
IRE1a与miR-346相互调节维持内质网应激强度促进宫颈癌细胞顺铂耐药
2021-09-22 03:12郭军飞赖卫明马从利穆小萍
实用医学杂志 2021年17期
郭军飞 赖卫明 马从利 穆小萍
广东省妇幼保健院检验科(广州510000)
宫颈癌是威胁妇女健康的常见疾病[1-2]。早期宫颈癌治疗的主要方法是手术切除,已转移或复发性宫颈癌的主要治疗方法是顺铂(cisplatin)单独或者联合其他药物化疗,但宫颈癌细胞对顺铂获得性耐受常导致化疗失败[3-4]。化疗常导致肿瘤细胞发生内质网应激(edoplasmic reticulum stress,ERS),研究[5-8]表明ERS 既可介导某些化疗药物对细胞的杀伤,又参与了肿瘤细胞对某些化疗药的耐受。顺铂通过诱导DNA 损伤导致细胞凋亡[4,9],目前ERS 在宫颈癌细胞顺铂耐药中的作用和机制尚未明确。
miRNA 异常表达与多种疾病发生、发展密切相关[11-12]。miRNA 既可调控ERS 过程,其表达水平也可被ERS 调节[13-16]。在宫颈癌细胞中,ERS 调节因子IRE1a 通过下调miR-125b 的表达促进细胞的凋亡[17],有研究[18]发现IRE1a 可以上调miR-346的表达抑制细胞凋亡,目前ERS 条件下IRE1a 在促进或者抑制细胞凋亡转换的机制尚未明确,IRE1a、miR-346 和miR-125b 三者间的关联以及其在宫颈癌细胞顺铂耐药中的作用还不清楚。
本研究发现ERS 调节因子IRE1a 促进miR-346的表达,同时miR-346 反馈抑制IRE1a 的表达间接调控miR-125b 的表达,防止内质网应激强度过大,促进宫颈癌细胞对顺铂耐药,本研究揭示了一种宫颈癌细胞顺铂耐药的机制。
1 材料与方法
1.1 细胞培养和相关试剂本研究中使用到的宫颈癌细胞系均从美国典型培养物保存中心(American type culture collection)购买。采用含10% 胎牛血清和1%青霉素/链霉素的RPMI 1640 培养基进行细胞培养,培养条件为37℃,5%CO2。通过向培养基中加入浓度逐渐增加的顺铂筛选顺铂耐药的宫颈癌细胞。采用LipofectamineTM2000(Invitrogen,Carlsbad,CA)进行细胞转染。通过向细胞培养基中加入终浓度为300 nmol/L 的毒胡萝卜素(thapsigargin,Tg)诱导细胞发生内质网应激。转染细胞所用的质粒等均来源于课题组前期构建。
1.2 RNA 提取以及实时荧光定量PCR 检测用Trizol(Invitrogen,Carlsbad,CA)试剂提取细胞总RNA。采用MMLV 逆转录酶(TAKARA,大连,中国)进行逆转录,并采用iCycler 实时定量PCR 检测系统(Bio-rad)进行目标基因的定量PCR 检测,U6 snRNA 和β-actin 作为内参基因同时被检测。本研究中使用到的引物序列如下,内参基因U6 反转录引物序列:5′-CGCTTCACGAATTTGCGTGTCAT-3′,上游序列:5′-CTCGCTTCGGCAGCACA-3′,下游序列:5′-ACGCTTCACGAATTTGCGTGTC-3′。IRE1a定量PCR 上游引物5′-CTCTTCCAGCCCTACTACTTCCACG-3′,IRE1a 定量PCR 下游引物5′-TGTACGATTCCCTCCTCTCCCTTCC-3′;IRE1a 3′UTR 上游引物:5′ CGGAATTCCTCATATCCATGCCCAATGCACACG 3′,IRE1a 3′UTR 下游引物:5′CCGCTCGAGTTCTCTGGGTGCAAGCAGTCAAGGC 3′;IRE1a 过表达质粒上游引物:5′CGGGATCCATGCCGGCCCGGCGGCTGCTGCTGC 3′,下游引物:5′ GGAATTCTCAGAGGGCGTCTGGAGTCACTGGG 3′。
1.3 细胞凋亡检测细胞进行相应转染和/或药物处理后,将细胞收集,用冰冷的1×PBS 洗细胞两遍后将细胞重悬在结合缓冲液中,加入5 μL FITC-标记的annexin V(Invitrogen,Carlsbad,CA),室温避光放置15 min 后加入5 μL 碘化丙啶,室温避光作用10 min 后,通过流式细胞仪进行检测,所有检测在染色完成后30 min 内完成。
1.4 Western blot细胞进行相应转染和/或药物处理后,将细胞收集,用含0.1% SDS,1% NP-40,1 mmol/L MgCl2and 10 mmol/L Tris,pH 8.0 的裂解液裂解细胞制备蛋白样品。通过SDS-PAGE 胶分离蛋白并转印至经甲醇预处理的PVDF 膜上,用1× TBST 溶液配制的5%脱脂牛奶封闭PVDF 膜1 h,加入相应的一抗,4℃孵育过夜,用1× TBST 溶液洗膜后加入辣根过氧化物酶标记的二抗室温孵育1 h,洗膜后显影曝光。
1.5 统计学方法计量资料数据以均数±标准差表示,组间的差异采用t检验进行比较。P<0.05为差异有统计学意义。
2 结果
2.1 内质网应激调节因子IRE1a 促进miR-346 的表达与亲本细胞相比,顺铂耐药的宫颈癌细胞存在基础水平的ERS(图1A),在顺铂处理条件下顺铂耐药细胞的ERS 强度维持稳定,而亲代细胞ERS 强度明显高于耐药细胞(图1B)。通过基因芯片和实时荧光定量PCR 检测,发现与亲本细胞相比顺铂耐药细胞中多种miRNA 表达发生显著变化,其中miR-346、miR-125b 等表达上调(表达分别上调4.9 倍和1.6 倍),miR-138 和miR-34a 表达下调(相对表达分别是亲代细胞的53%和40%)。通过特异性抑制剂或者通过小干扰RNA(small inhibitory RNA,siRNA)抑制UPR 不同通路的关键因子,发现IRE1a 参与顺铂耐药细胞miR-346 表达水平的上调(图1C、D)。
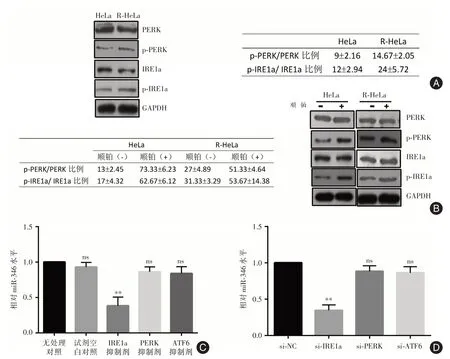
图1 亲本细胞和顺铂耐药细胞ERS 和miRNA 表达情况比较Fig.1 Comparison of ERS state and miRNA expression between parental and cisplatin-resistant cells
2.2 miR-346 参与宫颈癌细胞顺铂耐药的调控在顺铂耐药的宫颈癌细胞中封闭miR-346 可以降低顺铂的半数抑制浓度,促进细胞发生凋亡;而在亲本细胞中过表达miR-346 可以增加顺铂的半数抑制浓度,减少细胞凋亡(图2A-C)。在亲本细胞中过表达miR-346 可以减轻顺铂引起的ERS 强度,而在顺铂耐药细胞中封闭miR-346 导致顺铂引起的ERS 增强(图2D、E)。
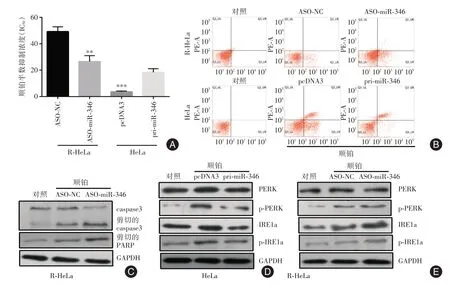
图2 miR-346 调节顺铂引起的ERS 强度和细胞凋亡Fig.2 miR-346 modulates ER strength and cellular apoptosis caused by cisplatin
2.3 miR-346 抑制IRE1a 的表达促进宫颈癌细胞顺铂耐药生物信息学预测提示miR-346 可以调控IRE1a 的表达,双荧光素酶报告实验结果证实miR-346 可以靶定野生型IRE1a 3′ UTR(图3A)。在亲本细胞中过表达外源性miR-346 可以导致IRE1a mRNA 和蛋白水平均降低,而在顺铂耐药细胞中封闭内源性miR-346 则导致IRE1a mRNA和蛋白水平升高(图3B、C)。封闭顺铂耐药细胞内源性miR-346 可降低顺铂的半数抑制浓度,增加顺铂诱导的细胞凋亡;封闭顺铂耐药细胞内源性miR-346 同时通过siRNA 敲降内源性IRE1a 的表达,可以部分挽救封闭内源性miR-346 引起的细胞凋亡(图3D、E)。封闭顺铂耐药细胞内源性miR-346 可导致ERS 强度增加,同时敲降内源性IRE1a可以部分逆转ERS 强度的增加(图3F)。
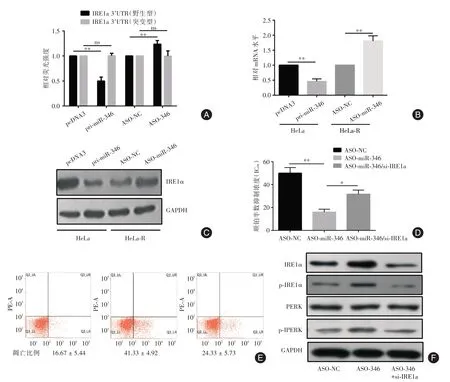
图3 IRE1a 是miR-346 的功能性靶基因Fig.3 IRE1a is functional target of miR-346
2.4 miR-125b 受IRE1a-miR-346 轴调节并参与宫颈癌细胞顺铂耐药在宫颈癌细胞中过表达外源性IRE1a 可显著降低宫颈癌细胞中pre-miR-125b 和miR-12b 的表达水平,而过表达外源性miR-346 可导致宫颈癌细胞中pre-miR-125b 和miR-125b 水平升高(图4A)。在顺铂耐药细胞中封闭miR-346 可导致细胞中pre-miR-125b 和miR-125b水平的降低,而同时敲降IRE1a 的表达部分逆转pre-miR-125b 和miR-125b 水平的降低(图4B)。封闭顺铂耐药细胞内源性miR-346 可导致顺铂半数抑制浓度的降低和细胞凋亡水平的增加,封闭miR-346 同时过表达外源性miR-125b 可以部分逆转封闭内源性miR-346 引起的顺铂半数抑制浓度的降低和细胞凋亡水平的增加(图4C、D)。
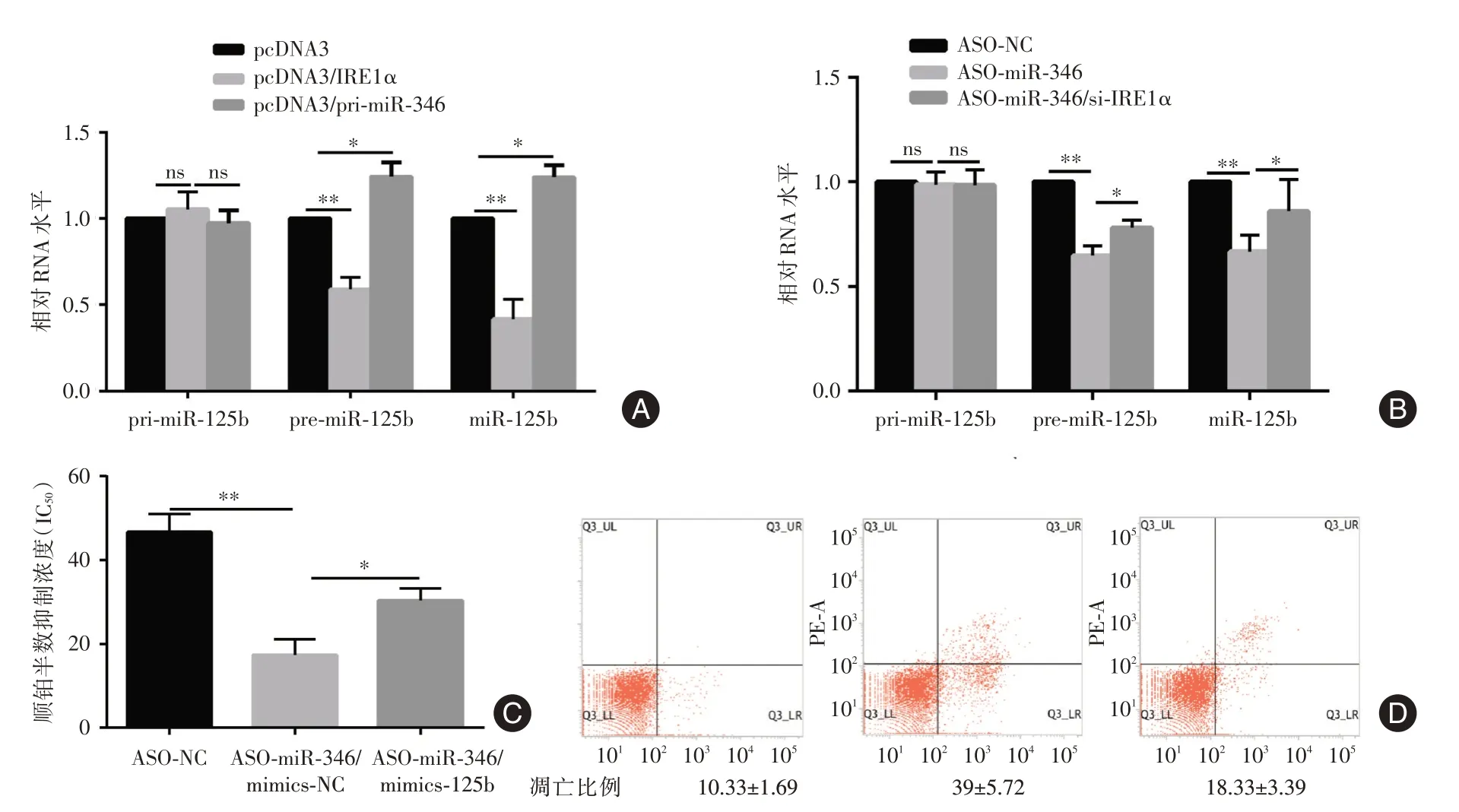
图4 miR-125b 受IRE1a-miR-346 轴调节并参与宫颈癌细胞顺铂耐药Fig.4 miR-125b is regulated by IRE1a-miR-346 axis and involves in cervical cell cisplatin-resistant regualtion
3 讨论
本研究发现相对于顺铂敏感的亲代细胞,顺铂耐药细胞存在基础水平的ERS 避免顺铂引起强度过大的ERS。耐药细胞中miR-346 的表达高于亲本细胞,且miR-346 在宫颈癌顺铂耐药中发挥着重要的功能。当细胞发生ERS 时,UPR 的三条通路将被激活以恢复细胞内环境的稳态,IRE1α、PERK以及ATF6是三条UPR通路的关键分子[19-21]。本研究中,特异性抑制IRE1a 通路可以抑制耐药细胞miR-346 表达并促进顺铂诱导细胞凋亡,提示IRE1a 是ERS 调控宫颈癌细胞顺铂耐药的关键分子。
IRE1a 是内质网上的跨膜蛋白。在轻度ERS条件下,IRE1a 通过剪切XBP1 mRNA 编码区的一个选择性内含子,生成具有超强转录能力的转录因子XBP1(s),促进多种应激基因的表达,使细胞内环境恢复稳态[15];但是长时间的ERS 或者ERS强度过大,IRE1a 导致多种mRNA 降解,细胞内环境无法恢复从而启动细胞凋亡[17]。ERS 条件下IRE1a 活性的调控受多种因素调控,研究表明内质网膜上IRE1a 的分布密度与其激活形式有关,而不同激活状态下IRE1a 对细胞活性的调节存在差异[21]。顺铂耐药细胞中miR-346 可以抑制IRE1a表达,影响内质网膜上IRE1a 的分布密度,继而影响其对细胞活性的调节。
miRNA 是调控基因表达的重要因子,其异常表达与多种疾病相关。miR-346 位于GRID1 基因第二内含子区,其表达独立于其宿主基因,在内质网应激条件下XBP1(s)可以促进miR-346 表达[15]。通过调节不同靶基因的表达,miR-346 在多种肿瘤的发生、发展和肿瘤对治疗的应答中发挥调节作用[22-24]。在宫颈癌细胞中,miR-346 通过调控hTERT 和GSK3β的表达促进宫颈癌细胞在应激条件下存活[18],但是miR-346 是否参与内质网应激本身的调控还未见有报道。本研究发现在宫颈癌细胞中miR-346 可以调控IRE1a 的表达,封闭内源性miR-346 导致IRE1a 的 表 达增加,ERS 强 度 增加,宫颈癌细胞对顺铂的耐药性降低。研究[15]已经证实IRE1a 可以通过XBP1 促进miR-346 的表达,而XBP1 可以反馈抑制IRE1a 的表达[25]。本研究提示miR-346 可能是XBP1 反馈抑制IRE1a 表达的中间分子,在ERS 条件下miR-346 通过负反馈抑制IRE1a 的表达,使ERS 强度维持在适当水平,避免ERS 强度过大引起细胞凋亡。
在持续ERS 存在条件下,IRE1a 核酸内切酶活性增强,其作用的RNA 谱发生变化,可以导致多种miRNA 的表达异常[17]。在ERS 条件下IRE1a 可以降解miR-125b 初始转录本,导致成熟miR-125b水平降低,解除miR-125b 对其靶基因caspase2 表达的抑制,导致细胞的凋亡[17]。在宫颈癌细胞中miR-125b 的表达受到IRE1a 和miR-346 的调控,且miR-125b 可以部分挽救封闭内源性miR-346 引起的耐药细胞顺铂半数抑制浓度的降低和细胞凋亡的增加,提示miR-125b 可能是miR-346 的下游分子。
综上所述,顺铂耐药宫颈癌细胞中基础水平ERS 通过上调miR-346,负反馈抑制ERS 调节因子IRE1a 表达,使得ERS 维持在适当水平,避免IRE1a 过度活化导致miR-125b 异常表达,维持细胞内环境的稳定,促进宫颈癌细胞顺铂耐药。在本次研究中,仅使用了前期实验筛选的宫颈癌顺铂耐药细胞进行实验,因此本研究结论是否具有普遍性还有待进一步验证。此外,本研究都是细胞/分子水平的研究,暂未开展动物实验,因此尚未明确体内是否存在调控关系。后期将在其他不同细胞系开展实验,进行动物实验,收集临床顺铂治疗有效和无效的宫颈癌患者肿瘤组织标本进行验证。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
中国农学通报(2022年12期)2022-06-01
临床肝胆病杂志(2021年7期)2021-12-26
中国种业(2021年11期)2021-11-25
昆明医科大学学报(2021年8期)2021-08-13
现代临床医学(2021年1期)2021-01-26
科学(2020年3期)2020-11-26
心理学报(2020年7期)2020-07-13
现代养生·下半月(2015年6期)2015-09-07
