
太行山南麓不同龄级荆条光合特性及光响应研究
2020-05-22 02:56刘雅辰赵琛迪杨子胡启立赵勇
河南农业大学学报 2020年2期
刘雅辰, 赵琛迪, 杨子, 胡启立, 赵勇
(河南农业大学林学院,河南 郑州 450002)
荆条(Vitexnegundo)是落叶灌木或小乔木,分布于中国河北、山西、山东、河南、陕西、甘肃等地,多生长于山地阳坡及林缘,抗旱耐寒,为中旱生灌丛的优势种。因其生命力顽强,耐旱性较高,被用来进行干旱半干旱地区的生态恢复。受到地理因素以及气候因素的限制,太行山南麓水资源缺乏,生态环境脆弱,荆条作为优势灌木在此广泛存在,并在太行山地区的生态恢复中占有重要地位。目前已经有许多学者对荆条灌丛进行了研究。赵河等[1-2]对荆条土壤养分的吸收、根系结构与生物量、养分变化以及微生物种类进行了研究,袁晓柯等[3]对荆条灌丛下不同林木幼苗根系周围土壤颗粒分形特征进行了研究,而对荆条的光合特征研究较少。光合作用是自然界中植物将太阳能转化成化学能,将无机物转变成有机物的重要过程[4],同时也对实现自然界的能量转换、维持大气的碳氧平衡具有重要意义[5]。光合是植物生长发育的基础,光合参数则直接反映了植物的生长特征[6-7]。因此,研究不同龄级荆条光合指标的差异对太行山南麓地区生态恢复有重要意义。本研究以太行山南麓地区荆条为研究对象,分析了不同龄级荆条的光合生理生态特征,探索其光合、蒸腾速率以及水分利用效率的变化规律,比较不同龄级的荆条光合参数指标、光合速率的光响应差异,从而分析其环境适应性之间的关系,为太行山南麓地区荆条灌丛的生态恢复提供理论依据。
1 研究地区概况及试验方法
1.1 研究地概况
试验地位于河南省济源市国家林业局小浪底生态站,属太行山南麓地区,试验地坐标范围为E112°24′~ 112°32′,N34°58′~35°04′,年平均降水量400 mm,海拔300~400 m,气候为温带大陆性季风气候,属干旱半干旱地貌特征。植被类型主要有裸露地、草本植物、灌木和乔木,草本植物主要有魁蒿(Artemisiaprinceps)、牛筋草(Eleusineindica)和狗尾草(Setariaviridis)等;灌木主要有荆条(Vitexnegundo)、鼠李(Rhamnusbungeana)、胡枝子(Lespedezabicolor)和酸枣(Ziziphusjujube)等,乔木主要有栓皮栎(Quercusvariabilis)、刺槐(Robiniapseudoacacia)、侧柏(Platycladusorientalis)等[8]。
本试验参照MUDRAK等[9]对灌木龄级的划分方法,测定生长于土壤养分异质空间内的荆条在不同生长阶段的特征,对荆条进行龄级划分,确定试验对象。本试验根据数据模型公式共设置5个荆条龄级,龄级1以Vshrub<0.87 m3为形态最小型的幼年荆条,龄级2以0.87 m3
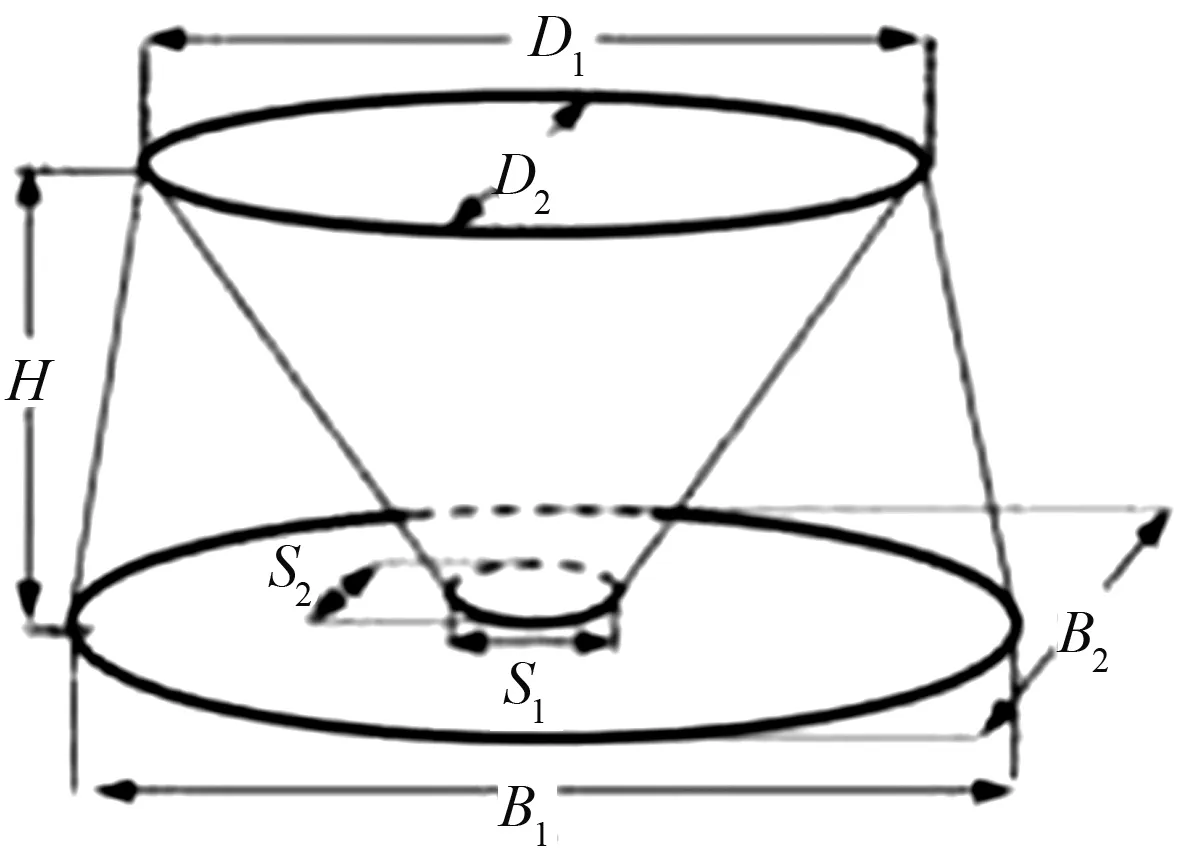
图1 荆条龄级划分模型示意图Fig.1 Schematic diagram of age classification model of Vitex negundo
ttop=0.25πD1D2
Abase=0.25π1B1B2
Astem=0.25πS1S2
式中:D1为灌木顶部长度;D2为灌木顶部宽度;B1为灌木底部长度;B2为灌木底部宽度;S1为灌木主茎长度;S2为灌木主茎宽度;Atop为灌木顶部面积;Abase为灌木底部面积;Astem为灌木主茎面积;Vshrub为灌木整体体积。按照实际测定,计算灌木体积大小。
1.2 试验方法
荆条叶片边缘有刻缺状锯齿,浅裂以至深裂,2019年5月,在天气晴朗的上午(9:00—11:00)选取5个龄级中相对独立生长的荆条,每个龄级3个重复,使用CIRAS3便携式光合仪选取1 mm×3 mm叶室,避开叶脉,测定叶片的光合生理生态指标,包括净光合速率(Pn)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)、蒸腾速率(E)、饱和水蒸气压亏缺(VPD)以及水分利用效率(WUE)。不同龄级荆条光合指标日变化的测定选取2019-05-24—2019-05-27的7:00—17:00,每隔2 h监测一次。光响应曲线测定时利用仪器自带光曲线与CO2曲线测定程序进行测定。取光合有效辐射(RAP)梯度为0、50、100、200、400、600、800、1 000 μmol·m-2·s-1进行测定。使用直角双曲线修正模型[10]:
式中:α为光响应曲线的初始斜率,β和γ为系数,I为光合有效辐射,Rd为暗呼吸速率。
饱和光照强度Isat为:
最大净光合速率Pnmax为:
叶片含水率的测定:从测定光合的荆条上摘下20 g成熟健康的鲜叶,放入105 ℃烘箱中杀青1 h,70 ℃烘至恒重,计算
叶片含水率=(叶片鲜重-叶片干重)/
叶片鲜重×100%
1.3 数据处理
应用Excel 2010和SPSS 25.0统计分析软件进行数据处理。
2 结果与分析
2.1 不同龄级荆条光响应曲线的变化规律
从图2可以看出,不同龄级荆条Pn对光照强度变化的响应趋势基本一致,但光合能力有所不同,当PAR小于400 μmol·m-2·s-1时,各龄级荆条Pn基本呈直线上升趋势,PAR大于400 μmol·m-2·s-1后各龄级荆条Pn增加明显变缓,各龄级在PAR达到800 μmol·m-2·s-1时趋于平缓,达到光饱和点。从图2可以看出,龄级3表现出更强的光合能力,在PAR的变化过程中,中龄级植株叶片Pn明显高于低龄级与高龄级植株叶片Pn。总体来说,中龄级荆条叶片处于生长旺盛阶段,光合效率更高,对光的利用能力较其他龄级效率更高。
2.2 不同龄级荆条光合生理生态因子的比较
2.2.1 不同龄级荆条Pn与光合生理生态因子的
相关分析 叶片的Pn是各种生理生态因子综合作用的结果,不同生长阶段的光合生理生态因子对其Pn的影响是不同的。通过对不同龄级荆条的Pn与相关生理生态因子相关分析(表1)得知,龄级1荆条Pn与Ci呈显著负相关关系,与VPD呈显著负相关关系;龄级3荆条Pn与Gs、E呈极显著正相关关系,与VPD呈显著负相关关系;龄级4荆条Pn与Gs、E、WUE呈极显著正相关关系,与VPD呈极显著负相关关系;龄级2和龄级5并没有表现出与光合生理生态因子的显著相关关系,各龄级荆条Pn与叶片含水率也没有表现出明显的相关关系。这说明不同龄级荆条的Pn受生理生态因子的影响不同,荆条Pn是其本身性质与各种因子的共同作用的复杂生理过程[11-12],其中Gs、E和VPD是影响各龄级荆条Pn的相对稳定因子。
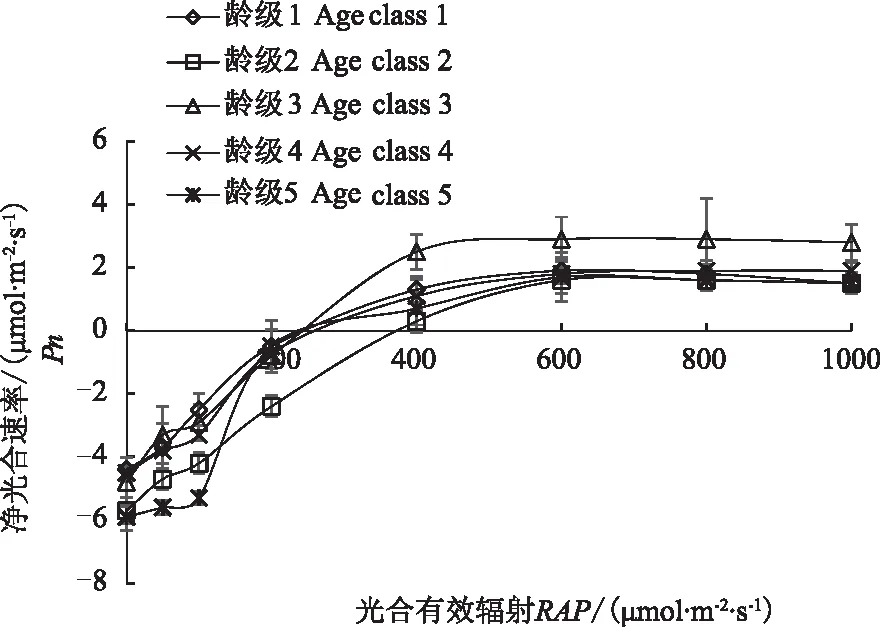
图2 不同龄级荆条叶片光响应曲线的变化Fig.2 Light response curves of leaves ofVitex negundo of different age classes

表1 不同龄级荆条Pn与光合生理生态因子的相关关系
注:**,在0.01水平(双侧)上显著相关; *,在 0.05 水平(双侧)上显著相关。
Note: ** means significant correlation at 0.01 level (bilateral) and * means significant correlation at 0.05 level (bilateral).
2.2.2 不同龄级荆条光合因子的差异分析 由表2可知,龄级1的Ci显著高于龄级3;Pn方面呈现明显的变化规律:龄级3显著高于龄级1、2、4、5;龄级3的Gs显著高于龄级1、5;龄级3 的E显著高于龄级1、2、5;龄级5的VPD显著高于龄级1;龄级3 的WUE显著高于龄级2、5。综合来看,龄级3的荆条光合能力更强,处于生长的旺盛期,龄级4荆条光合能力较龄级3略弱,龄级1、2、5光合能力更弱。
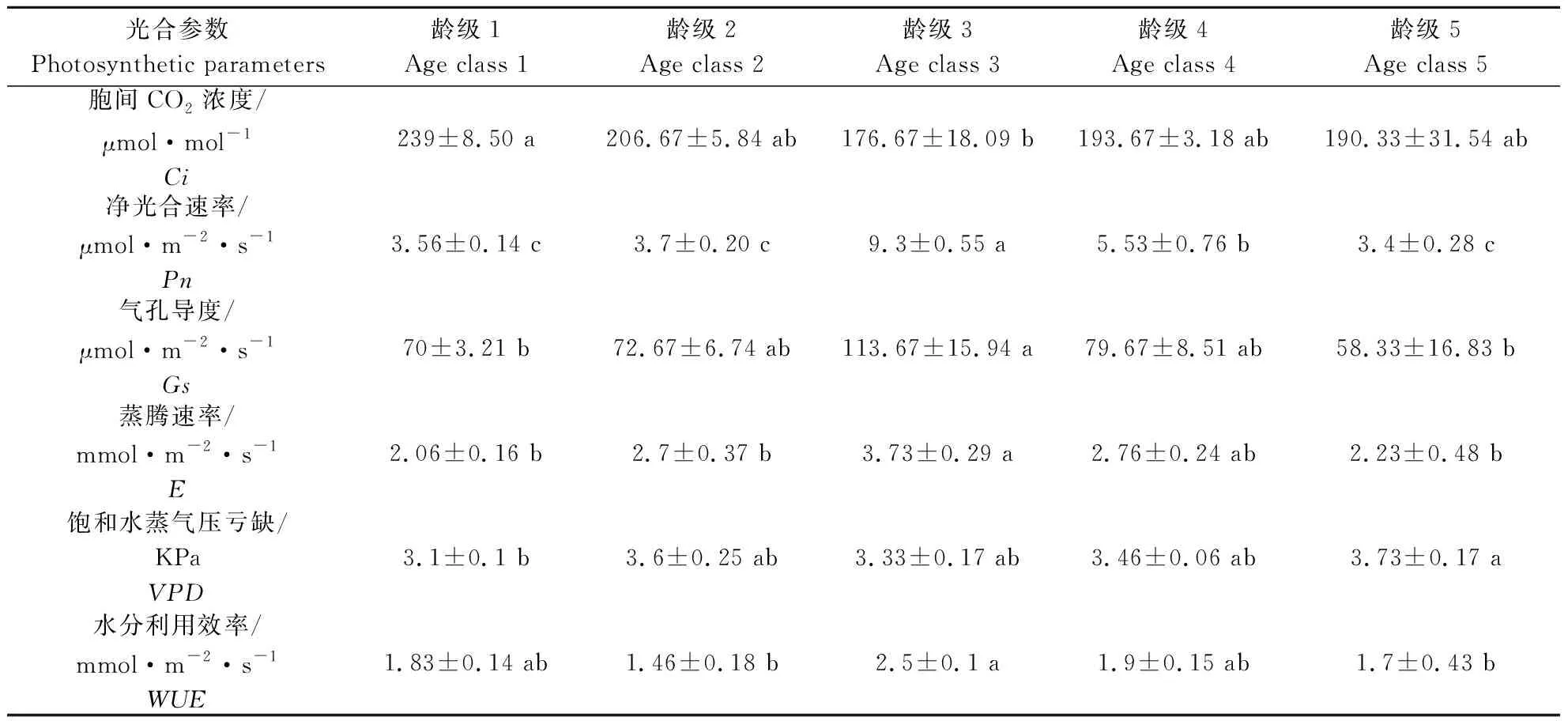
表2 不同龄级荆条的光合因子分析Table 2 Photosynthetic parameters analysis of Vitex negundo of different age classes
注:同行不同小写字母表示不同龄级具有显著性差异(P<0.05)。
Note: Different lower case letters in the same row indicate significant differences among different age classes(P<0.05).
2.3 不同龄级荆条光合参数的日变化规律
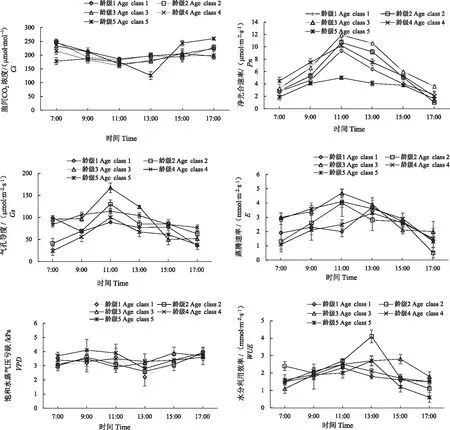
图3 不同龄级荆条光合生理生态因子的日变化规律
由图3知,5种不同龄级荆条Ci、VPD有相似的变化趋势,而Pn、Gs、E和WUE具有相似的变化趋势。不同龄级荆条Ci呈凹形曲线,早晚较高,整体最低值出现在11:00,龄级5最低值出现在13:00。龄级1、2、3、4的Pn均以单峰形式出现,最高值出现在11:00,龄级5净光合速率日变化呈双峰曲线,有明显的“午休”现象,峰值分别出现在11:00和15:00,主峰值为4.1 μmol·m2·s-1,次峰值为3.8 μmol·m2·s-1。且不同龄级间的光合特性有明显不同,净光合速率日变化均值表现为龄级3>龄级2>龄级4>龄级1>龄级5,整体水平上龄级3荆条的Pn最强,处于生长的旺盛年龄阶段。5个龄级荆条Gs最大值都出现在11:00;龄级2、3、4的E最大值出现在11:00,龄级1、5 的E最大值出现在13:00。各龄级荆条VPD呈凹型曲线,最低值均出现在13:00,且变化幅度不大。龄级1、2、4、5荆条WUE最高值出现在13:00,龄级3最高值出现在15:00。
3 讨论与结论
光合作用是植物生长的重要生理过程,植物的净光合速率作为植物叶片的重要功能性状,是衡量植物生产能力以及适应能力的一项重要指标[13],受植物类型、形态、发育阶段等生物因子和光照强度、温度、水分等非生物因子的共同影响[14]。因此,在研究植物生长及固碳能力时,要考虑不同年龄阶段植物对其有机质积累的影响。本研究表明,太行山南麓地区不同龄级荆条在对光的适应和响应上有明显的差异,光合有效辐射是影响植物光合作用最重要的生态因子[15-17],叶片对不同光照强度梯度的响应反映了叶片的光适应性及光能利用效率[18],随着光照强度的变化,不同龄级荆条的净光合速率表现为龄级3在不同光照强度下的光合能力均高于其他龄级,从低龄级到中龄级、光饱和点和光补偿点升高,随着植物的衰老,高龄级荆条的净光合速率、光饱和点和光补偿点降低。 即光照充足的条件下龄级3的光合能力最强,合成有机物迅速;随着荆条龄级的增加,叶片的生理活性降低,叶片逐渐衰老,这与翟洪波等[19]的研究结论一致。龄级1、2荆条处于生长发育初级阶段,叶片与气孔的发育不完全成熟,线粒体和酶活性处于增长阶段;龄级3荆条光合效率高,累计更多干物质,生长更快,叶片和气孔逐渐发育成熟,叶片对CO2的扩散阻止能力下降,叶绿素和蛋白质合成更加活跃,光合磷酸化速率增加,线粒体呼吸下降,因而光合能力增强;龄级4、5荆条进入衰老阶段,各生理因素朝着不利于光合作用的方向发展。
光合作用是复杂的生理过程,叶片的净光合速率与气孔导度、胞间CO2浓度、蒸腾速率、饱和水蒸气压亏缺等因子有关。荆条的光合速率从生长初期到生长旺盛期净光合速率呈增加趋势,到生长后期,随着植株的衰老,净光合速率逐渐下降,测定期间环境因子变化很小,因此,荆条的不同发育阶段,植物本身的性质对光合速率的影响较大。气孔导度是单位时间内单位面积气孔的水汽蒸腾量,气孔导度越大,气孔开启的程度越大,较高的气孔导度有利于叶肉细胞进行气体交换,吸收更多的CO2进行光合作用[20]。本研究表明,龄级3、4荆条的净光合速率与气孔导度、蒸腾速率与水分利用效率呈显著的正相关关系,且龄级3的气孔导度显著高于龄级1、5。这与许大全等[21]提出的光合速率对气孔导度具有反馈调节作用一致。太行山南麓荆条灌丛中龄级3的蒸腾速率显著高于龄级1、2、5。蒸腾作用是植物重要的生理特性,也反映着植物对水分的需求[22]。太行山南麓地区属于干旱半干旱地区,空气湿度相对较小,大气温度升高时使得叶内外蒸气压差增大,蒸腾速率增大,龄级3荆条属于生长旺盛期,对外界环境的变化能更快的做出适应。太行山南麓荆条灌丛中龄级3 的水分利用效率显著高于龄级2、5,总体表现为中龄级的水分利用效率高,低龄级、高龄级荆条水分利用效率低,表明中龄级荆条能更好地适应干旱环境,这与鲍婧婷等[23]对不同林龄柠条的水分利用特征结论不同,差异来源于植物种类对水分利用效率的影响。植物的水分利用效率可以有效地反映植物对水资源的利用程度,可以有效地评估植物的生长适宜性,水分利用效率受到植物的种类以及外界环境等多方面的影响[24]。随着荆条龄级的增长,气孔导度呈现上升又下降的趋势,蒸腾速率也呈现相似的变化规律,但总体比较稳定。低龄级荆条叶片气孔导度相对于高龄级荆条叶片较高,但蒸腾速率相对较低,水分利用效率和净光合速率均高于高龄级荆条。推断因低龄级荆条处于生长初期阶段,有机物的积累需求较高龄级较大,但其叶片组织不及中龄级荆条,其光合能力以及对水分的利用效率较低,高龄级荆条叶片衰老,叶片组织松弛,植株总体生长呈下降趋势,气孔导度有所下降,与光合系统的活性共同影响了高龄级荆条的水分利用效率。
本研究表明,太行山南麓不同龄级荆条光合生理生态指标的日变化存在显著差异。KRAMER等[25]研究表明,植物的光合速率日变化主要有单峰型、双峰型、多变型3种类型。在本研究中,自然条件下太行山南麓龄级1、2、3、4荆条的净光合速率呈明显的单峰形曲线,光合速率最大值均出现在11:00;龄级5荆条的净光合速率各时段较其他龄级低,11:00出现峰值,15:00有小幅度回升,中午净光合速率降低,气孔导度变小,水分利用效率升高,出现“光合午休”现象,其他龄级“午休”现象不明显。可以解释为龄级较高的荆条为了避免叶片水分过度丧失,以及光合器官的完整性采取的一种在不利生长环境下的对策,也表明随着荆条龄级的升高,其光合能力虽然有所降低,但是荆条的抗逆性有所提高。这与王庆伟等[26]的研究结果一致。
综上所述,本研究表明,太行山南麓龄级3荆条的光合能力显著高于其他龄级,处于生长的旺盛期;不同龄级的荆条在日变化方面多呈单峰型;气孔导度以及蒸腾速率对荆条净光合速率影响较大,虽然龄级5荆条叶片在光合能力以及水分利用效率上有所下降、植物的生长速度减缓甚至衰退,但对环境的变化有更高的抗逆性。本试验能够反映出太行山南麓地区荆条这一优势灌丛随龄级变化光合能力的变化趋势。
猜你喜欢
农业与技术(2022年22期)2022-12-02
农业工程学报(2022年3期)2022-04-16
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
节水灌溉(2020年7期)2020-07-15
农业与技术(2020年12期)2020-07-04
诗选刊(2019年9期)2019-11-20
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
学生天地·小学低年级版(2016年9期)2016-05-14
