
水稻低温发芽力QTL qLTG3-1 基因内分子标记的开发及其在华南籼稻中的应用评价
2021-11-12 11:34杨梯丰张子怡董景芳张少红赵均良
广东农业科学 2021年10期
杨梯丰,张子怡,董景芳,周 炼,张少红,刘 斌,赵均良
(广东省农业科学院水稻研究所/广东省水稻育种新技术重点实验室/广东省水稻工程实验室,广东 广州 510640)
【研究意义】随着我国经济快速发展,农村主要劳动力向城市转移,劳动力短缺已成为水稻生产的突出问题。采取轻简化、机械化的生产方式,是解决这一问题的根本出路。直播是一种轻简、高效的栽培模式,近年来发展迅猛。在双季稻地区,强的低温发芽力是保证早季直播水稻形成整齐、健壮秧苗的关键因素之一。但目前许多主栽水稻品种低温发芽力低[1],已经成为限制直播稻发展的关键 因素。选育耐低温发芽力强的水稻品种是推动水稻直播发展的关键。但水稻低温发芽力是多基因控制的数量性状[2],直接选择的效率很低,应用常规遗传育种技术难以进行有效育种。在水稻中挖掘低温发芽力优异基因并加以应用,将是水稻低温发芽力育种取得突破的关键。近年来,分子标记辅助选择已经被广泛应用到水稻育种中[3-8]。相比基因的连锁标记,位于基因内的分子标记可以有效避免遗传累赘,使目标基因的选择更高效、准确。因此,基因内分子标记的应用可提高选择的准确性,从而加快分子育种进程。
【前人研究进展】分子标记技术的发展为水稻低温发芽力的遗传基础解析提供了一种有效工具。自分子标记技术发展以来,国内外对水稻低温发芽力QTL 的标记定位已有一些报道。国内学者最早以典型的籼粳交(窄叶青8 号/京系17)的DH 群体为材料,用15 ℃低温处理种子16 d,鉴定到2 个低温发芽力QTL(qLTG9和qLTG4),其贡献率分别为11.1%和12.6%[9]。随着分子标记技术的成熟,利用双亲遗传作图群体对水稻低温发芽力的遗传解析逐渐增多[10-28]。近年来,全基因组关联分析(GWAS)作为一种高效的水稻QTL 鉴定方法,也被应用于水稻低温发芽力的定位中[29-36]。目前,通过双亲作图群体标记定位的水稻低温发芽力QTL 已超过110 个,通过GWAS 分析检测到与低温发芽力相关的位点多达174 个,这些 QTL 分布在水稻 12 条染色体上,其中2 个已被克隆,分别是qLTG3-1[37]和OsSAP16[38]。
qLTG3-1是最早被克隆的低温发芽力QTL,其功能基因Os03g0103300编码一个由184 个氨基酸组成的功能未知蛋白。在品系“Hayamasari”中该基因编码区71 bp 的缺失是导致低温发芽力出现差异的原因[37]。通过对62 份日本水稻核心种质中1 735 bp 的qLTG3-1区域测序比较,发现在编码区有1 个非同义替换(T/A)以及3 个插入缺失(18 bp 和9 bp 缺失,9 bp 或36 bp 插入),共有10 种等位基因,但在62 份水稻种质中未发现71 bp 缺失的变异类型[39]。最近,Shim 等[40]通过对2 个有低温发芽力差异的品种进行序列分析发现,在qLTG3-1基因的编码区存在3 处可导致氨基酸变异的差异(核苷酸替换变异T/A、CGG/TTC 和18 bp 缺失),并针对这3 处变异设计了引物对98 份亚洲栽培稻的基因型进行分析,根据这3 处编码区的变异以及71 bp 的序列缺失,将该基因划分为5 种单倍型。
【本研究切入点】Shim 等[40]发现18 bp 缺失(GGCGGTGGCGGTGGCGGT)以 及CGG/TTC变异与低温发芽力显著相关,带有18 bp 缺失的qLTG3-1有较强的低温发芽力。本项目组在对单片段代换系文库亲本的低温发芽力评价时发现,南洋占的低温发芽力显著强于华粳籼74[41];通过比较这两份品种的基因组重测序信息发现,南洋占带有在编码区+127 位无18 bp 缺失的qLTG3-1,而华粳籼74 带有在编码区+127 位有18 bp 缺失的qLTG3-1。阐明“18 bp 缺失能否标示qLTG3-1的低温发芽力强弱”这个问题,将为在水稻中利用qLTG3-1开展低温发芽力的分子育种提供借鉴。【拟解决的关键问题】虽然Shim等[40]根据测序结果,设计了扩增qLTG3-1中编码区+127 位18 bp 差异的引物,但是用这对引物对试验材料进行扩增,出现了3 种扩增带型,序列分析显示第3 种带型是因为其所用的5 个籼稻品种既含有18 bp 缺失,又增加了36 bp 插入,而36 bp 插入可能是籼稻中的一个特殊变异。可见,该引物无法特异区分该区域的18 bp 变异。因此,需要设计特异扩增这18 bp 变异的分子标记来进行基因型和低温发芽力表型对应关系的分析。本研究设计了可以特异检测qLTG3-1中这18 bp 变异的分子标记Gltg3-1,并对该标记在华南籼稻的检测效果进行了评价。
1 材料与方法
1.1 试验材料
1.1.1单片段代换系及亲本 供体亲本南洋占、受体亲本华粳籼74(籼稻)以及含有南洋占qLTG3-1的单片段代换系1037 由广东省植物分子育种重点实验室提供[42]。单片段代换系是通过多代回交以及分子标记辅助选择创制的一种特殊遗传材料。它与受体亲本之间只存在1 个已知代换片段的差异,所有单片段代换系与受体亲本之间可遗传的表型差异都与这个代换片段相关[43]。单片段代换系1037 带有来源于南洋占的第3 染色体的代换片段,代换区间为:短臂末端--RM14259-RM14312-RM14380-RM14427--RM175(0~3.8 Mb)。南洋占、华粳籼74 的qLTG3-1部分序列差异见图1。
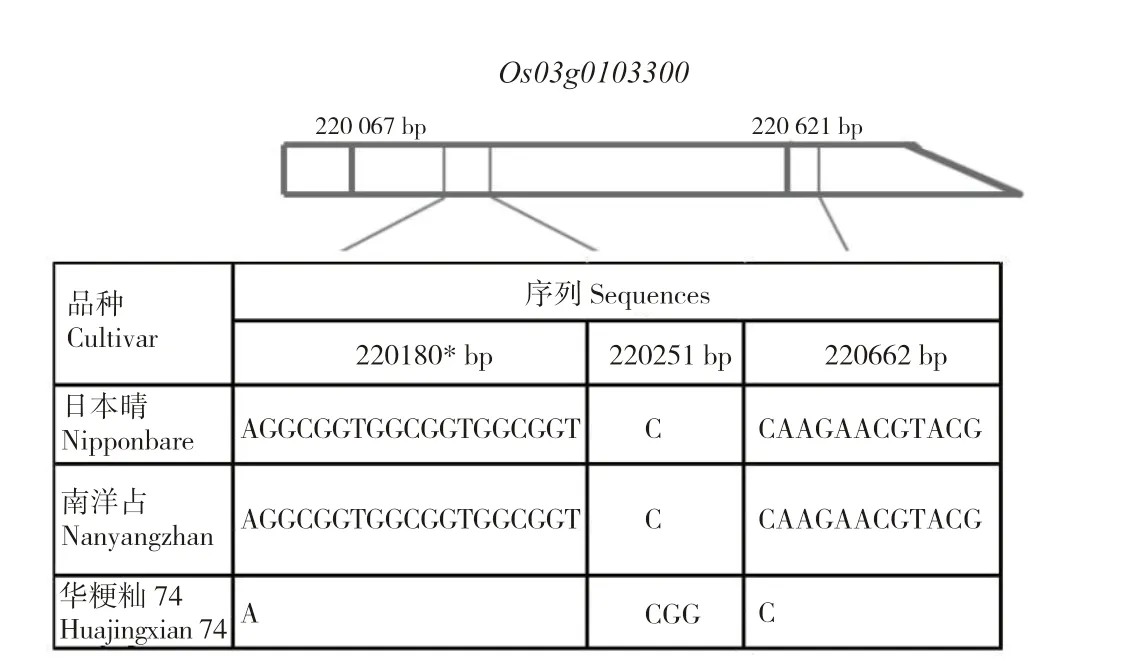
图1 南洋占、华粳籼74 的qLTG3-1 部分序列差异Fig.1 Partial sequence variations of qLTG3-1 in Nanyangzhan and Huajingxian74
1.1.2籼稻品种/系 以111 份国内籼稻品种/系为研究材料(表1),2019 年晚季种植于广东省农业科学院大丰试验基地,7 月23 日播种,8月8 日移栽,每个品种/系种植4 行,每行8 株,株行距为19.8 cm× 19.8 cm,采用常规栽培和水肥管理。种子成熟后,按株系收获种子,晒干,室温存储3 个月后用于低温发芽力评鉴。
表1 111 份华南籼稻品种/系Table 1 111 indica varieties/lines from South China
1.2 试验方法
1.2.1低温发芽力评价 参照Yang 等[36]的方法,略作修改。每个水稻品种/系取240 粒饱满的种子置恒温箱内,49 ℃处理96 h 以打破休眠。用3%次氯酸钠消毒20 min,蒸馏水冲洗干净,每80 粒种子排列于1 个铺有滤纸的9.0 cm 培养皿内,加入 12 mL 蒸馏水,置于15 ℃生长箱中进行低温发芽试验,设置3 个重复。以芽长或根长≥1 mm为萌发标准,在萌发10 d 时检测种子的发芽率,然后调整温度为26 ℃,4 d 后再次调查各品种/系的发芽率,以低温发芽力作为种子低温发芽的指标。

1.2.2总DNA 提取 DNA 的微量抽提采用TPS法[44]。具体过程如下:于分蘖盛期取上部长约2~4 cm 的叶片放入2 mL 离心管中,加入钢珠及800 μL TPS(100 mmol/L Tris-HCl、10 mmol/L EDTA、1 mol/L KCL)抽提液,用研磨仪震荡研磨,然后将其放入75 ℃水浴锅中水浴20 min,12 000 r/min 离心10 min,取上清液约500 μL 于1.5 mL 离心管中,加入等体积的冰乙醇至DNA 析出,12 000 r/min 离心10 min,去上清,取沉淀,晾干,加灭菌双蒸水200 μL 至沉淀溶解。
1.2.3引物设计 针对qLTG3-1的18 bp 缺失,从RAP-DB(https://rapdb.dna.affrc.go.jp/)下载以日本晴为参考基因组的Os03g0103300序列。找到目标变异位点,截取上下游各250 bp 序列,用Primer Premier 5 软件设计引物,并通过NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)的Primerblast 进行引物特异性检测,选取特异性好的引物。
1.2.4PCR扩增和电泳检测 PCR扩增参照Yang等[6]的方法。15 μL反应体系中包含0.15 μmol/L引物、200 μmol/L dNTP、1×PCR反应缓冲液(50 mmol/L KCl,10 mmol/L Tris-HCl、pH8.3,1.5 mmol/L MgCl2,0.01%明胶)、50~100 ng DNA模板、1UTaq酶。PCR反应在9700型DNA扩增仪中进行,反应程序为:94 ℃预变性5 min;94 ℃30 s、55 ℃ 30 s、72 ℃ 40 s,35次循环;72 ℃延伸8 min。扩增产物用8.0%聚丙烯酰胺凝胶电泳分离,经Goldview染色后用凝胶成像系统(美国Bio-rad公司)检测带型。
2 结果与分析
2.1 不同qLTG3-1 等位基因水稻的低温发芽力比较
为检验南洋占带有的qLTG3-1能否提升水稻的低温发芽力,从单片段代换系库中选取以华粳籼74 为遗传背景,对携带来源于南洋占qLTG3-1的单片段代换系1037(代换片段为第3 染色体短臂端0~3.8 Mb 区间)以及南洋占进行低温发芽力评价。结果(图2)表明,单片段代换系1037的低温发芽力显著高于华粳籼74,而南洋占的低温发芽力显著高于1037。可见,来源于南洋占的qLTG3-1能显著提高华粳籼74 的低温发芽力。

图2 来源于南洋占的qLTG3-1 对水稻低温发芽力的影响Fig.2 Effect of qLTG3-1 from Nanyangzhan on low temperature germinability in rice
2.2 qLTG3-1 基因内分子标记的开发
从RAP-DB 网站下载日本晴的Os03g0103300序列信息。根据文献[39-40],在缺失18 bp 序列(GGCGGTGGCGGTGGCGGT)的位置两侧,设计出特异性较好的可扩增该区域的分子标记Gltg3-1 和Gltg3-2(表2)。预测对没有18 bp 缺失的序列,分别可扩增出147、247 bp 的片段(R带型);而对存在18 bp 缺失的序列,则扩增出129、229 bp 的片段(D 带型)。

表2 两对引物的序列和扩增产物信息Table 2 Sequences and amplification products of two pairs of primers
2.3 分子标记的筛选
利用Gltg3-1 和Gltg3-2 引物,随机选取30 份籼稻(材料编号1~30,表1)进行扩增。Gltg3-1 扩增出两种带型,即存在和不存在18 bp缺失的带型;而 Gltg3-2 扩增出4 种带型(图3)。这可能是由于Gltg3-2 扩增区间较大,在扩增区间内还存在其他序列变异[39]。可见,Gltg3-1 能特异区分qLTG3-1的18 bp 差异。
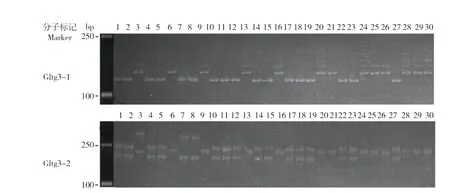
图3 Gltg3-1 和Gltg3-2 标记的扩增带型Fig.3 Amplification band patterns of two molecular markers(Gltg3-1 and Gltg3-2)
2.4 Gltg3-1 标记对华南籼稻qLTG3-1 的检测效果
利用分子标记Gltg3-1 对111 份籼稻DNA 进行扩增分型,结果显示,51 份含有18 bp 缺失(D带型),53 份无18 bp 缺失(R 带型),7 份为杂合型(图4A)。
对qLTG3-1纯合的品种/系进行低温发芽力测定,在51 份D 带型的品种/系中,4 份发芽力低于30.0%,其余47 份品种/系的发芽力为38.1%~97.5%,平均值为74.0%;在53 份R带型的品种/系中,2 份发芽力低于30%,其余51 份品种/系的发芽力为49.4%~95.7%,平均值为79.0%;但两组低温发芽力差异不显著(图4B)。结果表明,该分子标记能区分存在18 bp 缺失和无18 bp 缺失的qLTG3-1,但是这种带型差异在华南籼稻中与低温发芽力强弱无对应关系。
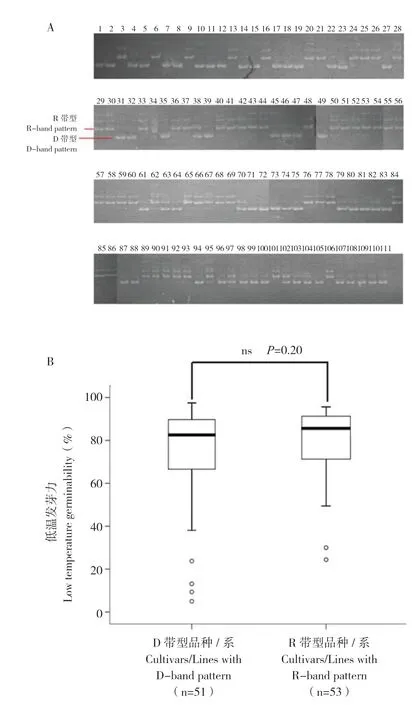
图4 Gltg3-1 标记的扩增带型(A)及不同扩增带型品种/系的低温发芽力(B)Fig.4 Band patterns amplified by molecular marker Gltg3-1 and low temperature germinability of varieties/lines with different band patterns
3 讨论
qLTG3-1是控制水稻的低温发芽力的QTL[37];在Hwaseong/IRGC105491 构建的近等基因系群体中也发现qLTG3-1控制水稻的低温发芽力,而且与QTLqLTG1无显著互作,带有这两个QTL 的株系低温发芽力显著高于只带有单个QTL的株系[40]。在本研究中,带有来源于南洋占的qLTG3-1能显著提高华粳籼74 的低温发芽力。可见,qLTG3-1是1 个控制水稻低温发芽力的重要QTL。
在找到功能基因后确定其功能位点将大大提升该基因在分子育种中的利用效率。已有的等位基因分析表明,qLTG3-1存在多种复等位基因[39-40]。在本研究中,通过比较南洋占和华粳籼74 的qLTG3-1发现,在编码区有18 bp 缺失以及2 bp插入,但是没有检测到其他变异位点。可见,qLTG3-1的序列变异丰富,存在多种类型的等位基因。在这些序列变异中,+127 位的18 bp 缺失在多个研究试验中都存在,可见这个变异在水稻种质中广泛存在。
为了探究这18 bp 变异是否与低温发芽力表型显著相关,本研究设计分子标记Gltg3-1 特异扩增该位点,通过扩增带型将品种/系分组进行低温发芽力差异分析,结果发现两组的低温发芽力无显著差异,即qLTG3-1的18 bp 变异并不能标示低温发芽力的强弱。这一结果与Shim 等[40]的结论不一致,可能的主要原因:一是因为Shim等进行低温发芽力表型鉴定的材料是韩国的核心稻种,大部分为粳稻,所有出现18 bp 缺失带型的都是籼稻(含2 份aus 稻),共27 份;而无缺失带型的都是粳稻,共42 份;而本研究所用的111 份试材均是华南稻区的籼稻,扩增带型显示华南籼稻中也广泛存在这18 bp 变异,其中51 份为缺失带型,53 份为无缺失带型。基于有限的试验材料得到的结果有一定局限性。二是由于水稻的低温发芽力是多基因控制的复杂性状,还有一些重要的低温发芽力QTL 在发挥作用。从文献报道的QTL 定位结果来看,通过作图群体定位到大量的低温发芽力QTL,绝大多数QTL 都位于不同的染色体区间[9-35]。可见,在不同遗传材料中,往往存在多个控制低温发芽力的QTL,QTL 位置一般并不相同,单个QTL 的效应比较小。在本研究中,单片段代换系1037 的低温发芽力显著高于华粳籼74,而南洋占的低温发芽力显著高于1037。可见,在南洋占中也存在多个控制低温发芽力的QTL。
在Shim 等[40]研究中,来源于O.ruf ipogon缺失了18 bp 的qLTG3-1等位基因能显著增强低温发芽力;在本研究中,来源于南洋占带有无缺失的qLTG3-1能显著增强低温发芽力。这进一步证实,qLTG3-1的18 bp 变异无法区分低温发芽力的高低。因此,在水稻低温发芽力改良中,需要对qLTG3-1等位基因的效应进行评价,获得增强水稻低温发芽力的等位基因用于品种改良。在本研究中,鉴定到来源于南洋占的qLTG3-1能显著提高华粳籼74 的低温发芽力,该等位基因可用于提高水稻低温发芽力的改良。
分子标记辅助选择育种是利用与目标QTL紧密连锁或者来自目标基因内部的分子标记,对杂交后代的基因型进行准确判别,进而辅助选择目标性状。它不受外界环境的干扰,能显著提高育种效率,已成为水稻遗传改良的重要工具。开发简便、经济和实用的分子标记是高效开展水稻分子标记辅助选择育种的基础。本研究依据在qLTG3-1复等位基因中广泛存在的18 bp 变异设计了分子标记Gltg3-1,该分子标记存在于基因内部,可以高效准确地在杂交后代中选择到目标基因,这将在水稻低温发芽力分子育种中发挥作用。
本研究仅对在qLTG3-1中广泛存在的18 bp变异与低温发芽力表型的关系进行分析,但已有研究表明,qLTG3-1中存在丰富的序列变异[39-40],其他变异位点也可能是功能位点。特别是华南籼稻中qLTG3-1的等位基因类型并不明确,在华南籼稻中是否存在功能位点,这些问题值得进一步研究。选用遗传背景一致、供体来源多样的单片段代换系来进行qLTG3-1的等位基因效应分析将获得更客观准确的结果。
4 结论
qLTG3-1是1 个控制水稻低温发芽力的重要QTL,其序列变异丰富,存在多种类型的等位基因,+127 位的18 bp 缺失变异在水稻种质中广泛存在。分子标记Gltg3-1 能特异区分这18 bp 变异。基于Gltg3-1 的扩增带型对品种/系分组进行低温发芽力差异性检测,发现两组的低温发芽力无显著差异,即qLTG3-1上的18 bp 变异并不能标示水稻低温发芽力的强弱。因此,需要对qLTG3-1等位基因的效应进行评价,获得增强水稻低温发芽力的等位基因用于品种改良。来源于南洋占的qLTG3-1能显著提高低温发芽力,该等位基因可用于水稻低温发芽力的改良。分子标记Gltg3-1存在于基因内部,可以准确地在杂交后代中选择到目标基因。
猜你喜欢
安徽农学通报(2022年8期)2022-05-06
华人时刊(2021年13期)2021-11-27
智慧健康(2021年17期)2021-07-30
东坡赤壁诗词(2018年5期)2018-12-17
作文中学版(2018年11期)2018-11-29
新课程·下旬(2018年9期)2018-11-14
意林·少年版(2018年11期)2018-07-04
作文新天地(初中版)(2015年10期)2015-10-23
青少年科技博览(中学版)(2015年10期)2015-01-11
作文周刊·小学六年级版(2014年30期)2014-09-15
