。植物花粉量巨大,且花粉诱变产生突变后,通过双受精作用能够快速将遗传变异传递给后代,但前人研究报道却较少使用花粉作为诱变对象[4]。这主要是因为对于完全花作物而言,花粉诱变后授粉给原株系材料需要进行大量的人工剥蕾去雄;而授粉给不育系株系,则后代子粒基因型杂合,导致后续突变体与突变表型鉴定困难[22-25]。本研究采用Ogu-CMS 恢复系16C 作为花粉供体,其群体中自然产生的不育单株16CS 为诱变花粉授粉对象,既不需要大量手工剥蕾去雄,同时授粉后代的遗传背景亦没有发生改变,方便后续突变体与突变表型的比较与鉴定,较好地解决了上述问题。
甘蓝型油菜Ogu-CMS 恢复系16C 为云南省主推杂交品种云油杂15 号父本,其在生产应用过程中存在两个问题需要改良与解决。一是16C 含油量偏低,多年多点含油量测试均值在42% 左右,制约了云油杂15 号的产油量;二是恢复系16C 纯合自交后代出现约4% 的不育单株,影响杂交制种产量与纯度。本研究利用16C 花粉作为诱变材料,在辐照30 min 的后代中发现了43 份子粒含油量超过50.2%的单株材料,后续这些株系材料通过纯化与筛选有望解决16C 含油量偏低的问题。同时本研究发现16C 为处理对照M0代不育株率为21.35%,这与16C纯合自交后代4% 的不育株率一致。 ARTP 辐照30 min 以上,M0代植株不育株率显著上升,最高达39.29%,表明16C 花粉外源恢复基因携带的染色体片段受到了ARTP 辐照的诱变,预示着有望在其后代中选育出育性遗传更为稳定的恢复株系。未来研究中,需要进一步加强高含油量、育性更加稳定遗传的突变材料的筛选,这样的突变株系有望替代原有的16C 株系进行杂交种配制与生产。
猜你喜欢
作物杂志(2021年6期)2021-12-17
植物研究(2021年5期)2021-03-02
热带农业科技(2020年4期)2020-10-23
生物灾害科学(2019年2期)2019-07-17
落叶果树(2018年6期)2018-11-28
吉林林业科技(2015年5期)2015-09-17
东北林业大学学报(2015年2期)2015-03-10
植物营养与肥料学报(2012年5期)2012-10-26
植物营养与肥料学报(2012年3期)2012-10-26
植物营养与肥料学报(2010年1期)2010-11-02
