
梭梭和白梭梭的种群结构与空间分布格局
2021-09-25 02:49:46孙芳芳马松梅王春成张云玲
西南农业学报 2021年8期
闫 涵,孙芳芳,马松梅,王春成,张 丹,张云玲
(1.石河子大学理学院,新疆 石河子 832000;2.石河子大学生命科学学院,新疆 石河子 832000;3.新疆维吾尔自治区草原总站,新疆 乌鲁木齐 830049)
【研究意义】植物种群结构及数量动态是种群生态学研究的核心问题,对了解种群更新策略、群落的形成和稳定性等都具有重要的意义[1]。种群结构包括年龄结构、高度结构和冠幅结构,反映了种群不同个体的配置情况,也体现了种群与环境间的相互关系[2-3]。种群结构按年龄组成,可分为增长型、稳定型和衰退型。种群动态是指种群大小或数量在时间、空间上的变化规律[4]。生命表和存活曲线是进行种群统计和研究种群数量动态变化的重要工具[5]。种群的空间分布格局是种群在水平空间上的配置或分布状况,受种群特性和环境条件等因素的综合影响,一般分为随机分布、聚集分布和均匀分布三种类型[6-8]。研究植物空间分布格局,可以揭示植物种群与生境的相互作用关系及其生态学过程[9-10]。因此,利用植物种群的生长指标、生命表、存活曲线等研究植物的种群结构及其数量动态,利于更好地认识种群的发展规律及其影响因素,对种群的更新复壮及生态系统的恢复与发展都具有重要的理论意义和应用价值[11]。世界上藜科(Chenopodiaceae)梭梭属(HaloxylonBunge)植物分布有11种,我国仅分布2种:梭梭(Haloxylonammodendron)和白梭梭(H.persicum)[12]。梭梭主要分布于我国西北地区,且集中分布于新疆准噶尔盆地、塔里木盆地,约占全国梭梭林总面积的68.2%[13]。而白梭梭在我国主要分布于新疆准噶尔盆地:乌苏、沙湾、精河县、阜康北沙窝等地[14],其中甘家湖梭梭林国家级自然保护区是梭梭和白梭梭的重要分布地。梭梭和白梭梭都是灌木状小乔木,是干旱荒漠区的主要建群种,具有防风、固阻沙以及维持生物多样性等重要的生态作用[15]。然而,长期以来,由于自然作用(玛纳斯河断流、地下水位下降等)和人为因素(土地开发、放牧、道路和油田工程建设等)对荒漠植被的严重破坏,造成大面积的天然梭梭林衰退甚至死亡[16-18]。【前人研究进展】梭梭和白梭梭都具有重要的科研价值,近年来,对两种植物已开展的研究主要体现在种子萌发、天然林更新复壮、封沙育林、抗旱生理以及遗传多样性[19-21]等方面。围绕新疆梭梭和白梭梭的种群结构及空间格局已开展的研究较少。对甘家湖梭梭和白梭梭的种群动态和对石河子梭梭种群的性状研究都表明两种梭梭的种群结构为稳定型[22-23]。对甘家湖、克拉玛依农业综合开发区外围荒漠和玛纳斯河下游的梭梭种群分布格局的研究表明:梭梭在0~2 m或0~12 m的小尺度上为聚集分布,在大于12 m的尺度上为随机分布或均匀分布[24-25]。【本研究切入点】目前,对古尔班通古特沙漠南缘自然分布的梭梭和白梭梭种群的生长特征、种群结构、种群数量动态及种群的空间分布格局还缺乏系统研究,成为科学管理与恢复我国现有梭梭和白梭梭荒漠植被的瓶颈。【拟解决的关键问题】本研究在野外调查研究的基础上,整合种群统计、种群生态学和统计学分析方法,借助地理信息系统GIS软件,研究甘家湖自然保护区梭梭和白梭梭的种群现状与空间分布格局,旨在解决以下科学目标:①探明甘家湖梭梭和白梭梭的种群结构及其数量动态,探讨其未来发展趋势;②分析甘家湖梭梭和白梭梭种群的空间分布格局。
1 材料与方法
1.1 样地设置
根据研究区梭梭和白梭梭的实际分布,在甘家湖梭梭自然保护区内,选择长势较一致的梭梭和白梭梭自然种群,利用测量绳分别标定了一个50 m×50 m研究样地。对样地内的所有个体,都利用GPS定位、记录经纬度和海拔,并记录每一个样地的生境特点和伴生植物的类型(表1)。

表1 梭梭和白梭梭样地的基本特征
1.2 种群的数量特征分析
在设置的甘家湖样地,根据中国科学院生态系统调查表对乔木和小乔木的株高、冠幅及基径调查的要求[26],利用卷尺、皮尺和测量绳测量所有梭梭和白梭梭活株个体(除幼苗外)的株高(对于株高大于150 cm 的个体,架设便携式梯子XB-3001进行测量)和冠幅(取东西、南北的平均值);利用游标卡尺测量基径。
对甘家湖样地梭梭和白梭梭种群的每个生长指标都进行正态分布检验和描述性统计分析,以揭示两种植物种群的数量特征。具体是:利用Kolmogorov-Smirnov正态分布检验的D=max|Fn(x)-F0(x)|统计量[28]检验各生长指标的正态性。检验时,根据样本计算检验统计量Dx,比较样本分布形状的检验统计量Dx和正态分布统计量D的差异显著性,判断生长指标的分布特征是否符合正态分布。基于Pearson相关分析,利用SPSS19.0分析梭梭和白梭梭种群各生长指标之间的相关性。
1.3 种群的龄级结构及动态分析
参考吕朝燕等[27]和蔡新斌等[23]对准噶尔盆地西北缘梭梭种群和甘家湖自然保护区白梭梭种群结构的研究,本文采用以梭梭和白梭梭的基径大小代替龄级的方法,分析两种植物种群的年龄结构特征。具体是,对梭梭和白梭梭的基径按照每3 cm为一级进行龄级划分,基径低于3 cm按幼苗统计,这样共分为9个龄级:幼苗(Ⅰ级)、幼树(Ⅱ级)、小树(Ⅲ级~Ⅳ级)、中树(V级~Ⅵ级)、大树(Ⅶ级~Ⅷ级)和老树(IX级)[23],分别统计不同龄级梭梭与白梭梭的个体数。
编制梭梭和白梭梭的静态种群生命表,根据公式计算以下指标:
lx=(ax/a0)×1000
dx=lx-lx+1
qx=(dx/lx)×1000
sx=1-qx
式中:x为龄级数;ax为在x龄级内的现有个体数;lx为在x龄级开始时标准化存活个体数(一般转化值为1000);dx为从x到x+1龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;sx为从x到x+1龄级间隔期间存活率。
以龄级为横坐标,以标准化存活个体数的对数为纵坐标,绘制存活曲线。植物种群的存活曲线分为:DeeveyⅠ型(凸型),种群表现为低龄级的个体死亡率较低,存活率较高,种群属于增长型;DeeveyⅡ型(直线型),种群各龄级的存活率基本相同,种群属稳定型;DeeveyⅢ型(凹型):种群低龄级的个体有极高的死亡率,种群属衰退型[28]。
1.4 种群的空间点格局分析
根据植物种群的密度与盖度的计算公式:密度=样地内某种植物的个体数/样地面积,盖度=样地内某种植物投影面积/样地面积[29],分别计算甘家湖自然保护区梭梭和白梭梭的种群密度(株/公顷)与盖度。在0~50 m的空间尺度上,利用Programita软件,基于Ripley'sK(r)函数对各样地内定位的梭梭和白梭梭种群进行点格局分析[26]。利用Monte-Carlo拟合检验方法计算上、下包迹线,即置信区间。假定种群是随机分布,利用随机模型拟合一组点的坐标值,对不同空间尺度上r的值,都计算物种的个体数L(r),这一过程重复模拟99次,以不同空间尺度下L(r)的最大值和最小值分别作为上下包迹线的坐标值。再以空间尺度r为横坐标,用种群实际分布数据计算得到的不同尺度下的L(r)值作为纵坐标,绘制种群的点格局分布图。若实际值L(r)位于上包迹线上方,则种群为聚集分布;若实际值L(r)位于上、下包迹线之间,则种群为随机分布;若实际值L(r)位于下包迹线下方,则种群为均匀分布。
2 结果与分析
2.1 种群数量特征的描述性统计分析
甘家湖梭梭和白梭梭种群的株高、冠幅和基径观测数列的K-S正态性检验结果表明,梭梭种群生长指标的阈值范围分别为S∈[-0.7132,0.7132]、K∈[-1.4357,1.4357];白梭梭种群的阈值范围分别为S∈[-0.6638,0.6638]、K∈[-1.2253,1.2253]。两种梭梭种群的生长指标均符合正态分布(表2),可以进行进一步分析。描述性统计表明:两种梭梭种群的生长指标均达到强度变异(变异系数>15%);除白梭梭株高外,其余指标均为平峰偏向右侧分布(峰度值>0,偏度值>0),基径偏离程度最大,说明两种梭梭处于低龄级的个体数较多;白梭梭的株高指标表现为尖峰(峰度值<0),其对称轴两侧的极端数据偏少(表2),说明白梭梭存在高龄级的个体数也较多的现象。

表2 梭梭和白梭梭种群数量特征的描述性统计
2.2 种群结构的动态分析
梭梭和白梭梭的株高、冠幅和基径之间均呈极显著的正相关关系(P<0.01),相关系数均大于0.531和0.593(表3)。

表3 梭梭和白梭梭株高、冠幅和基径的相关性
甘家湖梭梭和白梭梭的基径结构总体均呈倒“J”型分布,说明均为增长型种群(图1)。根据基径分级结果,梭梭的大多数个体集中在小于Ⅳ级(幼苗、幼树和小树),占比为86.09%;而白梭梭的大多数个体集中在小于Ⅱ级(幼苗和幼树),占比为78.26%。
梭梭和白梭梭种群的静态生命表显示(表4~5),两种梭梭种群均表现为龄级小于Ⅱ级(幼苗,幼树)的个体数量最多。甘家湖梭梭和白梭梭种群的存活曲线总体均趋近于DeeveyⅠ型(凸型,图2):在龄级小于Ⅴ级范围内呈平缓下降趋势,在Ⅴ级以后均呈显著下降趋势,表现为低龄级的个体死亡率较低和存活率较高,种群属于增长型。
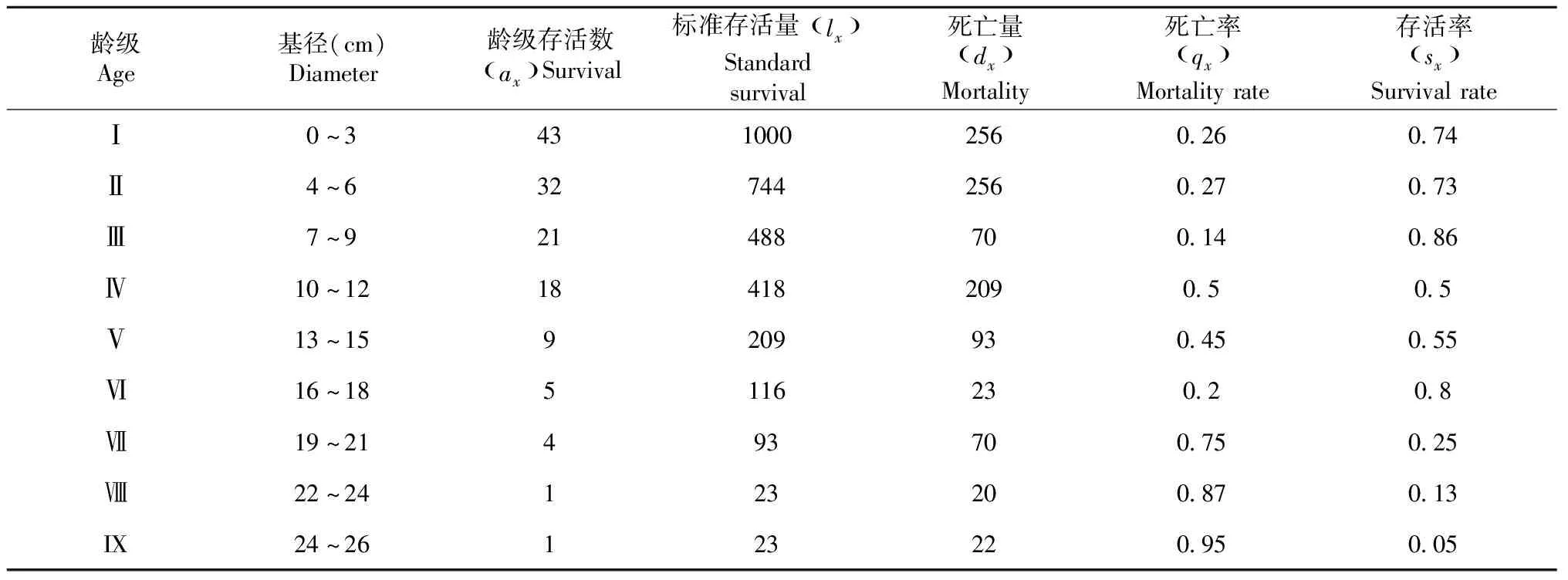
表4 甘家湖梭梭种群的静态生命

表5 甘家湖白梭梭种群的静态生命
2.3 种群的空间分布格局分析
甘家湖自然保护区梭梭和白梭梭的种群密度分别为4480和3920株/公顷,盖度分别为11.66%和11.32%。梭梭和白梭梭种群的点格局分析结果表明,在甘家湖自然保护区,梭梭和白梭梭种群分别在0~2 和0~3 m的尺度上呈聚集分布,在大于2 和3 m的尺度上均呈随机分布(图4)。
3 结论与讨论
3.1 种群的结构特征及其数量动态
种群结构分析可以直观地反映种群的现存状态、数量动态特征,以及种群与环境之间的相互关系[29]。本研究种群结构及数量动态方面的研究结果均表明甘家湖自然保护区样地梭梭和白梭梭种群为增长型,总体表现为低龄级(幼苗,幼树和小树)个体数较多,高龄级(大树和老树)个体数较少的特点。有学者对甘家湖、准噶尔盆地西北缘和石河子地区的梭梭种群结构的研究也都表明梭梭种群为增长型[23-24,30]。总体来看,甘家湖梭梭种群处于稳定发展阶段,且自然更新较好。
甘家湖梭梭和白梭梭的各生长指标数量特征总体均符合正态分布,个体间的变异性较强,低龄级的个体数较多,也说明种群的更新能力及稳定性较强,这与有关学者对古尔班通古特沙漠南缘的梭梭种群的研究结果相似[23]。甘家湖自然保护区近年来降雨量适宜梭梭生长,加上围栏封育,禁牧或少牧的人为措施,天然梭梭林处于大面积恢复阶段[18]。另外,梭梭和白梭梭种群的3个生长指标间均呈极显著的相关关系,这也与相关研究结果相似[23,30],说明选择基径指标替代株高、冠幅是可行的。
本研究甘家湖样地内两种植物种群的幼苗数量较为丰富(占总体的25%),低龄级个体死亡率相对高龄级也较低(<27%),并且植株在Ⅱ级数量较多(存活率大于72%,图1,表4~5),也说明种群处于稳定发展阶段。野外调查发现,研究样地内梭梭和白梭梭种群的幼苗多聚集分布在母树树冠周围。这可能是由于两种植物的种子主要散布在母株周围,且母树的肥岛效应促使幼苗分布较为集中,存活率相对较高[31-32]。而梭梭和白梭梭种群中,高龄级(Ⅶ-IX级)个体的死亡率较高(> 75%),生存曲线属于DeeveyⅠ型(凸型)(表4~5,图2),这可能是因为两种梭梭种群的高龄级个体间为争夺有限的资源的竞争更为激烈,造成植株的死亡率相对较高[33]。
3.2 种群的空间分布格局
种群的空间分布格局主要由种群本身的生物学特性、种内种间关系及环境条件共同影响决定,是种群生态关系在空间上的重要表现形式,能在一定程度上解释群落结构的生态学过程与环境变化的关系。本研究中,甘家湖的梭梭和白梭梭种群在较小尺度(<7 m)内呈聚集分布,这与已有研究结果相似[25-26]。另外,对濒危植物胡杨种群的空间分布格局研究也表明在小尺度范围内(<16 m)呈聚集分布特征[34]。小尺度范围内的聚集分布是很多濒危植物种群分布的一般特征[35]。一般来说,“群集有利于生存”,集群分布能更好地改变种群的微气候和小生境,因而集群分布提高了对不良环境的抗性[36]。生境异质性也是种群聚集分布的重要驱动因子,因此当外界条件比较恶劣的时候,植物会主动调整生物量分配模式,在小尺度上聚集分布,以适应荒漠资源贫瘠、干旱少雨的气候条件,以最大效率地利用资源,从而提高自身的生存适合度[37]。但本研究中甘家湖梭梭和白梭梭种群生境相似,却在小尺度内呈聚集分布,可能是由于随着植株个体的发育,其对外界环境条件需求增加,个体间竞争加剧,造成梭梭和白梭梭种群较高龄级的数量减少,其结果是产生随机扩散,逐步形成随机分布[38]。因此,甘家湖样地内梭梭和白梭梭种群的空间分布格局不同,可能是生物因子(如物种特性、种间竞争、种内竞争等)和非生物因子(如水、风、光照、土壤、微地形等)在种群不同发育阶段长期综合作用的结果。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
哈哈画报(2022年4期)2022-04-19 11:11:54
科学大众(2020年17期)2020-10-27 02:48:48
河南农业大学学报(2020年2期)2020-05-22 02:56:30
四川林业科技(2019年5期)2019-07-07 07:14:30
山西林业科技(2019年1期)2019-05-27 12:24:52
中学生天地(B版)(2019年4期)2019-05-08 03:22:54
安徽农学通报(2019年1期)2019-02-14 02:25:08
湖北林业科技(2019年6期)2019-01-06 02:14:29
湖北林业科技(2017年2期)2017-05-25 18:44:32
