
基于质谱的分子网络分析化学调控对土曲霉C23-3次生代谢产物及生物活性的影响
2021-09-14 04:36马小翔刘亚月聂影影黎燕媚王远薛欣怡洪鹏志张翼
生物技术通报 2021年8期
马小翔 刘亚月,,3 聂影影 黎燕媚 王远 薛欣怡 洪鹏志,,3 张翼,,3
(1. 广东海洋大学食品科技学院 广东省水产品加工与安全重点实验室 广东省海洋生物制品工程实验室 广东省海洋食品工程技术研究中心 水产品深加工广东普通高等学校重点实验室 湛江市脑健康海洋药物与营养品重点实验室 广东海洋大学海洋药物研究所,湛江 524088; 2. 广东海洋大学深圳研究院海洋医药研发中心,深圳 518120;3. 海洋食品精深加工关键技术省部共建协同创新中心 大连工业大学, 大连 116034)
海洋约占地球总面积的71%,蕴含着种类多样、活性丰富的微生物资源[1]。海洋生物尤其是海洋微生物在海洋高盐、高压、低氧、寡营养的特殊环境下,产生独特的新陈代谢机制,进而能生成结构新颖、活性丰富的次级代谢产物,为药物先导化合物提供丰富来源[2-3]。海洋真菌因遗传背景复杂、代谢产物种类丰富、产量高,逐渐成为海洋微生物新天然产物的主要来源,在2010-2013年的统计里约占64%[4]。而且研究表明真菌含有大量次级代谢产物生物合成基因簇(biosynthetic gene clusters)[5],但在实验室或者人工培养条件下,真菌大部分功能基因簇通常处于沉默状态。通过改变培养基组成、改变发酵条件(如培养容器)以及加入小分子化合物(如前体小分子、酶抑制剂等)等丰富真菌代谢产物多样性的方法越来越受到研究者的关注[6]。
新化合物发现的方法学研究正在引领海洋微生物天然产物研究的新浪潮。基于质谱的分子网络(molecular networking,MN)是一种可视化计算策略[7]。结构相似的化合物在质谱测试中会裂解产生相似的碎片离子,通过计算机算法计算样品经液相色谱-串联质谱分析(liquid chromatography-tandem mass spectrometry,LC-MS/MS)后得到的二级质谱的相似度,依据相似度将质谱信息整合成一种可视化的分子网络图。MN有利于快速、大规模地鉴定已知化合物、相似化合物以及挖掘新化合物。近年来,随着全球天然产物社会分子网络(Global Natural Product Social Molecular Networking,GNPS)数据库的开放使用,借助来自全球的化合物及其谱图积累,GNPS逐渐成为研究天然产物的重要MN工具[8]。如Nie等[9]将生物活性耦合LC-MS/MS和GNPS,高效地评估挖掘来自多种海洋真菌提取物的乙酰胆碱酯酶(acetylcholinesterase,AChE)抑制活性和抗氧化活性化合物。
土曲霉(Aspergillus terreus)属于子囊菌门(Ascomycota)散囊菌纲(Eurotiomycetes)散囊菌目(Eurotiales)发菌科(Trichocomaceae)曲霉属(Aspergillus),是一种广泛存在于海洋和陆地的真菌。丁内酯类和土震素类化合物是土曲霉次级代谢产物中的特征代谢产物[10],研究者发现丁内酯类化合物具有抗肿瘤、抗氧化、抗病毒、抑制葡萄糖苷酶、抑制葡萄糖醛酸酶等活性[11-13],可通过抗炎、抑制细胞周期素依赖蛋白激酶5、促进轴突生长等多种机制实现神经保护作用[14-17];土震素类化合物具有显著的AChE抑制活性[18]。本实验室于前期研究筛选出一株分离自南海珊瑚的海洋土曲霉C23-3,其代谢产物具有一定抗氧化、抑菌和AChE抑制活性且高产丁内酯Ⅰ[19]。考虑到这株菌的代谢产物及活性可能在抗阿尔茨海默氏症(Alzheimer’s disease,AD)等神经退行性疾病药物研发中具有较好研究价值,值得深入挖掘其代谢潜力。
考虑到土曲霉主体活性产物丁内酯I(butyrolactone I)的生源合成途径[20-21],本研究选用小分子前体酪氨酸/4-羟基苯丙酮酸的类似物(3-氨基-L-酪氨酸、3-羟基-L-酪氨酸、3-硝基-L-酪氨酸和3,4-二羟基苯丙酮酸)、异戊烯基转移酶的底物(萜类侧链供体金合欢醇和法尼焦磷酸铵盐(farnesyl pyrophosphate ammonium salt,FPP))、L-硒代蛋氨酸以及氧化胁迫剂(H2O2)和亚硫酸氢钠甲萘醌(menadione sodium bisulfite,MSB)共9种化学诱导剂对菌株 Aspergillus terreus C23-3 进行诱导培养,结合LC-MS/MS和GNPS研究化学调控对菌株次级代谢的影响,以期诱导菌株C23-3产生产量更高或多样性更丰富的丁内酯类等代谢产物,为后续筛选和开发利用具有AD活性的先导化合物奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株 海洋真菌土曲霉Aspergillus terreus C23-3(GDMCC No.60316)采自湛江徐闻珊瑚保护 区,现保存于广东省微生物菌种保藏中心。
1.1.2 培养基 海水马铃薯培养基:马铃薯汁500.0 mL(由马铃薯200.0 g煮熟过滤而成)、蔗糖20.0 g、蛋白胨5.0 g、海水晶20.0 g;固体培养基需加入琼脂20.0 g,加水定容至1.0 L。糙米培养基:糙米833.3 g、海水晶 20.0 g,加水定容至1.0 L。pH均为自然,1×105Pa灭菌20 min备用。
1.1.3 主要试剂与仪器 诱导剂:3-氨基-L-酪氨酸、3-羟基-L-酪氨酸、3-硝基-L-酪氨酸、3,4-二羟基苯丙酮酸、L-硒代蛋氨酸、H2O2、MSB、金合欢醇和FPP。用5%稀盐酸(HCl)或二甲基亚砜(DMSO)或无菌水溶解,于超净台经0.22 μmol/L滤膜过滤备用。
乙酰胆碱酯酶(acetylcholinesterase,AChE)、牛血清白蛋白(bovine serum albumin,BSA)、二苯代苦味酰自由基(1,1-Diphenyl-2-picryl-hydrazyl,DPPH),西格玛奥德里奇(上海)贸易有限公司;碘化硫代乙酰胆碱(acetylthiocholine,ATCh)、5,5'- 二硫代二硝基苯甲酸(dithiobisnitrobenzoic acid,DTNB),合肥博美生物科技有限责任公司;24孔板,无锡耐思生物科技有限公司;薄层层析板Silica gel 60 F254,德国默克公司;LC-MS/MS分析用试剂为Fisher质谱纯,其他试剂及材料均为国产分析纯。
多功能紫外透射仪,上海精科实业有限公司;霉菌培养箱及生物安全柜,上海博讯实业有限公司医疗器械厂;Thermo Finnigan液相色谱-二极管阵列检测器- LCQ Advantage MAX离子阱质谱联用仪,赛默飞世尔科技有限公司;旋转蒸发仪,上海爱朗仪器有限公司;离心浓缩仪,湖南赫西仪器装备有限公司。
1.2 方法
1.2.1 制备种子 将菌株C23-3冻存管置于温度28℃、湿度80%的霉菌培养箱活化过夜;将菌种接种到预先装有200 mL灭菌海水马铃薯固体培养基的锥形培养瓶中培养3-4 d,待菌落孢子生长茂盛备用。
1.2.2 诱导菌株 使用0.85%的无菌生理盐水将1.2.1培养的菌种孢子洗下,摇匀。海水马铃薯培养基和糙米培养基均使用24孔板,每孔加入2 mL培养基和200 μL孢子悬液,28℃、湿度80%培养 3 d。
3 d后,诱导组根据设置的诱导剂浓度(如图1标注所示),向菌株培养物加入化学诱导剂;对照组根据诱导剂的溶剂(5% HCl、DMSO和无菌水),向菌株培养物加入与诱导剂等体积的溶剂;空白组为不加诱导剂或溶剂的菌株培养物。每个条件设置2个复孔,24 孔板置于霉菌培养箱中继续培养,每天观察培养物形态并拍照记录。
1.2.3 提取次生代谢产物 分别在第14天和28天提取菌株次级代谢产物。先将用于微量发酵的24孔板冷冻干燥。对于液体培养物,每孔加入2 mL甲醇浸泡过夜,将孔内物质全部转移至离心管,加入2 mL甲醇洗净孔内残留后转移至离心管合并,功率500 W超声提取30 min,静置过夜取上清液备用;沉淀则加入4 mL甲醇,超声提取步骤同上,重复2次;将总的上清液过滤膜(孔径0.45 μm)浓缩干燥、称重,4℃ 冷藏备用。对于固体培养物,每孔加入1 mL甲醇浸泡过夜;将孔内容物全部转移至离心管后加3 mL甲醇洗净孔内残留,剩余操作同液体培养物的提取。
1.2.4 薄层层析分析、生物活性自显影及化学显色 此部分参考张翼等[22]的方法,不同培养基的发酵提取物均以三氯甲烷∶甲醇(体积比为20∶1)为薄层层析(thin-layer chromatography,TLC)展开剂,分别记录254、365 nm的紫外图像,DPPH自由基清除活性与AChE抑制活性自显影图像,茴香醛硫酸显色及铁氰化钾-三氯化铁显色图像。
1.2.5 LC-MS/MS 将各提取物均用色谱甲醇配成等体积样品溶液进行液质分析。
色谱分析条件:进样量为20 μL,洗脱条件为:0.00-1.00 min,30%的乙腈水溶液(含0.1%甲酸);1.00-8.00 min,30%-99%乙腈水溶液(含0.1%甲酸);8.00-12.00 min,99%的乙腈水溶液(含0.1%甲酸);12.00-12.20 min,99%-30%的乙腈水溶液(含0.1%甲酸);12.20-15.00 min,30%的乙腈水溶液(含0.1%甲酸),流速0.6 mL/min。DAD检测器信号采集波长为190-600 nm,监测波长为210 nm、254 nm、280 nm、310 nm和360 nm。色谱柱为Phenomenex Kinetex C18 100Å反相色谱柱(100×4.60 mm,5 μm)。
质谱检测条件:质量扫描范围设置为m/z 100-2 000,电喷雾离子化,正离子模式,离子源电压:4 kV,毛细管温度:325℃,二级质谱触发信号强度阈值1×105CPS(Count Per Second),碰撞能量:35 eV,离子传输管电压:10 V。
1.2.6 分子网络的创建和可视化分析 使用Proteo Wizard软件转换原始液质数据文件的格式,然后上传至GNPS数据库计算分析,将分析结果导入 Cytoscape 3.7软件绘制分子网络图。利用GNPS的二级质谱相似度匹配功能可注释样品所含的已知化合物,并根据分子网络图中不同节点的关系,结合质谱数据,来发现已知化合物的类似物或衍生物。
2 结果
2.1 诱导前后菌株表观形态变化
菌株C23-3在海水马铃薯液体培养基条件下,如图1-C所示,空白组的菌丝体呈现乳白色,边缘呈现灰黑色;如图1-A所示,分别添加3-氨基-L-酪氨酸、3-羟基-L-酪氨酸、3-硝基-L-酪氨酸后,随着诱导剂浓度的增大,菌株的菌丝体生物量无明显变化,菌丝体由浅黄色变成浅棕色;而添加MSB、H2O2、FPP以及无菌水的组(图1-B-C),菌膜边缘黑色部分增加。
菌株C23-3在糙米培养基条件下,如图1-C所示,空白组菌丝体呈棕色;如图1-A所示,分别添加3-氨基-L-酪氨酸和3-羟基-L-酪氨酸后,实验组各个浓度的菌丝体生物量变少且颜色变浅;加入3-硝基-L-酪氨酸、3,4-二羟基苯丙酮酸、金合欢醇后,菌丝体生物量减少;如图1-B所示,添加L-硒代蛋氨酸后,菌株产生少量白色菌丝体。
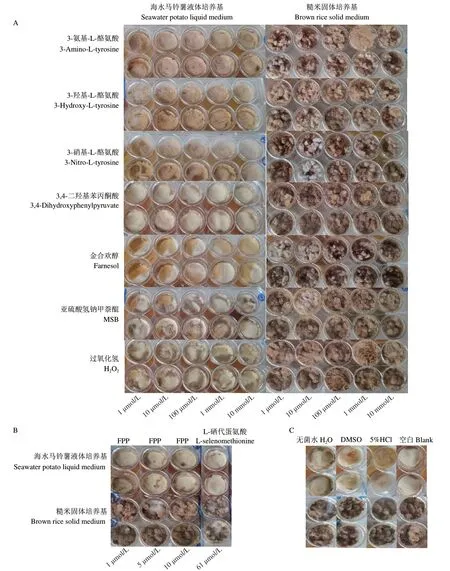
图1 菌株C23-3化学调控前后生长形态(培养28 d)Fig.1 Growth morphology of strain C23-3 before and after chemical regulation(cultured for 28 days)
同等培养条件下,培养14 d的菌株表现形态与培养28 d的无明显差异。综上所述,在3-氨基-L-酪氨酸、3-羟基-L-酪氨酸、3-硝基-L-酪氨酸、3,4-二羟基苯丙酮酸、金合欢醇和L-硒代蛋氨酸诱导前后菌株C23-3外观形态上产生了明显变化,而菌株形态变化与次级代谢产物通常密切相关[23-24],为了验证诱导剂引起的次生代谢变化,本文对菌株提取物进行了TLC指纹图谱与生物活性自显影的分析。
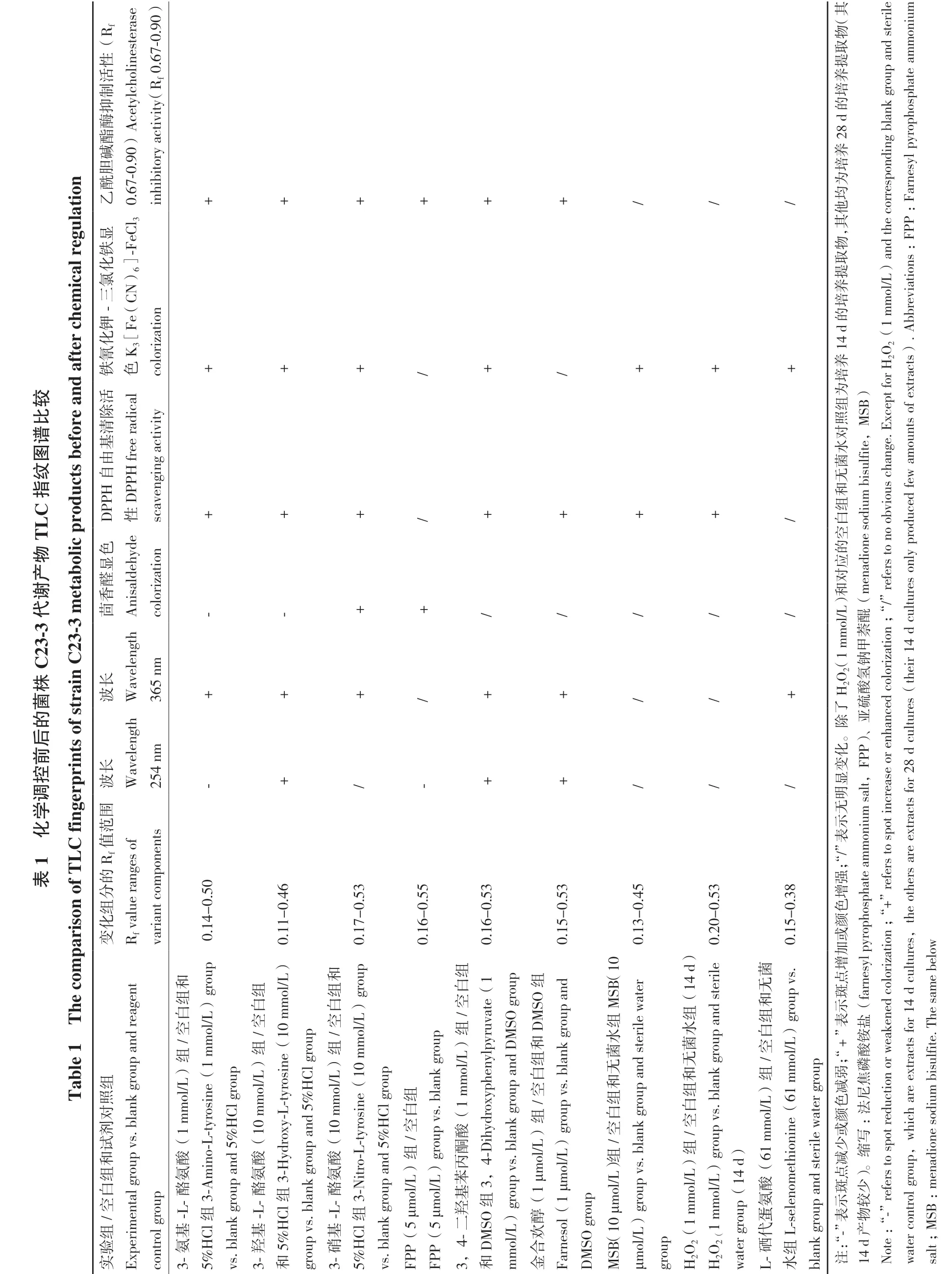
2.2 诱导前后TLC指纹图谱比较
对添加化学诱导剂前后的菌株提取物进行了不同波长下的吸收/荧光斑点、茴香醛显色斑点、DPPH自由基清除活性斑点、三氯化铁-铁氰化钾显色斑点和AChE抑制活性斑点的TLC指纹综合比较,如表1及图2所示,添加化学诱导剂前后菌株TLC指纹图谱发生了显著变化。在糙米培养基中,3-氨基-L-酪氨酸(1 mmol/L)、3-羟基-L-酪氨酸(10 mmol/L)、3-硝基-L-酪氨酸(10 mmol/L)、3,4-二羟基苯丙酮酸(1 mmol/L)、金合欢醇(1 μmol/L)、MSB(10 μmol/L)、H2O2(1 mmol/L)、FPP(5 μmol/L)和L-硒代蛋氨酸(61 mmol/L)这9种诱导条件下代谢产物更丰富或发生变化,且具有一定生物活性。而海水马铃薯液体培养基由于代谢产物过少,未做进一步分析。

图2 菌株 C23-3 化学调控前后发酵产物 TLC 指纹图谱(糙米培养基)Fig.2 TLC fingerprints of strain C23-3 fermentation products before and after chemical regulation(brown rice medium)
2.3 诱导前后代谢产物总体多样性比较
利用LC-MS/MS对糙米培养基上经化学诱导后菌株微量发酵代谢产物进行分析,结合GNPS数据库和Cytoscape等软件做分子网络图。首先使用同一诱导剂的不同条件(改变浓度或培养时间)及其对应空白组和对照组的菌株提取物单独做分子网络分析,然后从9种诱导剂中汇总次生代谢产物丰富或有明显变化的培养条件再做分子网络(图3)。基峰图(base peak chromatograms,BPC)是将每个时间点质谱图中最强离子的离子强度连续描绘而得的图谱,结合分子网络中的节点数量、大小、比例分布等,二者在一定程度上可反映菌株代谢产物的总体多样性。
在分子网络图中,一个节点代表一种化合物(不排除为同分异构体的集合),实质上是液质数据中具有相同整数质荷比、视为相同二级质谱图的所有母离子的集合,其上标注的质荷比为采集的全部母离子质荷比的均值;化合物二级质谱之间的相似性用余弦值(即cosine值,范围为0-1)表示,余弦值与相似性成正比,节点之间的连线粗细与节点之间的cosine值正相关;节点不同颜色的扇形分布代表化合物不同来源的相对含量;节点的大小代表不同化合物的总离子强度。如图3所示,分子网络图直观的显示某些代谢产物在不同的培养条件下分布不同,有的有明显的偏向性,只在少数甚至个别条件下才产生,如最大离子簇中m/z 461.259、459.243、551.168等节点,在Cytoscape软件中点击相应的节点可浏览详细信息。根据样品与GNPS数据库里收录化合物二级质谱的相似性匹配(cosine值大于0.7),判断菌株C23-3的已知产物主要有丁内酯Ⅰ(compound 1)、土震素B(compound 2)、洛伐他汀(compound 3)和和一个相对微量成分洛伐他汀羟酸(compound 4,洛伐他汀内酯键开环产物)。
?
对于这四种特征化合物,根据GNPS提取的化合物的出峰时间和分子量信息,在BPC(图4)中可找到对应的色谱峰及质谱信息(丁内酯Ⅰ:[M+H]+m/z 425,保留时间在8.35 min;土震素B:[M+H]+m/z 527,保留时间8.84 min;洛伐他汀:[M+H]+m/z 427,保留时间10.39 min;洛伐他汀羟酸:[M+H]+m/z 445,保留时间9.43 min;其二级质谱见图5-7),发现在不同培养条件下它们的产量有较大变化。
除了上述已知代谢产物的变化外,实验中也观察到一些未知成分的变化,在图4中用箭头标注了相应样品中较独特的成分。

图4 菌株C23-3不同培养条件下代谢产物的液质联用基峰色谱图Fig.4 Base peak chromatograms of the metabolites of strain C23-3 under different culture conditions
如加入3-氨基-L-酪氨酸(1 mmol/L)后,在12.63 min产生基峰为m/z 736.43的化合物;加入3-羟基-L-酪氨酸(10 mmol/L)后,在13.07 min产生基峰为m/z 338.80的化合物;加入3-硝基-L-酪氨酸(10 mmol/L)后,在7.71 min产生基峰为m/z 513.04的化合物;加入3,4-二羟基苯丙酮酸(1 mmol/L)后,在10.05 min和11.12 min分别产生基峰为m/z 520.30和529.25的化合物;而空白组、试剂对照组以及其他诱导组在相同保留时间则没有,推测这些前体小分子可能对菌株的代谢产物产生影响。
添加FPP(5 μmol/L)后,在7.33 min(基峰m/z 422.99)和11.24 min(基峰m/z 429.25)的化合物强度比其他组略高;添加了金合欢醇的实验组在13.49 min的化合物(基峰m/z 409.24)与其他组有所区别,由此推断萜类侧链供体FPP和金合欢醇可能刺激菌株C23-3提高部分化合物的产量。
添加MSB(10 μmol/L)后,在10.64 min产生基峰为m/z 459.26的化合物;添加H2O2(1 mmol/L)后,在12.84 min产生基峰为m/z 338.21的化合物,推测氧化剂MSB和H2O2对菌株C23-3可能造成氧化胁迫,刺激其代谢产物发生变化。
加入L-硒代蛋氨酸(61 mmol/L)的实验组与其他组的BPC无显著差异,与TLC结果有一定差异,推测可能某些代谢产物在正离子条件下未能检测到。
综上所述,这9种化学诱导剂可对菌株C23-3代谢产物的种类或产量产生不同程度的影响。
2.4 诱导前后特征代谢产物衍生物分析
除了通过与数据库收录的二级质谱匹配以指认已知化合物,GNPS还可利用二级质谱相似性来分析样品中各代谢产物之间的关系,由已知化合物节点来发现其可能的同族化合物。
根据图3中通过对已知化合物Compounds 1-4相连的主要节点的二级质谱(图5-图7)的分析,主要是其母离子和碎裂离子与已知化合物相应离子质荷比的关系,推测这些化合物可能为相应已知化合物异构体或由其脱水、脱氢、脱氧或降碳而来。与质荷比为m/z 425.027的丁内酯Ⅰ结构相似的节点有4个(Compounds 5-8),质荷比依次为m/z 425.028(和丁内酯I二级质谱相似又有显著不同,虽未与其聚在同簇,但推测可能为其异构体)、m/z 423.01(丁内酯I脱氢产物)、m/z 441.046(Compound 5加氧产物)和m/z 407.021(丁内酯I脱水产物);与质荷比为m/z 527.166的土震素B结构相似的节点有2个(Compounds 9-10),质荷比分别为m/z 511.169(土震素B脱氧产物)和m/z 513.128(土震素B降碳产物,如甲氧基变为羟基);与质荷比为m/z 427.236的洛伐他汀结构相似的节点有3个(Compounds 11-13),依次是m/z 409.247(脱水产物)、m/z 411.184(脱氧产物)和m/z 429.256(加氢产物)。其中部分化合物在上文BPC谱图比较中也是独特色谱峰。

图3 菌株C23-3不同培养条件下代谢产物基于二级质谱联系的分子网络图及局部放大图Fig.3 Molecular network and its partial enlarged diagram of the metabolites of strain C23-3 under different cultural conditions based on MS2 relationships

图5 Compound 1及其可能衍生物的二级质谱图Fig. 5 MS2 spectra of compound 1 and its possible derivatives

图7 Compounds 3、4及其可能衍生物的二级质谱图Fig.7 MS2 spectra of compounds 3,4 and their possible derivatives
为了进一步分析9种诱导剂对菌株C23-3丁内酯类、土震素类及洛伐他汀类特征代谢产物多样性及产量的影响,对上述GNPS注释或推测的该三类化合物Compounds 1-13的提取离子色谱图(Extracted ion chromatogram,EIC)相应化合物峰面积进行了计算和比较。

图6 Compound 2及其可能衍生物的二级质谱图Fig.6 MS2 spectra of compound 2 and its possible derivatives
结果(图8)显示,添加诱导剂金合欢醇(1 μmol/L)的丁内酯Ⅰ产量相对最多,FPP(5 μmol/L)以及MSB(10 μmol/L)诱导组代谢产物中的丁内酯Ⅰ略少于金合欢醇组,但FPP组和MSB组的丁内酯类的种类和产量更丰富;FPP(5 μmol/L)诱导组的土震素B产量和种类在这九组里相对较优,其次是3-氨基-L-酪氨酸(1 mmol/L)诱导组。而洛伐他汀类代谢产物在3,4-二羟基苯丙酮酸(1 mmol/L)、3-氨基-L-酪氨酸(1 mmol/L)和硒代蛋氨酸(61 mmol/L)组中产量与多样性较优。综上所述,菌株C23-3在不同化学诱导剂的作用下,菌株的特征产物(丁内酯Ⅰ、土震素B和洛伐他汀)及其可能衍生物的种类和产量发生了较大变化。

图8 菌株C23-3不同化学调控条件下的代谢物中特征产物及其衍生物的含量Fig.8 Contents of the characteristic products and their derivatives in the metabolites of strain C23-3 under different chemically regulated conditions
3 讨论
研究表明,很多微生物虽然具有丰富的次生代谢产物合成基因簇,但在实验室培养条件下相当部分处于沉默状态[25],活性菌株的潜力有待进一步挖掘。前体导向生物合成技术有利于在已知的先导化合物基础上开展结构优化与改造,如黄婷婷[26]向吡啶霉素链霉菌NRRL B-2517的野生型菌株饲喂与天然底物3-羟基吡啶羧酸结构类似的芳香羧酸和氨基酸,分离得到几种新的吡啶霉素结构类似物。本研究考虑了不同基础培养基和多类型化学调控手段(包括化合物骨架的小分子前体类似物、萜类侧链供体类似物、氧化胁迫及硒代氨基酸)的综合运用,以利于发现更好的诱导条件。
实验结果证明不同类型的化学诱导剂可对菌株C23-3代谢产物和生物活性的确能产生不同程度的影响,特别是丁内酯类、土震素类和洛伐他汀类化合物的多样性和产量发生了较大的变化,丁内酯类和土震素类化合物被发现具有一定的抗氧化、神经保护、抗神经炎症和乙酰胆碱酯酶抑制活性[15-18],本研究为抗阿尔茨海默氏症等活性的新化合物发现提供了基础。
另一方面,本研究中不同的调控条件包括一些前体类似物虽然改变了菌株的代谢,但在质谱中并未明显检测到含有特定饲喂前体结构片段的预期丁内酯衍生物,推测可能是由于培养基中原有的天然前体(如酪氨酸、苯丙氨酸、对羟基苯丙酮酸和二甲基烯丙基二磷酸)与相应合成酶系的亲和力更强,故菌株的特征代谢产物丁内酯Ⅰ仍是主产物,可能抑制了目标化合物的产生,实验中发现的代谢产物改变可能是调控物对菌株代谢产生了间接影响。如李亚伟等[27]利用真菌有机相粗提物诱导纤维堆囊菌,发现加入的诱导物对该菌生长和营养利用无显著影响,而可能通过小分子信号物质改变关键酶的活性,调整代谢途径,进而影响目标产物埃博霉素的产量。在今后的研究中通过改变前体类似物的加入时间或加入相应的酶抑制剂,有可能会促进相应衍生物产生,这有待进一步研究。
本研究也显示基于LC-MS/MS的GNPS分子网络对于挖掘菌株多样性次生代谢产物的优点,可对已知化合物进行注释,并基于二级质谱相似性发现多样化的衍生物,比较不同条件下代谢产物差异。但我们也注意到,这一研究工具还有待继续完善,有些情况下仍需要辅以人工判断,但随着数据库中特定类型化合物二级质谱的积累,它的识别、聚类能力也会越来越强。
4 结论
菌株C23-3在不同化学诱导剂作用下,其TLC紫外斑点、化学显色及生物活性斑点对比空白组和对照组有显著变化;利用GNPS和LC-MS/MS分析其代谢产物,发现9种诱导剂可刺激菌株C23-3代谢产物的种类和产量发生不同程度的变化,其中金 合 欢 醇(1 μmol/L)、FPP(5 μmol/L)、MSB(10 μmol/L)和3-氨基-L-酪氨酸(1 mmol/L)等 4 种诱导剂效果较好,可刺激菌株产生多样化的丁内酯类化合物和土震素类化合物。这为抗氧化或乙酰胆碱酯酶抑制活性化合物的深入研究提供基础,对抗AD药物的研发具有一定的科学意义。
猜你喜欢
江苏农业科学(2022年2期)2022-02-15
江西农业学报(2021年1期)2021-02-01
北华大学学报(自然科学版)(2020年5期)2020-10-21
天津中医药(2020年5期)2020-06-01
中成药(2018年12期)2018-12-29
天然产物研究与开发(2018年1期)2018-02-02
中成药(2018年1期)2018-02-02
中成药(2017年4期)2017-05-17
上海医药(2016年23期)2016-12-22
恋爱婚姻家庭·青春(2016年10期)2016-10-10
