
云南南部地区蕨类植物多样性与地理分布
2021-10-08 14:45PhyoKayKhineHaraldSchneider
绿色科技 2021年17期
陈 柯,Phyo Kay Khine,Harald Schneider
(1.中国科学院西双版纳热带植物园综合保护中心,云南 勐腊 666303; 2.中国科学院大学,北京 100049)
1 引言
蕨类植物是地球上最古老的陆生维管植物,也是第二大维管植物类群,全球约分布有蕨类植物12000种[1]。蕨类植物最早起源于4亿多年前的志留纪末期,繁盛于石炭和二叠纪,在白垩纪后逐渐衰退,现存的蕨类植物大多为伴随着被子植物兴起而新演化出的类群[2]。蕨类植物通过孢子繁殖和扩散,不依赖于生物媒介,其多样性分布能更好的为环境变量所解释,是研究生物多样性分布的理想材料[3]。云南南部地区位于全球生物多样性热点的交汇地带,是蕨类植物多样性分布中心,蕨类植物资源尤为丰富。独特的地理环境和地质历史,还使得该地区保存了大量珍稀濒危蕨类植物物种,如二回莲座蕨(Angiopterisbipinnata)、海南实蕨(Bolbitishainanensis)和苏铁蕨(Braineainsignis)等[4,5]。近年来,人口、环境和土地利用等问题的加剧,使得该地区蕨类植物的生存日益受到威胁[6]。本研究利用数字化的蕨类植物标本记录,分析云南南部地区蕨类植物多样性的组成和分布情况,并结合环境数据探讨蕨类植物多样性与气候和地形变量的关系,以期为地区蕨类植物多样性的保护和相关政策的制定提供参考(图1)。
2 数据来源和方法
2.1 研究区域概况
研究区域包括了云南南部的勐海、景洪、勐腊和麻栗坡等6个县市(图1)。该区域与缅甸、老挝以及越南相邻,属于亚洲热带的北缘。气候上受东南和西南季风的影响,年温差小,日温差大,年均温20 ℃;降水集中,干湿分明,年降水1500 mm;地形地貌多样,高山、盆地、河谷、岩溶等地貌交错;植被垂直分布明显,植被类型丰富多样,以季节雨林、湿润雨林和季风常绿阔叶林为主[7~9]。
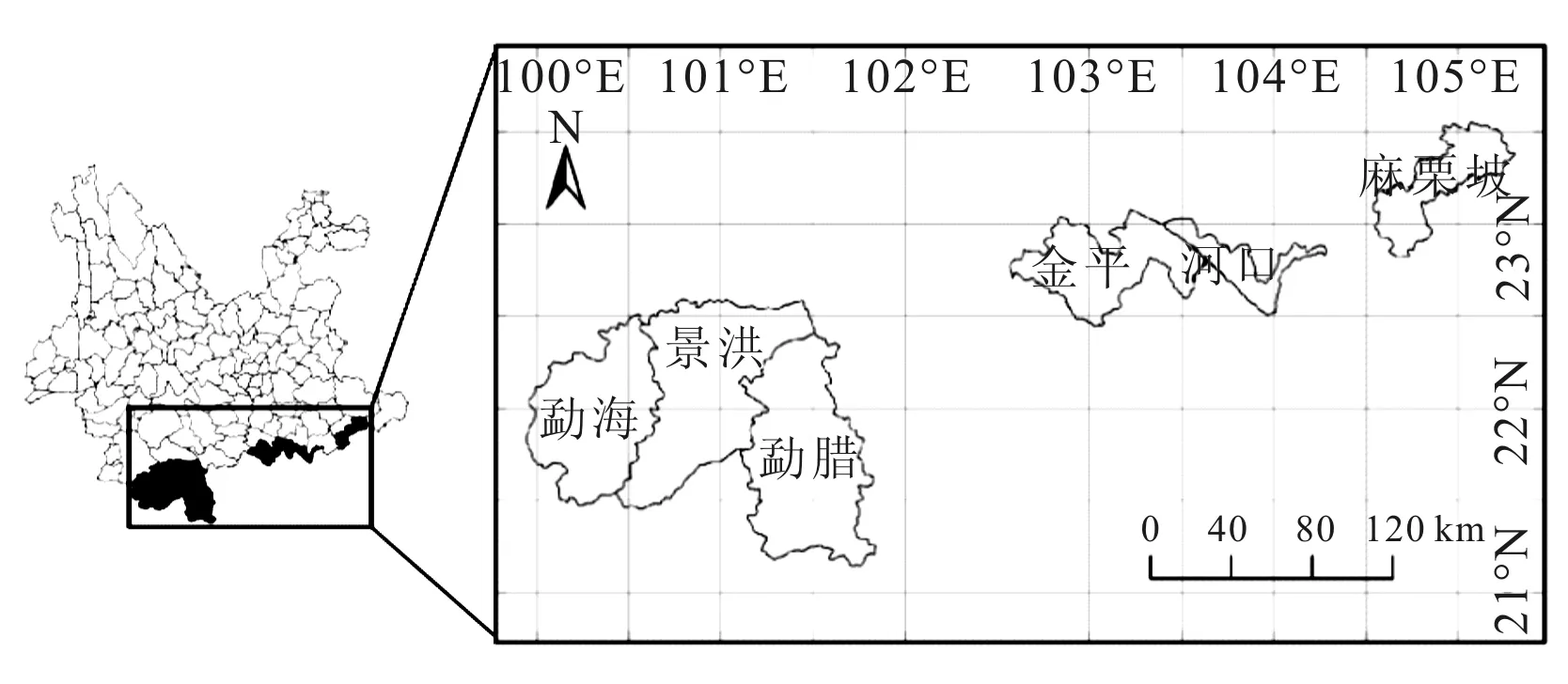
图1 研究区域的地理位置
2.2 物种分布数据
本研究的物种分布数据来源于中国数字植物标本馆(https://www.cvh.ac.cn/)和全球生物多样性信息网络(https://www.gbif.org/)提供的数字化标本记录。本研究依据最新的蕨类植物PPG I分类系统[1]以及《中国植物志》英文版(http://www.iplant.cn/foc)对所有标本记录的分类学名称进行了校对,通过R语言CoordinateCleaner包对标本记录的经纬度信息进行了校对,存在问题或信息不全的标本记录被剔除。通过ArcMap10.3将研究区域划分为332个10 km×10 km的栅格,各标本记录根据经纬度信息匹配到相应的栅格中,每个栅格中只保留每个物种的唯一出现记录,重复的记录被删除。最后,共保留标本记录3784条用于本次研究。
2.3 系统发育树
为了计算云南南部地区蕨类植物的系统发育多样性,本研究基于叶绿体rbcL基因片段构建了云南南部地区蕨类植物的系统发育树。用于系统发育树构建的rbcL分子序列数据来自于GenBank数据库(https://www.ncbi.nlm.nih.gov/)以及测序实验。测序所用分子材料取自蕨类植物叶片标本。本研究采用北京全式金生物科技有限公司的Plant Genomic DNA Kit试剂盒对蕨类植物的总DNA进行了提取,并参照Schuettpelz 和 Pryer[10]的方法对目标片段进行了扩增,扩增产物被送至测序公司完成测序。最后,所有的序列在Geneious 8.1.3 软件中完成比对、校正和排序,利用RAxML插件程序构建出系统发育树。
2.4 生物多样性指数
本研究中各栅格的物种丰富度(SR)和系统发育多样性(PD)由R语言picante包计算产生,物种β多样性及其周转和嵌套组分(βsor.tax、βsim.tax和βsne.tax)和系统发育β多样性及其周转和嵌套组分(βsor.phy、βsim.phy和βsne.phy)由R语言betapart包计算产生。各多样性指数的空间分布格局,通过ArcMap10.3在地图中进行了可视化展示。
2.5 环境变量数据
本研究共选取了22个环境变量,用于分析蕨类植物多样性与环境变量的关系。其中,20个气候变量的数据来自于地球陆地表面高分辨率气候数据库(https://chelsa-climate.org/),分别为Bio1~Bio19和年潜在蒸散量(PET)。2个地形变量数据提取自CGLAR-CSI SRTM的高程数据(https://srtm.csi.cgiar.org/),分别为海拔(Alt)和坡度(Slope)。
2.6 统计分析
本研究通过Spearman相关分析检验了物种丰富度(species richness,SR)、系统发育多样性(phylogenetic diversity,PD)与各环境变量的相关性,通过Mantel test相关分析检验了物种β多样性及其周转和嵌套组分(βsor.tax、βsim.tax和βsne.tax)、系统发育β多样性及其周转和嵌套组分(βsor.phy、βsim.phy和βsne.phy)与地理距离(Geog.dist)和环境距离的相关性。地理距离(Geog.dist)为两两栅格地理中心之间的欧几里得距离。环境距离,包括气候距离(Clim.dist)和地形距离(Topo.dist),为两两栅格间气候和地形变量的欧几里得距离。使用LOWESS局部加权回归分析获取了系统发育多样性(PD)关于物种丰富度(SR)的回归模型。
3 研究结果
3.1 蕨类植物物种组成情况
标本记录的统计结果显示,云南南部地区共有蕨类植物37科116属690种,占云南蕨类植物总种数的52.6%。其中,含50种以上的科有5科,分别是水龙骨科Polypodiaceae、鳞毛蕨科Dryopteridaceae、凤尾蕨科Pteridaceae、金星蕨科Thelypteridaceae和蹄盖蕨科Athyriaceae,这5个科共有402种,占地区总种数的58.3%,是该地区主要的优势科。含30种以上的属共有5属,分别是铁角蕨属Asplenium、凤尾蕨属Pteris、三叉蕨属Tectaria、双盖蕨属Diplazium和卷柏属Selaginella,这5个属共有175种,占地区总种数的25.4%,是该地区主要的优势属。该地区出现频率较高的物种有薄叶卷柏Selaginelladelicatula、黑顶卷柏Selaginellapicta、攀缘卷柏Selaginellahelferi、垂穗石松Palhinhaeacernua、金毛狗Cibotiumbarometz、截裂毛蕨Cyclosorustruncatus、乌毛蕨Blechnumoriental、线羽凤尾蕨Pterisarisanensis、半月形铁线蕨Adiantumphilippense和裸叶石韦Pyrrosianudae等。
3.2 蕨类植物多样性的空间分布格局
云南南部地区蕨类植物物种丰富度(SR)的空间分布如图2(a)所示,物种丰富度(SR)的高值区域主要出现在勐海县的勐海镇、勐宋乡和布朗山地区,景洪市的纳板村、勐养以及勐宋村地区,勐腊县的勐仑镇、补蚌和尚勇地区,金平县的勐拉镇和金河镇地区,河口县的南溪镇地区以及麻栗坡县的下金厂乡、八布乡和老君山地区。
系统发育多样性(PD)的空间分布格局与物种丰富度(SR)相似(图2b)。物种丰富度(SR)较高的区域,系统发育多样性(PD)也相对较高,而物种丰富度(SR)较低的区域,系统发育多样性(PD)也相对较低。相关性分析显示,物种丰富度(SR)与系统发育多样性(PD)之间具有很强的正相关(r= 0.98)。
系统发育多样性(PD)关于物种丰富度(SR)回归模型残差的空间分布格局如图2(c)所示,部分物种丰富度(SR)高的区域系统发育多样性(PD)却相对偏低(残差小于0),如河口县的南溪镇、勐腊县的补蚌和尚勇等地区;部分物种丰富度(SR)高的区域系统发育多样性(PD)也较高(残差大于0),如勐海县的勐宋乡、勐腊县的勐仑镇、金平县的勐拉镇和麻栗坡县的老君山等地区。

图2 云南南部地区蕨类植物(a)物种丰富度、(b)系统发育多样性和(c)残差的空间分布
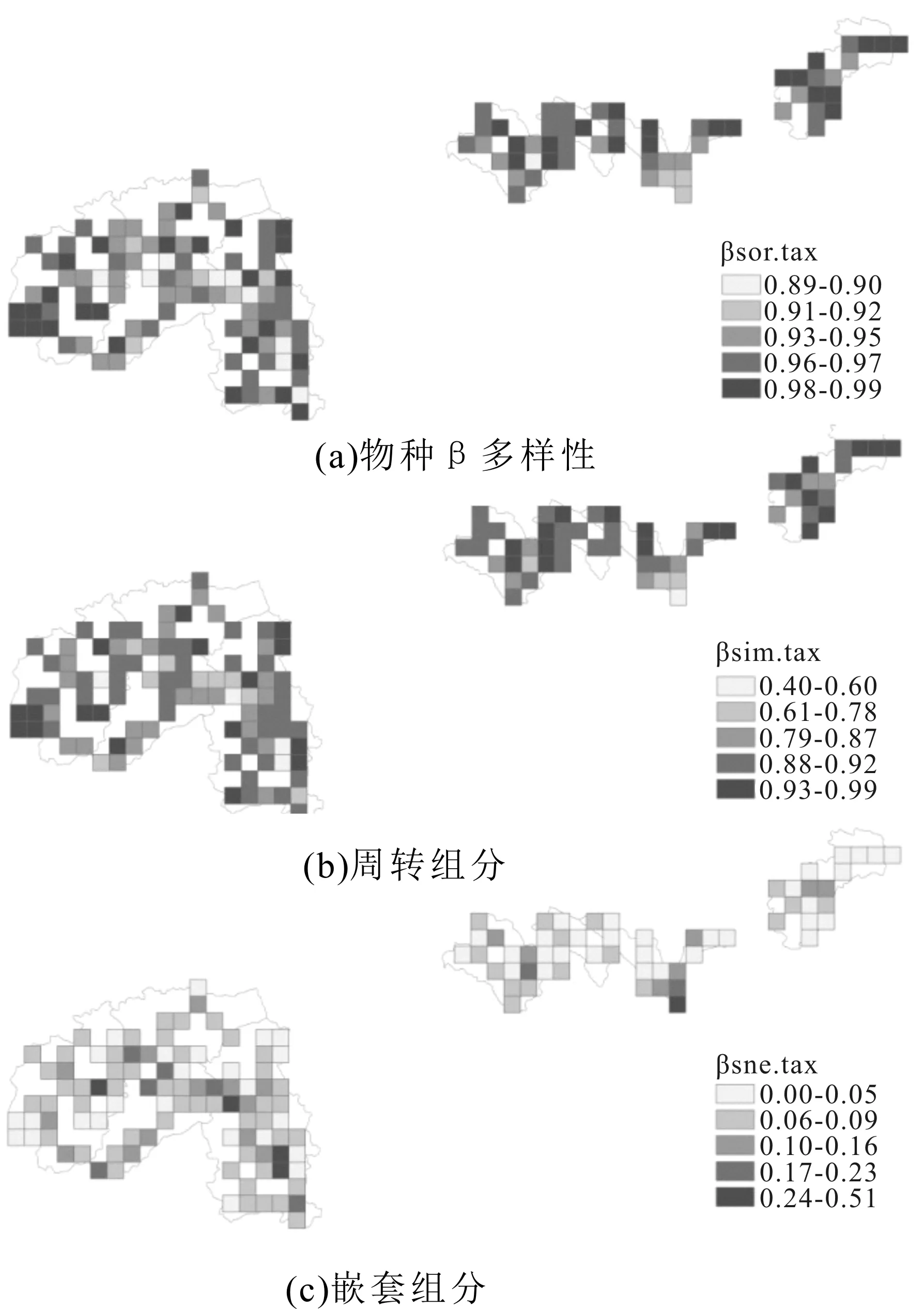
图3 云南南部地区蕨类植物(a)物种β多样性及其(b)周转和(c)嵌套组分的空间分布
云南南部地区蕨类植物的物种β多样性和系统发育β多样性(βsor.tax、βsor.phy)及其周转组分(βsim.tax、βsim.phy)和嵌套组分(βsne.tax、βsne.phy)的空间分布格局如图3和图4所示。云南南部地区所有栅格的物种β多样性(βsor.tax)及其周转组分(βsim.tax)和嵌套组分(βsne.tax)的平均值分别为0.96±0.03、0.87±0.09和0.08±0.07,系统发育β多样性(βsor.phy)及其周转组分(βsim.phy)和嵌套组分(βsne.phy)的平均值分别为0.64±0.07、0.34±0.13和0.31±0.09。物种β多样性(βsor.tax)及其周转组分(βsim.tax)高于系统发育β多样性(βsor.phy)及其周转组分(βsim.phy),而物种β多样性的嵌套组分(βsne.tax)低于统发育β多样性嵌套组分(βsne.phy)。物种β多样性的周转组分(βsim.tax)远高于嵌套组分(βsne.tax),而系统发育β多样性的周转组分(βsim.phy)和嵌套组分(βsne.phy)相当。

图4 云南南部地区蕨类植物(a)系统发育β多样性及其(b)周转和(c)嵌套组分的空间分布
物种β多样性和系统发育β多样性(βsor.tax、βsor.phy)与其周转组分(βsim.tax、βsim.phy)和嵌套组分(βsne.tax、βsne.phy)的相关分析结果显示,物种β多样性和系统发育β多样性(βsor.tax、βsor.phy)与其周转组分(βsim.tax、βsim.phy)均有极显著的正相关(r=0.88,P<0.001;r=0.35,P<0.001);物种β多样性(βsor.tax)与其嵌套组分(βsne.tax)有极显著的负相关(r=-0.69,P<0.001),系统发育β多样性(βsor.phy)与其嵌套组分(βsne.phy)的负相关不显著(r=-0.10,P>0.05);物种β多样性和系统发育β多样性的周转组分(βsim.tax、βsim.phy)与其嵌套组分(βsne.tax、βsne.phy)均有极显著的负相关(r=-0.93,P<0.001;r=-0.94,P<0.001)。
3.3 蕨类植物多样性与环境变量的关系
云南南部地区蕨类植物的物种丰富度(SR)和系统发育多样性(PD)与环境变量的Spearman相关分析结果如表1所示。物种丰富度(SR)和系统发育多样性(PD)与气候变量的相关性较强,与地形变量的相关性较弱。物种丰富度(SR)与年均温(Bio1)、最冷月最低温(Bio6)、最暖月均温(Bio10)以及最冷季降水(Bio19)的相关性最强,系统发育多样性(PD)与最冷月最低温(Bio6)以及最冷季降水(Bio19)的相关性最强。物种丰富度(SR)和系统发育多样性(PD)与降水的季节性(Bio15)均有显著的负相关。物种丰富度(SR)和系统发育多样性(PD)与海拔(Alt)和坡度(Slope)两个地形变量均只有较弱的负相关。
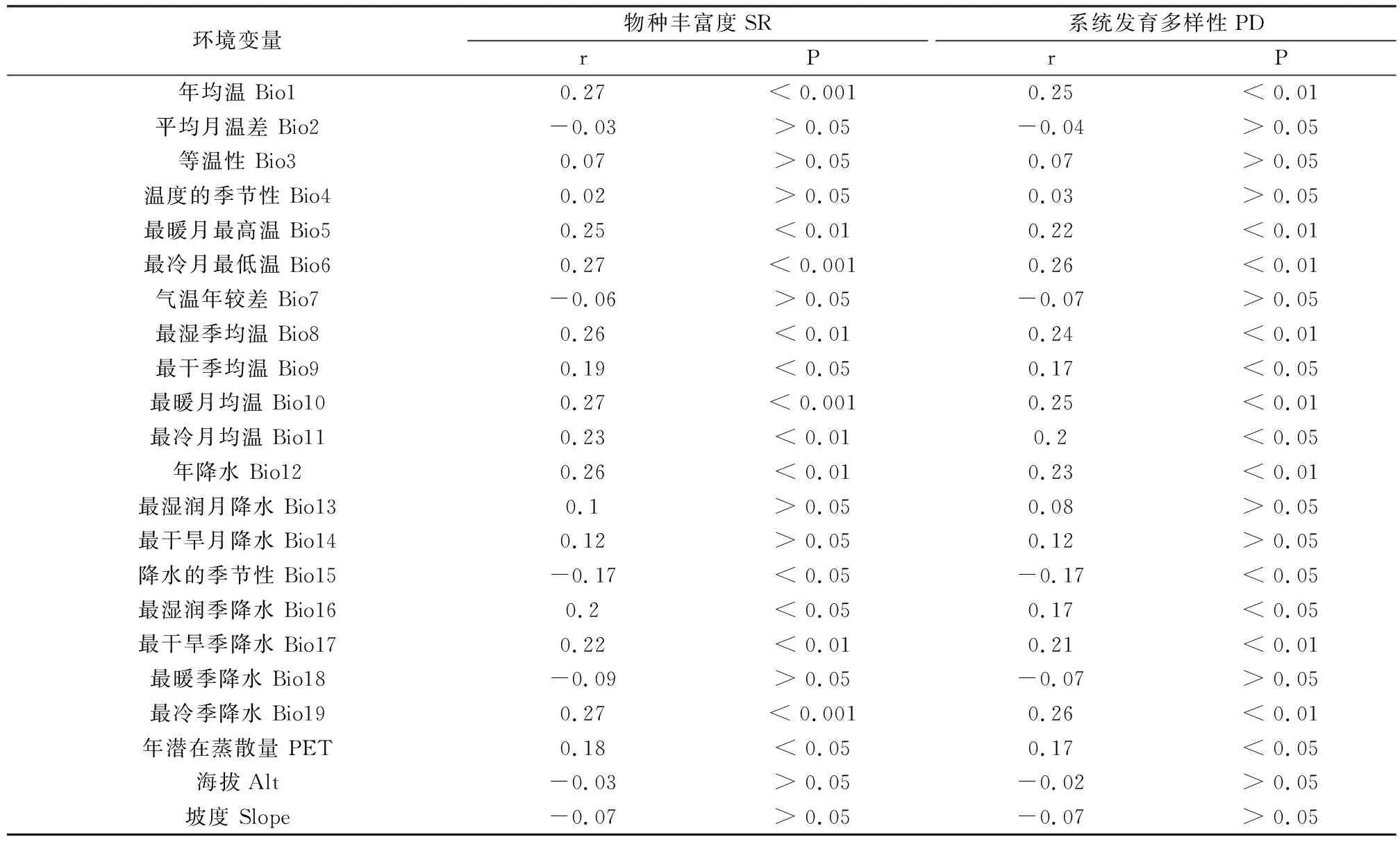
表1 云南南部地区蕨类植物物种丰富度和系统发育多样性与环境变量的关系
云南南部地区蕨类植物的物种β多样性和系统发育β多样性(βsor.tax、βsor.phy)及其周转组分(βsim.tax、βsim.phy)和嵌套组分(βsne.tax、βsne.phy)与地理距离(Geog.dist)、气候距离(Clim.dist)以及地形距离(Topo.dist)的Mantel test相关分析的结果如表2所示。物种β多样性和系统发育β多样性(βsor.tax、βsor.phy)与地理距离(Geog.dist)、气候距离(Clim.dist)以及地形距离(Topo.dist)均表现出一定的相关性,但物种β多样性(βsor.tax)与各距离矩阵的相关性更强。物种β多样性的周转组分(βsim.tax)与各距离矩阵也有较强的相关性,系统发育β多样性的周转组分(βsim.phy)只与地理距离(Geog.dist)有一定的相关性。物种β多样性和系统发育β多样性的嵌套组分(βsne.tax、βsne.phy)与各距离矩阵的相关性都不强。
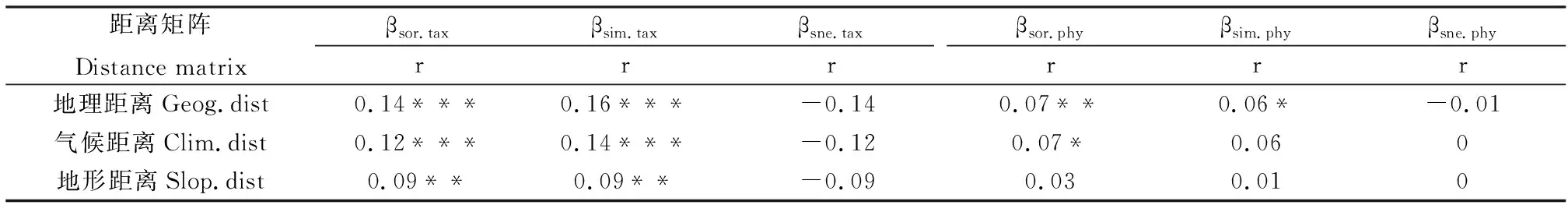
表2 云南南部地区蕨类植物β多样性与地理距离、气候距离及地形距离的关系
4 讨论
研究结果显示,云南南部地区的蕨类植物多样性水平较高,物种组成丰富。云南南部地区位于亚洲热带的北缘,是印度-缅甸生物多样性热点地区的重要组成部分,分布有大量的蕨类植物。云南南部地区虽然只占云南总面积的7%,但是却拥有云南一半以上的蕨类植物物种。因此,该地区是云南蕨类植物多样性研究和保护的重点地区。
相关分析表明,云南南部地区蕨类植物的物种丰富度(SR)和系统发育多样性(PD)有具很强的正相关(r= 0.98)。表现在空间分布上,即物种丰富度(SR)和系统发育多样(PD)具有相似的空间分布格局。云南南部地区的勐宋乡、勐仑镇、勐拉镇和老君山等地区,既是蕨类植物物种丰富度(SR)的高值区域,也是系统发育多样性(PD)的高值区域。也有部分物种丰富度(SR)较高的区域,系统发育多样性(PD)却相对偏低,如南溪镇、补蚌和尚勇等地区。有研究发现,云南南部地区的历史气候稳定,在第四纪受冰期影响较小,保存了很多古老的谱系,如观音座莲属Angiopteris等[11]。同时,云南南部地区的季风气候和局域特殊地形加速了地区的物种分化,又使得该地区成为了生物进化的前沿[12,13]。因此,云南南部地区既保存了很多进化历史久远的古老谱系,是蕨类植物多样性的博物馆,也拥有大量亲缘关系较近的年轻谱系,是蕨类植物多样性的摇篮。
云南南部地区蕨类植物物种β多样性(βsor.tax)和系统发育β多样性(βsor.phy)也有很强的正相关。表现在空间分布上,即物种β多样性(βsor.tax)和系统发育β多样性(βsor.phy)的空间分布格局有较强的相似性。同时,本研究还发现,云南南部地区各栅格蕨类植物的物种β多样性(βsor.tax)普遍大于系统发育β多样性(βsor.phy)。而且,云南南部地区蕨类植物物种β多样性(βsor.tax)主要来自于周转组分(βsim.tax),嵌套组分(βsne.tax)对物种β多样性(βsor.tax)的贡献较小,系统发育β多样性的两个组分(βsim.phy、βsne.phy)对系统发育β多样性(βsor.phy)的贡献则相当。这些研究结果都表明,云南南部地区蕨类植物β多样性的形成,主要来自于亲缘关系较近的物种在空间上的交换,而不是亲缘关系较远的物种
研究结果显示,云南南部地区蕨类植物的物种丰富度(SR)和系统发育多样性(PD)与环境变量中的气候变量相关性较显著,而与地形变量相关性不显著。气候变量反映了地区水热条件的差异,根据水热动态假说,水分和热量条件与植物的生理活动密切相关,是影响植物多样性分布的主要因子[14]。蕨类植物由于自身生理和生殖结构的特殊性,对水分和热量条件的响应更为明显[15]。因此,蕨类植物的多样性与降水和温度等气候变量往往会有更显著的相关性。海拔和坡度等地形变量是衡量地区生境异质性的重要指标,根据生境异质性假说,地形越复杂、生境异质性越高的地区,所拥有的生态位也越多,植物多样性也更加丰富[16]。但是,本研究却发现,云南南部地区蕨类植物的多样性与地形或生境异质性没有显著的相关性,这可能与研究的尺度有关。生境异质性对多样性的影响通常会随着研究尺度的减小而降低[17],本研究使用的10 km×10 km的栅格尺度相对较小,无法反映大尺度的生境异质性特征对植物多样性的影响。同时,研究数据的完整性也会对分析结果产生影响,很多研究都发现标本记录数据普遍存在采集不完整和地理偏差等问题[18~20]。因此,在之后的研究分析中,研究者需要对数据的完整性进行评估,并通过野外考察对相关数据进行补充,已获得更准确的分析结果。
研究结果还显示,云南南部地区蕨类植物的物种β多样性、系统发育β多样性与地理距离和环境距离都有显著的相关性。表明了扩散限制和环境过滤过程对云南南部地区蕨类植物物种β多样性和系统发育β多样性的形成都有重要的作用。同时,地理距离与物种β多样性有更强的相关性,反映了扩散限制过程对云南南部地区蕨类植物物种β多样性的形成有更重要的作用。
猜你喜欢
科学家(2022年5期)2022-05-13
重庆理工大学学报(自然科学)(2022年1期)2022-02-18
北方经贸(2021年12期)2021-12-25
北京航空航天大学学报(2021年5期)2021-06-09
意林(2020年3期)2020-03-17
大科技·C版(2018年11期)2018-10-21
中国总会计师(2017年12期)2018-02-06
电影文学(2017年24期)2017-11-16
金融经济(2017年7期)2017-07-15
大众理财顾问(2016年9期)2016-10-11
