
水稻耐热相关功能基因的克隆及其分子机理研究进展
2021-11-12 11:34丁杰荣孙炳蕊王庆林范芝兰潘大建陈文丰
广东农业科学 2021年10期
丁杰荣,孙炳蕊,王庆林,范芝兰,潘大建,陈文丰,李 晨,刘 清
(1.广东省农业科学院水稻研究所/广东省水稻育种新技术重点实验室/广东省水稻工程实验室,广东 广州 510640;2.广州国家现代农业产业科技创新中心,广东 广州 510520)
水稻起源于低纬度的热带地区,形成了适应高温和短日照生态环境的生长习性。但水稻生长发育受温度影响较大,以孕穗至抽穗扬花期对温度最敏感,此阶段水稻的最适温度为25~30℃,如遇日均温度高于32℃或日最高温度高于35℃,水稻会出现花器官发育不全、花粉发育不良且活力下降,同时开花散粉和花粉管伸长也会受阻,导致大量空秕粒形成,从而造成严重的产量损失和品质下降[1-6]。
随着全球工业化进程的加速,全球温室效应日益加剧。自1880—2012 年,全球气温上升了0.85 ℃,预计到21 世纪末将增加1.5~4.5 ℃,极端高温出现频率将大幅提高[7]。Peng 等[8]20 多年跟踪研究发现,全球气温每升高1 ℃,水稻产量将下降10%。异常高温已对我国的水稻生产造成了严重影响。2010 年和2013 年,我国 长江中下游地区发生了大范围的持续高温,2013 年江苏省许多地区甚至出现长达45 d 的持续高温天气,导致该地区水稻严重减产[9-10]。高温热害天气越来越频繁,已成为水稻生产的主要灾害性气候之一[6,11-12],解决水稻热害问题刻不容缓。
目前,培育耐热水稻品种被认为是解决热害问题最经济、最有效的途径。然而,利用常规方法开展水稻耐热性育种非常困难,一是水稻热害研究的条件(尤其是自然条件下的环境温度)难以控制;二是水稻的耐热性是由多个基因控制的数量性状,遗传机理较为复杂[13]。深入了解水稻耐热的分子机理,开展高效准确的分子育种,水稻耐热问题将有望解决。近年来,随着测序技术的迅猛发展和功能基因组学研究的不断进步,水稻耐热性的相关研究取得了较大进展,定位了一些与水稻耐热性相关的QTL,部分相关功能基因被成功克隆。此外,水稻耐热分子机理的研究也取得了一定进展。本文就水稻耐热性相关QTL的鉴定、功能基因的克隆及分子机理研究等方面进行综述,以期为水稻耐热分子育种的开展提供理论参考。
1 水稻耐热相关QTL 的鉴定
近年来,随着分子标记技术的发展以及对水稻热害的高度重视,水稻耐热性分子机理的研究已取得了一定进展。国内外迄今已定位80 个以上水稻耐热QTL,这些QTL 分布于12 条染色体上[13-27]。
1.1 耐热QTL 的定位
水稻在孕穗和抽穗灌浆期对高温比较敏感,目前定位的耐热QTL 大部分是孕穗、抽穗开花和灌浆结实期相关的QTL,只有少数是苗期相关的QTL。耐热和热敏感亲本杂交后代的重组自交系是定位耐热QTL 的重要材料。陈庆全等[14]利用T219 和T226 构建的重组自交系,两年间共定位了6 个抽穗开花期耐热性主效应QTL,分别位于第2、3、8、9、12 号染色体上,qHt2、qHt3、qHt8和qHt12来 自T226,qHt9a和qHt9b来 自T219,其中qHt3在两年实验中都被定位到,有潜在的分子育种应用价值[14]。Jagadish 等[15]利用Bala 和Azucena 构建的重组自交系定位了分布在第1、2、3、8、10、11 号染色体上的8 个耐热QTL,其中1 个QTL 对表型贡献率高达18%。Tazib 等[16]利用 Nipponbare 和Kasalath 构建的回交重组自交系,以花药长度等作为性状指标,共定位了12 个耐热QTL。盘毅等[17]利用996 和4628 构建的重组自交系,定位了2 个花粉育性相关的耐热 QTL,其中qPF4可使水稻在高温下花粉可育率提高7.15%,qPF6同样能提高水稻在高温下的花粉可育率。Ps 等[18]利用N22 和IR64 构建的重组自交系也定位了5 个耐热QTL,分别位于第3、5、9、12 染色体上,其中一个主效QTL与前期定位的QTL 重叠(qSTIY5.1/qSSIY5.2),另一个主效QTL(qSTIPSS9.1)为新定位的耐热QTL。
除重组自交系外,研究人员构建的近等基因系、染色体片段代换系和渗入系也是定位水稻耐热QTL 的重要材料。曹志斌等[19]利用元江普通野生稻荷花塘3 号为供体、籼稻恢复系蜀恢527为轮回亲本构建的种间近等基因系成功定位到qHTH5,其在F2和F3代的表型贡献率分别为8.6%和19.4%。Li 等[20]利用Liaoyan241 和IAPAR-9构建的近等基因系,以水稻结实率为性状指标,定位了11 个耐热QTL,其中qNS1、qNS4、qNS6和qRRS4等4 个主效QTL 能稳定地在不同年份或环境中被定位到。张昌全等[21]利用以日本晴和9311 构建的染色体片段代换系定位了3 个耐热性QTL。奎丽梅等[22]利用遗传背景为特青的云南元江野生稻渗入系材料,在第1、3、8、10 号染色体上各定位了1 个抽穗开花期耐热相关QTL。
上述是孕穗、抽穗开花和灌浆结实期的水稻耐热QTL,而有关苗期耐热QTL 报道较少。Kilasi 等[23]利用N22 和IR64 构建的近等基因系对水稻幼苗进行热害处理,以根长和株高为性状指标,共定位到10 个耐热QTL,其中rlht5.1、slht6.1/slpc6.1、slpc2.1、slpc10.2和slpc10.3的 表型贡献率都超过10%。而Lei 等[24]利用与奎丽梅相同的材料[22],也定位到5 个苗期耐热性相关QTL,其中位于3 号染色体的耐热性QTL 与奎丽梅等[22]定位的qHT3位置相近,说明苗期耐热性与抽穗开花期耐热性具有一定的相关性。
1.2 耐热QTL 间的互作
水稻耐热性是由多基因控制的数量性状,单个耐热QTL 起作用的同时,不同耐热QTL间的互作也对水稻耐热性起重要作用。赵志刚等[25]定位的7 个耐热QTL 分布在4 条染色体上,它们的表型贡献率为6.4%~19.7%,加性效应为-21.3%~20.9%。曹立勇等[26]利用IR64 和Azucena 花药培养的DH 群体定位到6 个开花结实相关的具有加性效应的耐热QTL,这6 个QTL能提高结实率4.33%~10.37%;此外,在第 1、2、3、4、5、7、8、11 等8 条染色体间还检测到 8 对加性×加性上位性效应,其贡献率为2.27%~8.13%。陈庆全等[14]在定位水稻耐热QTL时也检测到7 对上位性QTL,位于第1、2、4、11、12 染色体上的4 对QTL 间,以及位于第2、3、4、7、8、9 染色体上的3 对QTL 间分别存在互作效应。朱昌兰等[27]定位到3 个灌浆期耐热性主效QTL,并检测到8 对上位性互作QTL 位点,它们对粒重变化的贡献率为2.45%~5.29%。不同耐热QTL 表型贡献率差异较大,主效QTL 的定位及对应基因的克隆对水稻耐热品种的选育意义重大。但不同QTL 之间错综复杂的互作使水稻耐热QTL 的定位困难重重,水稻耐热基因的克隆难度很大。
2 水稻耐热相关功能基因的克隆
水稻耐热性是由多基因控制的复杂性状,且难以准确进行表型鉴定,因此水稻耐热功能基因的克隆鉴定显得尤为困难。目前,仅有不到30 个水稻耐热相关基因被克隆鉴定,其中多数基因是通过反向遗传学方式克隆鉴定得到,仅有少数几个基因由正向遗传学手段克隆鉴定得到。克隆鉴定所得的水稻耐热相关基因多数编码蛋白质,其中11 个基因编码蛋白酶类,10 个基因编码热激蛋白等其他蛋白质,3 个基因是转录因子(表1)。
2.1 通过正向遗传学方式克隆功能基因
通过正向遗传学手段克隆的水稻耐热基因对水稻耐热性均起正向调控作用。Li 等[28]以生长于热带的非洲稻为材料,通过与亚洲栽培稻杂交构建遗传群体,并进一步进行遗传分析和定位克隆,成功分离克隆了控制非洲稻的一个耐高温主效QTL 的功能基因OgTT1(Oryza glaberrimaThermo-tolerance 1)。OgTT1编码一个26S 蛋白酶体的α2 亚基,在泛素化介导的蛋白降解通路中发挥重要作用。过表达OgTT1能够显著增强水稻在苗期、开花期和灌浆期的耐热性[28]。Wei 等[30]以耐热水稻品系HT54(源自广陆矮4号)与热敏感品系HT13(源自籼稻05-占)构建遗传群体,定位了调控水稻苗期耐热的主效位点OsHTAS(Oryza sativaHeat Tolerance at Seeding Stage)。进一步研究表明,OsHTAS编码一个泛素连接酶,过表达OsHTAS能显著增强水稻的苗期耐热性[23]。Wang 等[31]从籼稻品种中籼3037 自然突变体中克隆到耐热功能基因TOGR1(Thermotolerant Growth Required 1),该基因编码一个DEAD-Box RNA 解旋酶,其表达同时受到温度和昼夜节律的调控,且与日常温度的波动紧密相关,温度升高会直接增强其解旋酶活性,TOGR1转录水平的变化还与株高呈正相关,过表达TOGR1会使水稻在高温条件下生长更好。此外,Shen 等[32]从拟南芥中分离得到了1 个富含亮氨酸重复序列的类受体激酶基因ER(Receptor-like Kinase ERECTA),在水稻中过表达ER基因能显著增强水稻的耐热性。“高ER水稻”与野生型植株相比,在高温条件下的结实率显著增加;反之,在水稻中功能性突变ER的同源基因会显著降低植株的耐热性[32]。
2.2 通过反向遗传学方式克隆功能基因
目前通过反向遗传学手段克隆鉴定的水稻耐热基因有20 多个,其中多数基因起正向调控作用,仅有几个基因起负向调控作用(表1)。
2.2.1发挥正向调控作用的功能基因 热激蛋白(Heat Shock Protein,HSP)是植物热信号传导通路中的重要组成部分。在酵母细胞中过表达水稻OsHSP90基因能显著增强细胞在42、50、70 ℃下的耐热能力[33]。此外,Lin 等[34]研究发现,HSP101 通过与热相关蛋白HSA32(Heat Stressassociated 32-kd Protein)的互作,在水稻苗期的基础耐热性和后天长期的耐热性驯化中发挥了重要的调控作用。OsANN1是水稻膜联蛋白家族中的一个成员,其表达受高温处理显著诱导[35]。过表达OsANN1能显著改善水稻在高温条件下的生长,而OsANN1的基因敲除株对高温热害更加敏感[29]。OsHCI1(Oryza sativaHeat and Cold Induced 1)编码一个环finger E3 连接酶,也受高温诱导[36]。在正常生长条件下,该基因定位于高尔基体中,且能够沿着细胞骨架快速移动,但是在受到高温处理后,OsHCI1可能累积在细胞核中[36]。在拟南芥中过表达OsHCI1能显著增加植株在高温下的存活率[36]。与OsHCI1基因相似,OsHIRP1(Oryza sativaHeat-induced RING Finger Protein 1)也编码一个环finger E3 连接酶,经45 ℃高温处理后,OsHIRP1主要在细胞核中富集,在拟南芥中过表达OsHIRP1能显著提高种子的发芽率[37]。此外,OsMYB55、SNAC3和OsNTL3基因的表达也受到高温热害的显著诱导。过表达OsMYB55显著改善水稻在高温条件下的生长,同时减小高温对产量的影响[38]。过表达SNAC3和OsNTL3的水稻植株耐热性显著增强,反之,SNAC3的基因沉默植株和OsNTL3的基因编辑植株对热害更加敏感[39-40]。
SUMO(Small Ubiquitin-related Modifier)化修饰是一个类泛素化的过程,在高等植物的生命活动中发挥重要作用。在匍匐翦股颖(Agrostis stoloniferaL.)中过表达编码SUMO E3 连接酶的OsSIZ1基因会使植株在高温条件下的耐热性显著增强[41]。转录后RNA 修饰也广泛存在于活体生物中,其中一种丰富的mRNA 转录后修饰方式就是胞嘧啶的5'端甲基化(m5C)修饰。编码m5C甲基转移酶的OsNSUN2基因在水稻耐热性中也发挥了重要调控作用,osnsun2突变植株的苗期耐热性显著降低[42]。Shiraya 等[43]研究表明,高尔基体/质体类型的锰超氧化物歧化酶(Manganese SOD 1,MSD1)在耐热水稻品种中的表达量显著高于耐热性较低的水稻品种,持续高表达MSD1的植株在高温条件下的籽粒品质显著优于野生型植株的籽粒品质,反之,msd1敲除的水稻植株对热胁迫的反应更加敏感。此外,过表达编码水稻异三聚体G 蛋白β 亚基的OsRGB1显著增强转基因植株的发芽期和苗期耐热性[44]。过表达编码小GTP 结合蛋白的OsRab7也使转基因水稻植株的苗期耐热性显著增强[45]。此外,已有研究表明钙信号在植物响应非生物胁迫过程中也发挥重要作用。Cui 等[46]研究发现,OsCNGC14(Oryza sativaCyclic Nucleotide-gated Ion Channel 14)和OsCNGC16(Oryza sativaCyclic Nucleotide-gated Ion Channel 16)这两个环核苷酸门控离子通道蛋白正向调控水稻植株的耐热性。oscngc14/16双突变植株的苗期耐热性显著降低,且突变这两个蛋白的任何一个都可降低或者消除热胁迫诱导的胞质钙信号[46]。
2.2.2发挥负向调控的功能基因OsHSBP1(Oryza sativaHeat Shock Factor Binding Protein 1)、OsHSBP2(Oryza sativaHeat Shock Factor Binding Protein 2)、OsMDHAR4(Oryza sativaMonodehydroascorbate Reductase 4)、OsFBN1(Oryza sativaFibrillin 1)和OsUBP21(Oryza sativaUbiquitin-Specific Protease 21)在水稻耐热性中起负向调控作用。OsHSBP1和OsHSBP2编码HSF(Heat Shock Factor)结合蛋白,正常生长条件下它们广泛表达于各个组织,且在高温处理后的恢复期表达水平急剧上调[47]。过表达OsHSBP1或OsHSBP2的水稻植株对高温热害更加敏感,而OsHSBP1或OsHSBP2的基因沉默株系在高温条件下幼苗的存活率显著增加[47]。过表达编码一个单脱水抗坏血酸还原酶的OsMDHAR4会显著降低水稻植株在苗期的耐热性,而osmdhar4突变体植株的苗期耐热性显著增强[48]。此外,过表达编码质体-脂质相关蛋白的OsFBN1显著降低水稻植株在苗期以及生殖生长期的耐热性[49],而基因敲除或者降低编码泛素特异性蛋白酶的OsUBP21的表达,可明显增强转基因植株的苗期耐热性[50]。
3 水稻耐热分子机制研究
目前水稻耐热基因介导的耐热分子机制研究主要集中在以下方面:维持植物体内蛋白及rRNA的正常水平及稳定性、维持细胞膜的完整性和维持细胞内正常的活性氧(Reactive Oxygen Species,ROS)水平(图1)[27-28,31-32,35,39,41,43,45]。
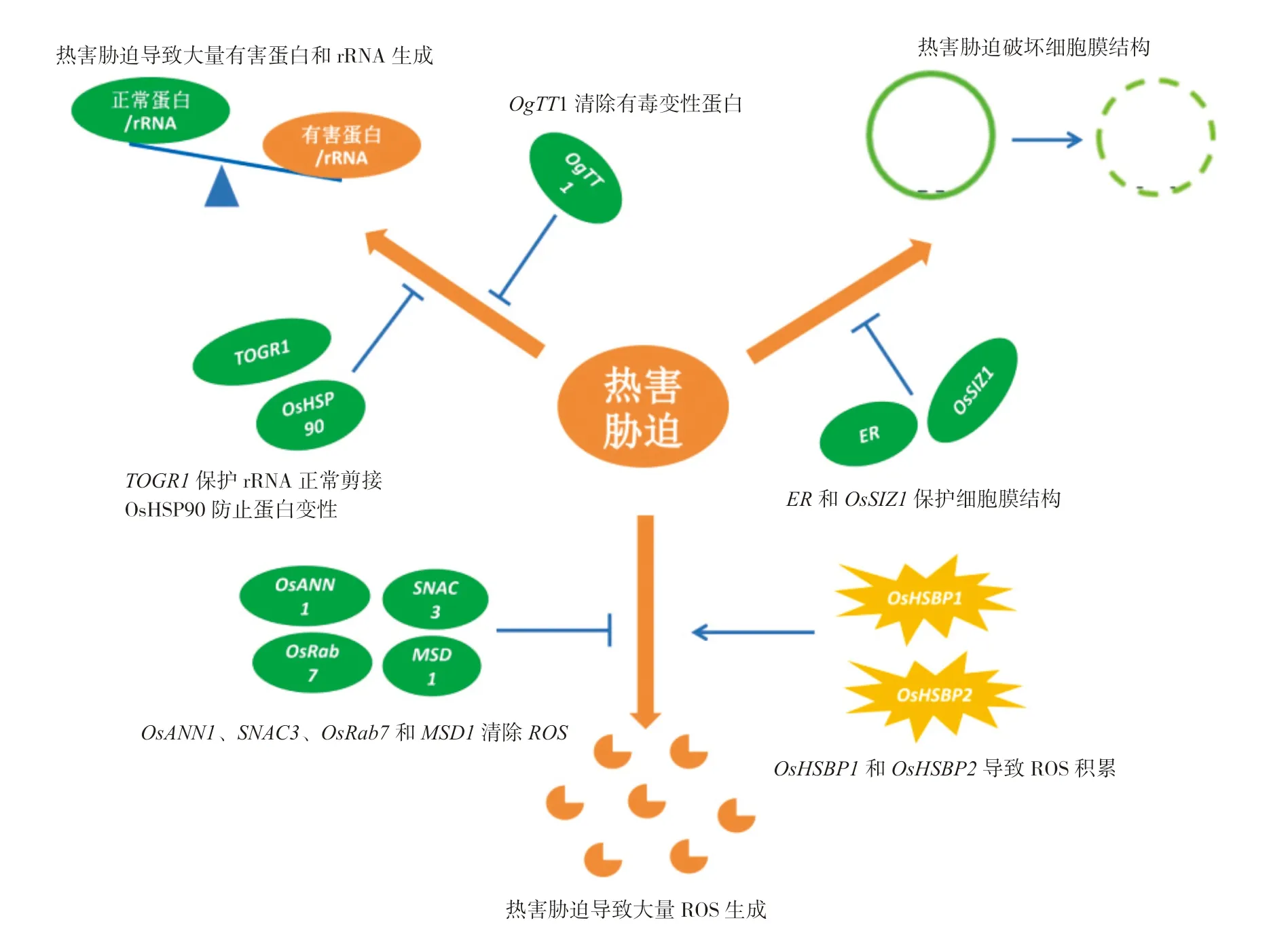
图1 水稻耐热相关基因介导的分子机制Fig.1 Molecular mechanism mediated by heat tolerance related genes in rice
3.1 维持植物体内蛋白及rRNA 的正常水平及稳定性
植物细胞在高温逆境下积累大量的变性蛋白,这些错误折叠的蛋白会对细胞产生毒性[51]。OsHSP90介导的耐热性与在高温条件下预防蛋白变性密切相关,而且温度越高,这种预防蛋白变性能力越强[27]。与热激蛋白预防蛋白变性不同,对OgTT1控制水稻高温抗性的机理研究表明,OgTT1在转录水平上对高温的响应更有效,而且其编码的蛋白使细胞中的蛋白酶体在高温下对泛素化底物的降解速率更快,促使水稻细胞中积累的有毒变性蛋白的种类和数量均显著降低,进而保护水稻细胞。该研究也揭示了植物细胞响应高温的新机制,即及时有效清除变性蛋白,对维持高温下胞内蛋白平衡至关重要;尤其在极端高温时,清除变性蛋白比复性变性蛋白的效率更高[28]。除了高温条件下维持蛋白的平衡外,水稻的耐热性与rRNA 稳定性也密切相关。TOGR1作用于小亚基(SSU)rRNA 前体的加工,即使在高温热害的条件下也能保证rRNA 前体的正常剪切,因而能保持植物体内正常的rRNA 水平[31]。
3.2 维持细胞膜的完整性
非生物胁迫条件下,细胞膜的完整性与植物耐热性密切相关。对ER基因介导的耐热性机理研究表明,ER基因表达水平提高后,植物细胞膜在逆境下更完整,细胞不易死亡。细胞增多变大,组织器官与生物量也随之变大,使植株总体壮实得以增产[32]。高温胁迫下,过表达OsSIZ1的转基因匍匐翦股颖植株比无OsSIZ1的对照植物表现出更完整的细胞膜、更强的保水性,所受的热害也更轻微[41]。在番茄中的研究也发现,SlSIZ1的干扰株系比其野生型株系对热更敏感,SlSIZ1的过表达株系则更耐热[52]。进一步研究发现,热胁迫下SlSIZ1过表达株系中的HSP70、HSP90和HsfA2的表达量显著增加,而SlSIZ1 可与SlHsfA1 互作并介导SlHsfA1 的SUMO 化[52]。由此可见,SlSIZ1可能是通过调控HSP70和HsfA2等下游基因的表达对某些蛋白起保护作用或介导其SUMO 化以保护细胞膜结构。
3.3 维持细胞内正常的ROS 水平
ROS 的积累影响耐热性,而植物细胞在遭受高温热害后,体内会富集过多的ROS,从而危害细胞内的膜脂、蛋白和核酸等生物分子,造成植物细胞死亡[53]。已有研究表明,OsANN1、SNAC3、OsRab7和MSD1介导的耐热性是通过降低热害产生的ROS 水平来实现的[35,39,43,45]。过表达SNAC3基因,显著降低了植株体内的H2O2水平,表现出更好的耐热性[39]。与SNAC3基因相反,OsHSBP1或OsHSBP2介导的热敏性是由体内过多的ROS 导致的[47]。OsNSUN2突变导致的耐热性降低也与植株体内积累了过多的ROS 密切相关[41]。
4 展望
目前尽管水稻育种学家已鉴定出众多水稻耐热相关QTL,但是水稻耐热功能基因的分离克隆还较少,应用分子标记辅助选择培育耐热水稻品种也暂未见成功报道[28-29]。这主要有以下原因:一是水稻耐热性为多基因控制的数量性状,且一般单个QTL 的表型贡献率较低,不同QTL 之间经常会有互作;二是水稻耐热性的表型鉴定较为困难,难以控制田间实验处理的环境和温度;三是水稻耐热性相关基因表达受环境影响较大,有些基因只在特定时期才表达发挥功能,因此难以准确鉴定。
目前已克隆鉴定的水稻耐热相关功能基因还较少,所开展的研究也主要集中在水稻苗期进行,目前对苗期外的水稻耐热性相关功能基因克隆鉴定仅有个别报道。但是水稻从孕穗期开始,对热害的敏感度明显高于苗期,遇热害时产量受损更严重,而且不同生育期可能有不同的耐热分子机制。因此,克隆鉴定水稻生殖生长期耐热性相关的功能基因将是水稻耐热性研究的侧重点。克隆鉴定目标功能基因的同时,还需要深入研究它们所介导的水稻耐热性相关分子机制,以期为培育耐热且高产优质的水稻品种奠定理论基础。
猜你喜欢
作物学报(2022年12期)2022-10-14
环球时报(2022-09-20)2022-09-20
今日农业(2021年9期)2021-11-26
今日农业(2021年9期)2021-11-26
当代水产(2021年6期)2021-08-13
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
安徽农学通报(2015年4期)2015-07-30
时代英语·高二(2015年2期)2015-05-18
