
基于克里金插值的白纹伊蚊空间分布研究*
2021-11-06 06:13周毅彬
寄生虫与医学昆虫学报 2021年2期
周毅彬
(上海市疾病预防控制中心传染病防治所,上海 200336)
在过去的30—40年里,白纹伊蚊Aedesalbopictus已经入侵了除南极洲以外所有大陆(Kotsakiozietal.,2017)。它是一种主要的人类叮咬害虫,可显著降低被叮咬者的生活质量,也是登革热、基孔肯雅热、黄热病和寨卡病毒病等主要虫媒疾病的传播媒介。在2004(Xuetal.,2007)、2009(凌锋等,2010)和2017年(Yanetal.,2018)毗邻上海市的浙江省发生多起本地登革热疫情,其主要传播媒介也是白纹伊蚊。2017年上海报告的登革热病例是上海近五十年来首例本地登革热病例(上海年鉴,2018),2018年上海市又报告了3起本地案例(须俊明等,2019)。预防和控制登革热最有效的方法依旧是媒介蚊虫密度控制和监测,因此白纹伊蚊的密度监测显得尤为重要。空间分析是基于地理信息系统(Geographic information system,GIS)的一门学科。其中克里金法(Kriging)是依据协方差函数对随机过程/随机场进行空间建模和预测(插值)的回归算法,广泛应用于蚊虫的研究(Aziletal., 2014)。
目前的白纹伊蚊监测方法中,如诱蚊灯法、人诱停落法法和BG诱捕器,通常都是劳动密集型和监测成本昂贵且难以大规模实施(Manicaetal.,2017)。与这些方法相比,诱卵器的实施相对简单和便宜,并且它是便携式的,不需要电力或诱饵。这种方法即使在较低的种群密度下也能监测到伊蚊的存在(Silver, 2008),因此被广泛用作监测小容器中产卵蚊虫空间活动的方法(Service, 1993)。
本研究在约60 000 m2研究区域内设置132个诱蚊诱卵器监测点,2020年7月28日至10月6日,每周1次布放诱蚊诱卵器,旨在使用克里金(Kriging)模型(Matheron,1963)研究预测非监测点白纹伊蚊的密度,并确定其热点和危险区。
1 材料与方法
1.1 研究区域
本次研究选取位于上海市徐汇区的某处科教园区,研究区域东西向约250 m、南北向约240 m,总面积约60 000 m2。内有11幢大小不一的建筑,园区内绿化以乔木和草地为主(图1)。在研究区域内共设置132个诱蚊诱卵器监测点,各监测点相距10 m以上,选择避阳绿化地带放置,并对每个监测点进行定位(图1)。
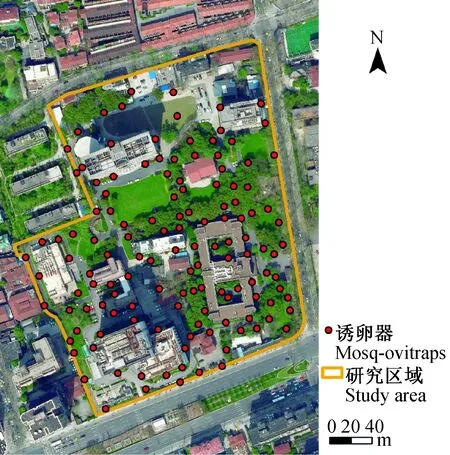
图1 研究区域与诱蚊诱卵器监测点位置Fig.1 Study area and monitoring points of mosq-ovitraps
1.2 白纹伊蚊密度监测方法
本次研究使用《GB/T 23797-2009病媒生物密度监测方法 蚊虫》标准的诱蚊诱卵器,直径70 mm、高100 mm的圆柱形透明塑料瓶,瓶底向上突出高20 mm的椭圆圆锥形,瓶盖为黑色塑料,上有3个向内突出21 mm的倒圆锥管,上口直径12 mm,下口直径7 mm。使用时在瓶内倒入约20 mL脱氯水,并放入一张70 mm直径的滤纸,滤纸浸湿并浮于水的上方。在每个监测点布放1只诱蚊诱卵器,连续放置7 d,于第8 d回收并观察记录结果。
1.3 研究时间与频次
研究时间为2020年7月28日至10月6日(上海市一年中蚊虫密度较高的季节),每周一布放诱蚊诱卵器,监测时间持续7 d,于下一周的周一回收并放置新的诱蚊诱卵器。
1.4 诱蚊诱卵器阳性判断标准及计算方法
诱蚊诱卵器内有成蚊或(和)蚊虫卵的,判定为阳性;既无成蚊也无蚊虫卵的,判定为阴性;丢失、干涸或破损的诱蚊诱卵器,判定为无效。对阳性诱蚊诱卵器内的蚊虫卵进行计数。诱蚊诱卵器指数=阳性诱蚊诱卵器个数/回收诱蚊诱卵器总数×100。
1.5 统计学分析
1.5.1空间分析:使用ArcGIS10.8软件(ESRI China)进行空间分析。通过计算增量空间自相关(多个距离的全局Moran′sI)来评估白纹伊蚊卵粒数量是否存在空间自相关。全局Moran′sI(Moran,1950)使用基于特征位置和属性值(每个位置诱蚊诱卵器捕获的总卵粒数)的置换过程,对照空假设(没有空间自相关)进行检验。全局 Moran′sI可以识别整个研究区域的空间格局,但是无法确定某一位置的空间格局。局部 Moran′sI(Local Moran′sI)分析用以评估每一监测位置是否存在具有统计意义的聚集、冷点和空间异常值的点(Anselin L,1995)。
1.5.2克里金插值:使用ArcGIS 10.8软件地统计学分析扩展模块的经验贝叶斯克里金(Empirical Bayesian Kriging)模型(Gribovetal.,2020)进行。克里金插值方法用于量化数据的空间结构并预测未采样位置的丰度。包括探索性统计分析、变异函数建模。输出是预测每个位置预测值的面和预测标准误差的面。使用交叉验证方法来确定克里金插值是否在未采样位置提供可靠的估计。交叉验证中用于准确预测的标准如下:标准化均方根(RMSS)约为1,标准化平均值(MS)约为0,均方根(RMS)约为平均标准误差(ASE)。
2 结果
2.1 诱蚊诱卵器结果
本次研究期间,共回收有效诱蚊诱卵器1 308 只,阳性424只,诱蚊诱卵器指数为32.42,阳性诱蚊诱卵器中卵粒数的中位数为23粒,最少为0粒(只捕获到成蚊)、最多为253粒;阳性诱蚊诱卵器捕获成蚊中位数为1只,最少为0只(只有卵粒)、最多为6只成蚊。
2.2 诱蚊诱卵器指数时间变化
2020年7月28日至10月6日实验期间,研究区域的诱蚊诱卵器指数变化见图2。7月28日放置的诱蚊诱卵器指数为26.36;其后逐渐上升,8月18日达最大值53.08;9月29日下降至17.42。
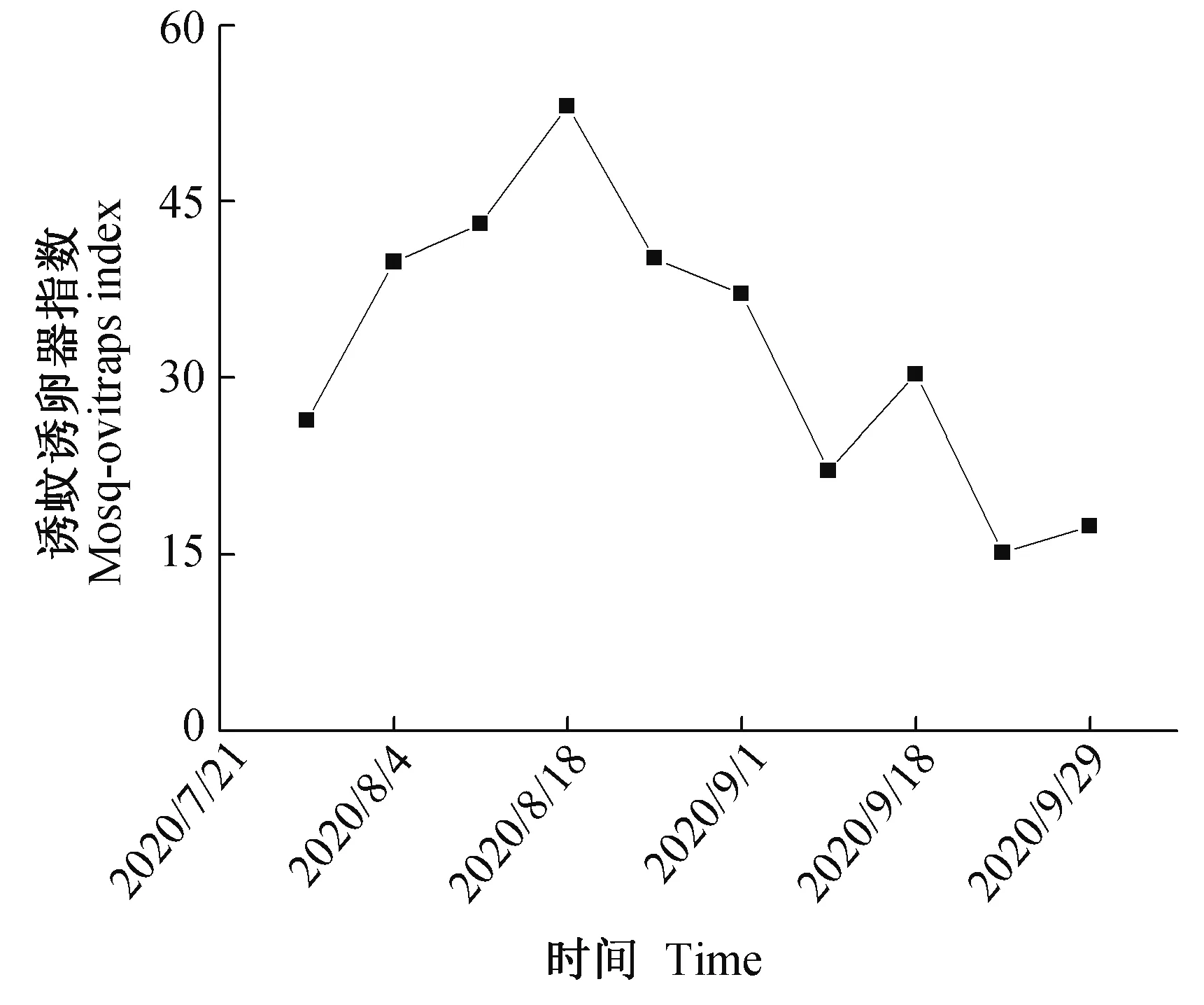
图2 实验期间诱蚊诱卵器指数变化Fig.2 Mosq-ovitraps index in study area during the experimental period
2.3 诱蚊诱卵器捕获成蚊与卵粒
2020年7月28日至10月6日实验期间,共放置了10次诱蚊诱卵器,每次放置132个。每个诱蚊诱卵器10次共计捕获的白纹伊蚊成蚊和卵粒数的散点图见图3。捕获的成蚊和卵粒数的散点呈线性分布,两者具有相关性,相关系数为0.8793(P<0.05)。线性回归结果:y=-0.638+22.953x(x为成蚊数,y为卵粒数)。由于监测的卵粒数的数据,相较于阳性率能获得更多信息,其次诱蚊诱卵器监测时成蚊有一定的逃逸率,因此选用卵粒数作为空间分析数据。
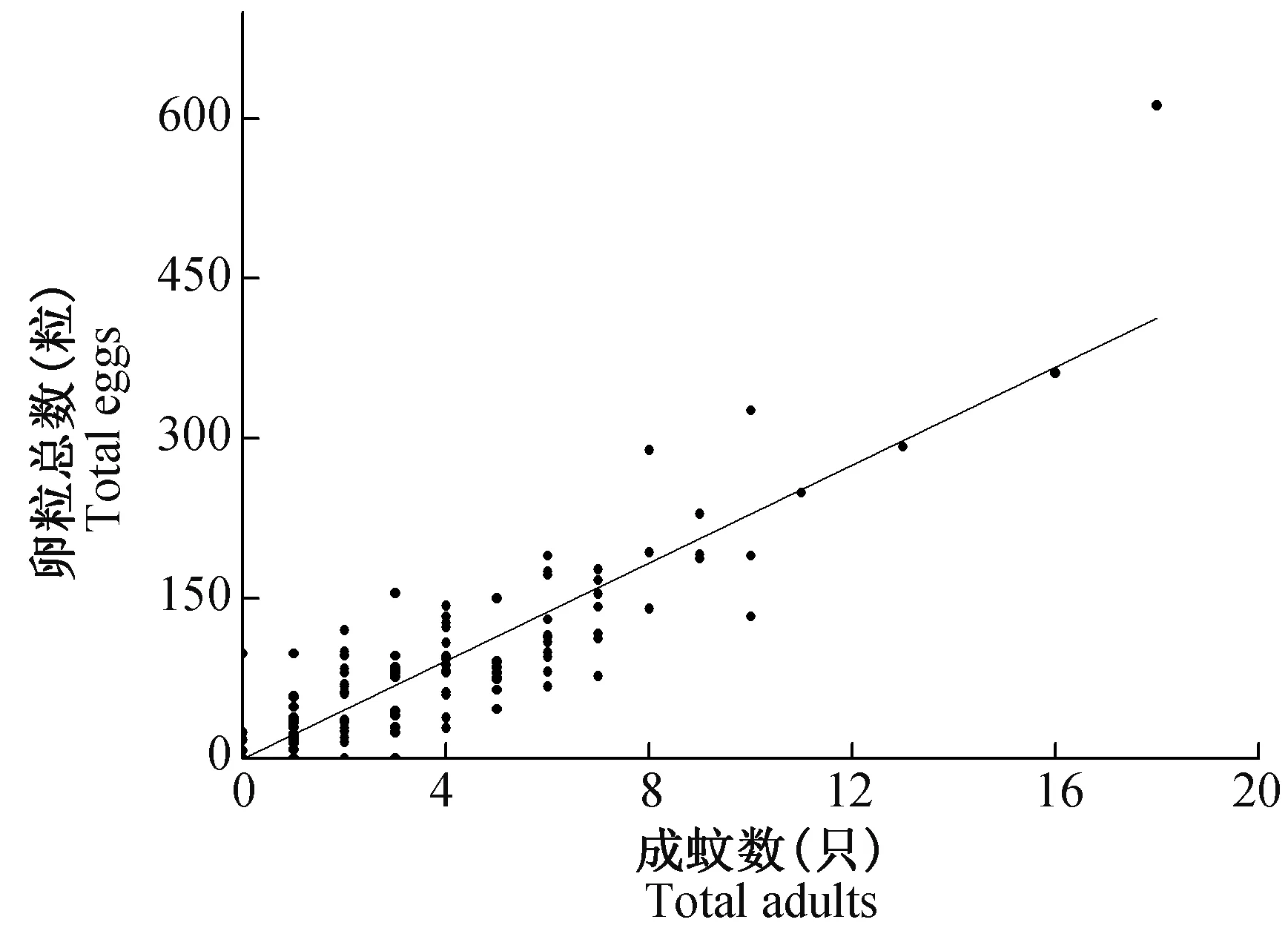
图3 实验期间每个位置诱蚊诱卵器捕获总卵粒与成蚊散点图Fig.3 Scatter plot of total adults and eggs in each mosq-ovitrap in study area during the experiment
2.4 空间自相关分析
本次研究期间,每个监测点10次监测合计捕获的卵粒数增量空间自相关分析显示,全局空间自相关峰值半径(Z值最大)为62 m,Moran′sI指数为0.067,Z值为2.995(P<0.05),即卵粒数的空间自相关在半径62 m达最大值。局部自相关分析结果见图4,研究区域呈现出西南角蚊密度较低,但其中有3个高密度的异常点;东侧蚊密度较高,但其中也有数个低密度的异常点。
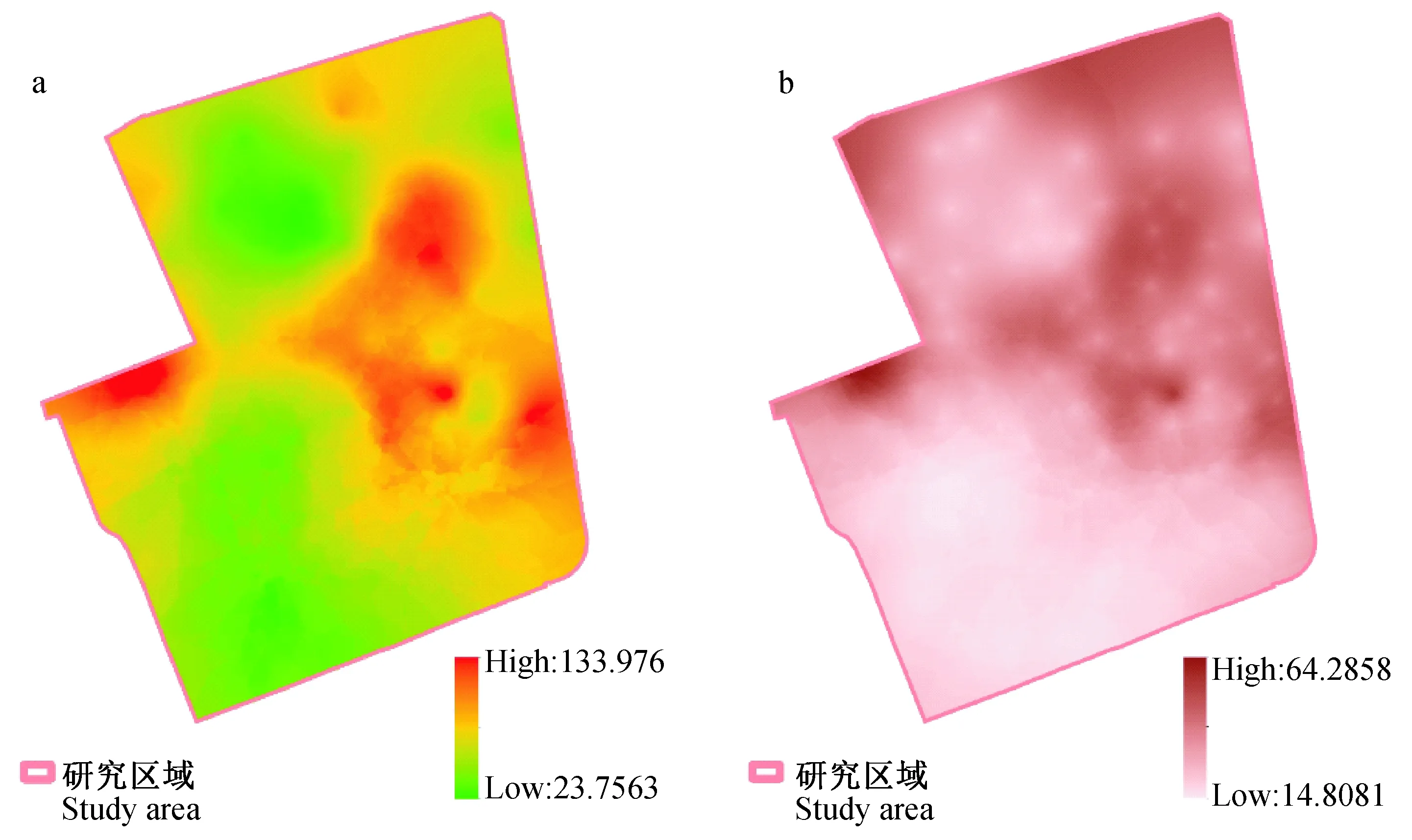
图5 经验贝叶斯克里金插值(a)及误差面(b)Fig.5 Prediction surface(a) and standard errors (b) of empirical Bayesian kriging
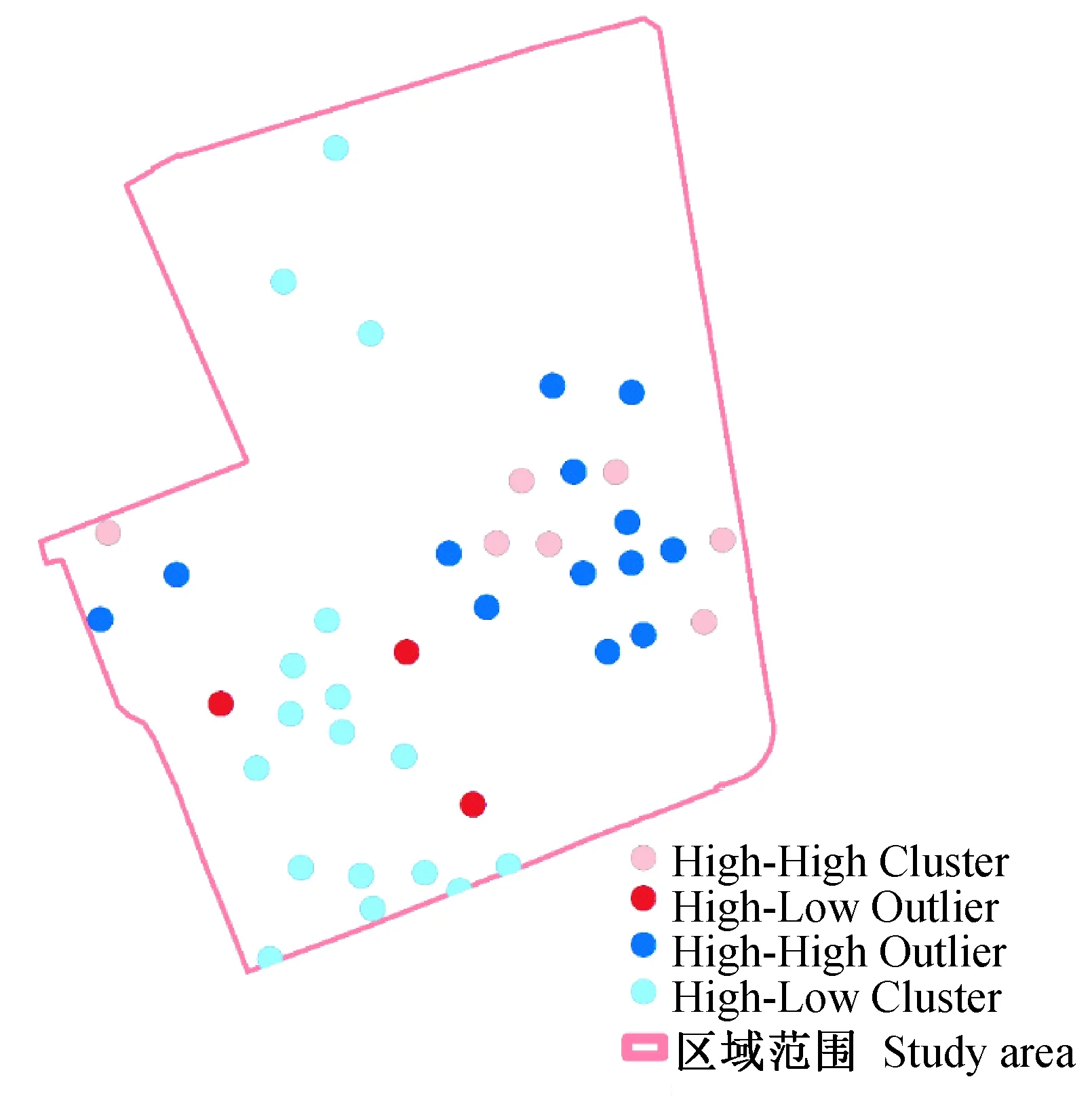
图4 实验期间每个位置诱蚊诱卵器捕获总卵粒局部自相关分析Fig.4 LISA of total eggs′numbers in each mosq-ovitrap in study area during the experimental period
2.5 经验贝叶斯克里金模型
以本次研究期间每个监测点10次监测合计捕获的卵粒数为数据,进行经验贝叶斯克里金的空间插值分析。模型参数:数据变换选择经验法,半变异函数选择指数模型。经验贝叶斯克里金插值结果见图5-a,插值误差见图5-b。交叉验证结果:标准化均方根(RMSS)为1.017,标准化平均值(MS)为0.027,均方根(RMS)为81.571,平均标准误差(ASE)为82.617。
3 讨论
地理统计学(Geostatisitics)是以区域化变量为基础,借助变异函数研究具有空间相关性和依赖性的地理现象,对样本数据进行最优的无偏内插估计,模拟地理现象空间分布的相关性和变异性的统计分析法(李俊晓等,2013)。最初,在空间统计中,地统计等同于克里金法,当前定义已经扩展到不仅包括克里金方法,还包括许多其他插值方法,如空间插值的确定性方法等。克里金插值也称为空间局部插值法,是以变异函数理论和结构分析为基础,在有限区域内对区域化变量进行无偏最优估计的一种方法,是地统计学的主要内容之一(Giordano,2018)。由于地统计学的统计特征,运用克里金插值方法进行的插值,不仅可以得到预测结果,而且可以得到预测误差,有利于评估预测结果的不确定性。克里金插值方法主要分为普通克里金法和泛克里金法。普通克里金法是最普通和广泛使用的克里金方法,该方法假定恒定且未知的平均值,泛克里金法假定数据中存在覆盖趋势。经验贝叶斯克里金法则可自动执行构建有效的克里金模型过程中的那些最困难的步骤。它可以用构造子集和模拟的方法自动计算这些参数,而不像其他克里金方法需要手动调整参数。
本次研究期间,每个监测点诱蚊诱卵器捕获的卵粒总数成空间聚集分布(Moran′sI指数为0.067,Z值为2.995,P<0.05)。空间插值结果显示研究区域东侧的中部,以及西北角白纹伊蚊密度较高(图5)。克里金插值方法可以用来补充或代替传统的热力图和热点分析等,图5更好地展示了蚊密度数据的空间分布和可视化结果(Krivoruchko,2011)。从图1可见,东侧中部的白纹伊蚊高密度区域与大片乔木分布范围相吻合。Cianci等(2015)的研究发现乔木与白纹伊蚊卵的分布数量正相关,而草地与卵的数量呈负相关,这与本次结果相一致。本次研究的空间自相关在62 m出现峰值,表明研究区域白纹伊蚊的平均活动范围约为半径60 m (朱伟等,2020),这也说明了研究区域西北角与其北侧的居民区相连,可能是由于居民区较高的蚊密度影响了该处的白纹伊蚊分布。但是本次研究中,临近的诱蚊诱卵器结果也往往并不相同(图4),在东侧的高密度聚集区有数个异常点,这可能是由于诱蚊诱卵器所处的不同环境及其周边环境所造成的。这也是造成本次研究期间空间自相关系数仅0.067的可能原因,提示在研究蚊虫密度分布时候临近环境的因素会起到较大的影响。
本次实验期间,共放置了10次诱蚊诱卵器。每个诱蚊诱卵器监测点共计捕获的白纹伊蚊成蚊和卵粒数具有较好的相关性(图3)。提示在实验期间,诱蚊诱卵器内每只白纹伊蚊成蚊平均产卵22.95个。白纹伊蚊具有跳跃性产卵(skip oviposition)的特点,蚊虫不会在一个地方产卵,而是选择在几个不同的地点产卵(Chadee,2009)。Apostal等(1994) 报道,白纹伊蚊雌蚊会把它们的卵散布到几个容器,每个产卵容器大约有11~30个卵粒,本次观察到的现象与其相一致。
本研究以诱蚊诱卵器捕获卵粒的空间自相关为基础使用经验贝叶斯克里金模型,今后使用协同克里金可以提高空间插值的精度。在丰度空间建模中,未来的研究应包括幼虫孳生地和成虫栖息地等,以及植被情况和人群活动密度等,这将提高模型的准确性和全面性。
克里金模型在蚊虫监测和种群动态空间分布的研究中,能有效识别白纹伊蚊高密度区域,并预测没有监测点处的蚊虫密度,为制定高效的防控策略提供帮助,在今后结合幼虫孳生地和成虫栖息地等参数将具有广阔的应用前景。
猜你喜欢
小学生学习指导(爆笑校园)(2022年9期)2022-09-30
保健医苑(2022年6期)2022-07-08
疯狂英语·读写版(2022年3期)2022-03-31
今日农业(2021年12期)2021-11-28
寄生虫与医学昆虫学报(2021年1期)2021-05-15
化工管理(2021年7期)2021-05-13
寄生虫与医学昆虫学报(2020年4期)2020-03-26
风流一代·经典文摘(2019年12期)2019-09-10
读者(2018年24期)2018-12-04
知识窗(2017年3期)2017-03-09
