
玉米生长素响应因子基因家族全基因组鉴定及表达分析
2021-09-13 07:28曹丽茹张前进郭子宁鲁晓民皇甫柏树王振华
核农学报 2021年9期
曹丽茹 张前进 郭子宁 鲁晓民 张 新 魏 昕 皇甫柏树 王振华
(1河南省农业科学院粮食作物研究所,河南 郑州 450002;2河南省农业科学院科技成果示范推广处,河南 郑州 450002;3焦作市农林科学研究院,河南 焦作 454150)
生长素在植物生长的不同阶段、不同部位及不同生理活动中都发挥着重要的作用,如根的发育、细胞增殖、花器官发育等生物过程。其中生长素响应因子(auxin response factor,ARF)主要通过结合下游基因启动子区域的顺式元件AuxRE (TGTCTC)来调控响应基因的转录水平[1]。目前拟南芥 (Arabidopsis thaliana)[2]、水稻(Oryza sativa)[3]、 杨树(PopulusL.)[4]、玉米(Zea maysL.)[5]、苹果(Malus pumilaMill.)[6]和黄瓜(Cucumis sativusL.)[7]等植物中分别鉴定出了23、25、39、31、29 和18 个ARF 成员,但在动物和微生物中尚未鉴定到ARF 类基因,表明ARF 家族是植物特有的。研究表明,ARFs 蛋白包含3 个结构域,位于NH2端植物特有的DNA 结合域B3(DNAbinding domain,DBD)、中间区域的激活或者抑制结构(Auxin-resp 结构域)和COOH 末端的二聚化结构域(CTD)。B3 结构与顺式元件AuxRE 结合需要其他氨基酸的辅助,并且二者结合与外源生长素的使用无关[8];CTD 结构域较保守,与Aux/IAAs 结构域相似度较高,可产生二聚化区域[9];对于中间区域,若氨基酸序列含有大量的谷氨酰胺、丝氨酸、亮氨酸该区域起激活作用,若这些氨基酸被脯氨酸、甘氨酸等代替该区域起抑制作用[10]。ARF 家族功能方面,研究表明AtARF5 在维管组织发育过程中起重要作用,是生长素重要的转录激活子[11];AtARF5 通过调节DORNROSCHEN(DRN)基因来间接调控植物子叶发育[12];GmARF17主要在侧根中表达,调控大豆[Glycine max(Linn.)Merr.]下胚轴发育[13];OsARF1在水稻的生长营养期和生殖期均起着重要作用,该基因也是生长素调节的早期反应调控因子[14]。
近年来研究表明,不仅脱落酸(abscisic acid,ABA)、乙烯、赤霉素等激素参与植物逆境胁迫过程,生长素也参与这些过程[15-16];生长素合成路径中的核黄素单加氧(YUCCAs)限速酶,其过量表达可以提高拟南芥的抗旱性[17];此外,Xu 等[18]研究表明,茶树(Camellia sinensisL.)CsARFs基因可能参与植物非生物胁迫和激素信号传导路径。玉米作为粮食、饲料和加工原料对保证粮食安全和农民增收至关重要,但每年由于高温、干旱等非生物胁迫造成严重的经济损伤,故挖掘响应逆境胁迫的基因,通过分子育种技术提高玉米的抗逆性对农业生产具有十分重要的现实意义。目前关于玉米ZmARFs基因的非生物胁迫响应报道较少,本研究对鉴定到的36 个玉米ARFs 基因家族进行系统的进化关系、启动子区域的调控元件、多种非生物胁迫和激素诱导下基因的表达模式及组织表达特性进行分析,以期为深入了解玉米ARFs家族成员的遗传进化及在玉米逆境胁迫中的功能提供理论基础,为玉米抗逆育种提供优异的分子资源。
1 材料与方法
1.1 玉米基因组中ARFs 基因的鉴定、染色体位置及蛋白的理化特性分析
通过Plant Transcription Factor Database (http:/ /planttfdb.cbi.pku.edu.cn/)和Pfam (http:/ /pfam.xfam.org/)对玉米、高粱、拟南芥和水稻的ARFs 基因家族成员进行筛选,去除无DNA 结合结构域和激活或抑制结构域的基因。利用Phytozome (https:/ /phytozome.jgi.doe.gov/pz/portal.html) 确定玉米ARFs的染色体位置,并使用MapChart 软件绘制家族基因的染色体位置。利用在线软件ProtParam (https:/ /web.expasy.org/protparam/)和Sopma (https:/ /npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl? page =npsa_sopma.html)预测蛋白的理化性质。
1.2 ARFs 蛋白的复合进化树构建及分类
利用Clustal W 对玉米、高粱、水稻和拟南芥的序列进行比对,再利用MEGA5.0 中的邻近法(neighborjoining,NJ,bootstrap =1 000)构建复合进化树,根据进化树分支的距离,对ARFs 进行分类。
1.3 玉米ARFs 结构域和共线性分析
根据NCBI 的CDD 下载玉米ARFs 蛋白的保守结构域;采用MEME 在线工具分析ARFs 蛋白的保守基序,基序的最大数目设置为10;根据蛋白序列的相似性(大于75%),利用R 语言中的RCircos 软件绘制玉米、高粱、水稻和拟南芥ARFs 的共线性关系图。
1.4 玉米ARFs 启动子区域的顺式元件及GO 功能分析
使用Plant care 在线预测ZmARFs启动子区域(ATG 上游2 000 bp)的顺式调控元件;Ensemble plant下载玉米ARFs基因参与的GO 功能(细胞组分、分子功能和生物过程)。
1.5 玉米ARFs 基因的表达模式分析
以优异自交系郑6541 为试验材料,玉米在Hoagland 营养液中长到三叶期时进行处理:聚乙二醇(polyethylene glycol,PEG)处理(Hoagland 营养液+20% PEG)、 NaCl处理(Hoagland 营养液+200 mmol·L-1NaCl)、 ABA 处 理(Hoagland 营养液+5 μmol·L-1ABA)和高温处理(42℃),分别在0、4、8、12和24 h 取4 个处理的叶片;以玉米自交系B73 为材料,取正常水分下生长的根、茎、叶、雄穗、雌穗、胚和胚乳作为基因组织表达特性分析,三株相同部位混为一个样品,每个部位取3 个生物学重复。
根据ZmARFs基因序列设计特异引物,引物合成在北京华大基因公司完成。使用EasyPure RNA Purification Kit(上海翊圣生物科技有限公司)提取上述样品的总RNA,TansScrip One-Step gDNA Removal and cDNA Synthesis 试剂盒(上海翊圣生物科技有限公司)反转录成cDNA,以玉米18S为内参基因,使用SYBR Premix Ex TaqTM 试剂盒(上海翊圣生物科技有限公司),采用两步法进行实时荧光定量PCR(quantitative real-time PCR,RT-qPCR):95℃预变性30 s,95℃变性15 s,60℃退火30 s,40 个循环。根据2-△△Ct法计算基因的相对表达量。
1.6 玉米ARFs 蛋白的亚细胞定位分析
根据玉米ARFs基因的开放阅读框(open reading frame,ORF)序列设计引物,将克隆到的ZmARFs基因的ORF 序列装载到荧光表达载体pMDC83 上,然后转化至EHA105 农杆菌感受态细胞。将含有阳性克隆的农杆菌采用注射法侵染6 叶期的烟草叶片,2 d 后在共聚焦荧光显微镜下观察注射农杆菌的叶片部位的绿色荧光蛋白的位置。
2 结果与分析
2.1 玉米ZmARFs 基因的鉴定及理化特性分析
通过对ZmARFs基因家族成员进行筛选,最终确定36 个ZmARFs基因,暂将其命名为ZmARF1 ~ZmARF36(表1)。除了B3 和ARF 结构域,ZmARF1、ZmARF3、ZmARF6~ZmARF8 等19 个基因还含有AUX_IAA 结构域。编码ZmARF 蛋白的氨基酸长度范围从273aa(ZmARF9)到1 147aa(ZmARF23),分子量从29.73 kDa(ZmARF9)到127.24 kDa(ZmARF11),等电点从5.41(ZmARF34)到8.77(ZmARF32)。这些蛋白的氨基酸长度和蛋白特性相差均较大,表明ZmARF家族成员特性不同,在生物学进程中发挥不同的作用。二级结构分析显示,这些蛋白α-螺旋所占蛋白序列比例范围是14.35%(ZmARF4)~37.42%(ZmARF36),β-转角所占蛋白序列比例范围是3.16%(ZmARF30)~8.47%(ZmARF1),无规则卷曲所占蛋白序列比例范围是42.36%(ZmARF36)~66.32%(ZmARF29),延伸链的范围是 13.09% ( ZmARF29 ) ~ 22.35%(ZmARF31),表明这些蛋白由α-螺旋、β-转角、无规则卷曲和延伸链4 种结构组成,且无规则卷曲是主要的存在形式。
ZmARFs基因所在的染色体物理位置定位分析显示(图1),36 个基因随机不均匀地分布在玉米的10 条染色体上(除第7 染色体外),分别分布着5、5、6、4、6、3、2、1 和4 个ZmARFs基因。分析发现,ZmARFs基因大多位于染色体两端,仅1 号(ZmARF1~ZmARF5)、3 号(ZmARF12、ZmARF13、ZmARF114)、4 号(ZmARF18 和ZmARF19)和6 号(ZmARF28)染色体上的基因位于染色体中部。此外,1 号染色体上的ZmARF4 和ZmARF5,2号染色体的ZmARF7 和ZmARF8、ZmARF9 和ZmARF10,10 号染色体的ZmARF34 和ZmARF35 所在的物理位点较近。
2.2 不同作物ARFs 蛋白的系统进化分析
对获得的拟南芥(22 个)、水稻(27 个)、高粱(25个)和玉米(36 个)的ARF 蛋白,利用MEGA 的NJ 法构建系统进化树(图2)。整个进化树分为四大类(Ⅰ~Ⅳ),其中Ⅱ类含有较多的同系或旁系同源物,根据不同物种蛋白序列相似程度又细分为IIa、IIb 和IIc。玉米的36 个ARF 蛋白主要聚在第Ⅱ分支(14个)的IIb 上(9 个),其次是第Ⅲ分支(11 个),而IIa亚分支上只含有拟南芥的6 个ARF 蛋白。总体来看,其他分支上均含有4 个物种的ARF 蛋白,并且玉米的ARF 蛋白家族与高粱的ARF 蛋白家族亲缘关系较近,与水稻的亲缘关系次之,与拟南芥的关系最远。此外,IIb 分支上的ZmARF13 和ZmARF30、 ZmARF4 和ZmARF5 聚在一个小分支上,说明ZmARF13 和ZmARF30、ZmARF4 和ZmARF5 蛋白进化关系更近。
2.3 不同作物ARFs 蛋白的共线性分析
蛋白共线性分析是以不同物种基因编码的蛋白序列为对象,根据蛋白序列相似性来探究物种内或物种间存在的同系或旁系同源对,及潜在的进化关系。本研究为了进一步分析拟南芥、水稻、高粱和玉米间的进化关系,对4 个物种进行。如图3 共线性片段分析可知,玉米与高粱直系同源的家族成员有17 个、与水稻直系同源的家族成员有2 个,高粱与水稻直系同源家族成员有4 个,但高粱、玉米、水稻均与拟南芥无直系同源片段。旁系同源关系分析,发现玉米家族成员含有2 对旁系同源,高粱含有1 对、水稻含有1 对、拟南芥有3 对。
2.4 玉米ARFs 基因的顺式作用元件分析
Plant care 在线对ZmARFs基因起始密码子上游2 000 bp 序列进行顺式调控元件分析(表2),发现这些基因除了含有基本的核心元件和光响应元件外,还存在多个顺式元件,激素类如ABA、生长素、茉莉酸甲酯、赤霉素、水杨酸响应元件,非生物胁迫类如干旱、低温、渗透胁迫及厌氧诱导相关的调控元件,除此调控元件外,ZmARF1、ZmARF2 和ZmARF3 启动子区还含有分生组织表达相关的元件。表明玉米在遭遇胁迫时,可以通过结合这些顺式元件来调控ZmARFs基因的表达,从而抵御胁迫。
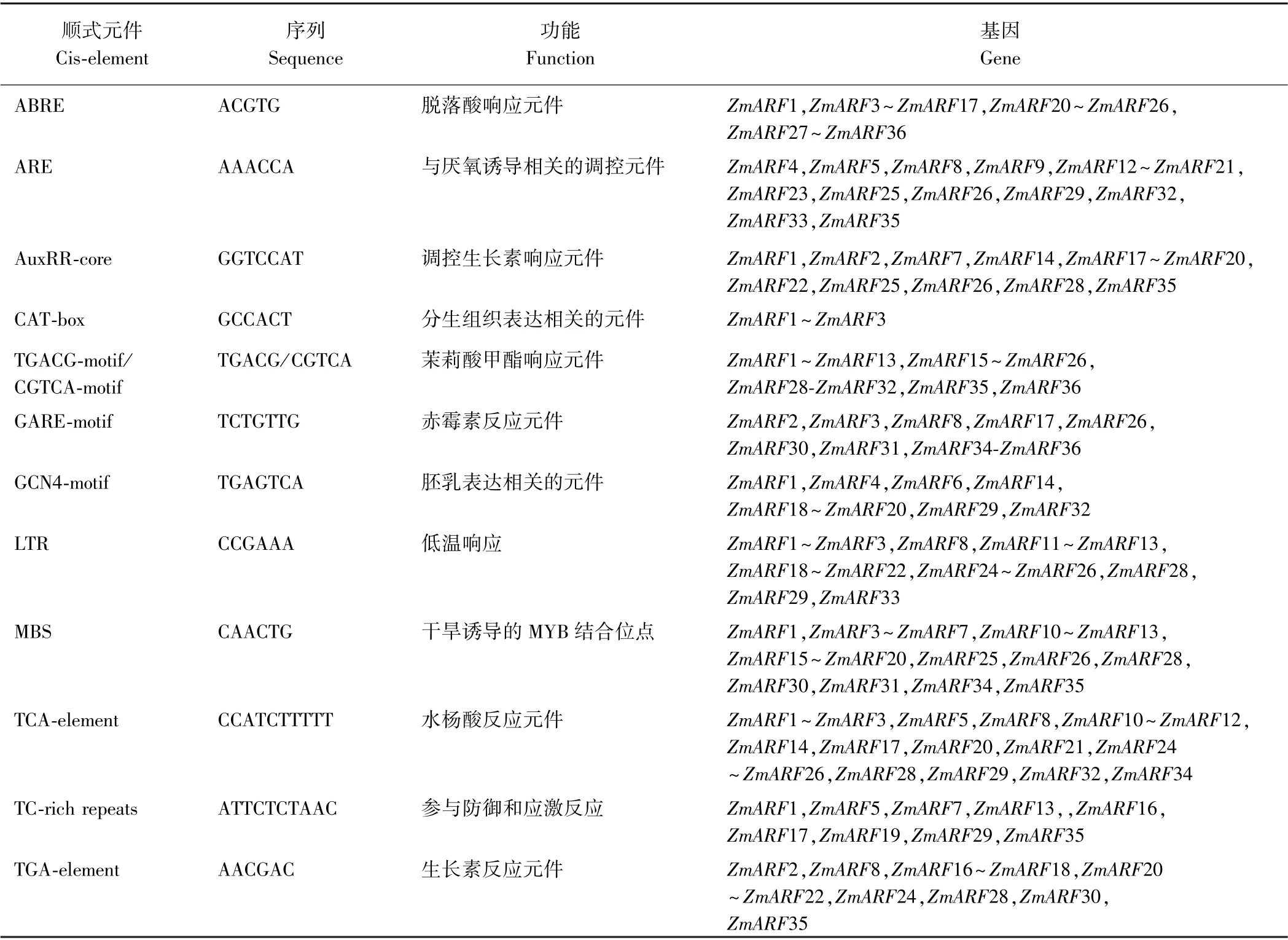
表2 ZmARFs 基因家族启动子区顺式元件Table 2 Cis-element regulatory in prometer of ZmARFs gene family
2.5 玉米ARFs 基因的GO 功能富集分析
对36 个ZmARFs基因进行GO 注释分析(图4),这些基因参与分子生物学功能(GO:0003677-DNA 结合,GO:0005515-蛋白结合,GO:0042802-一致性蛋白结合,GO:0003700-转录因子活性)、生物学过程(GO:0006355-转录调控,GO:0009734-激活生长素信号通路,GO:0010150-叶片衰老,GO:0010158-细胞远轴,GO:0010582-花分生组织,GO:0045892-转录负调控,GO:0051301-细胞分裂,GO:0009908-根发育,GO:0048607-分生组织发育)和细胞学组分(GO:0005634-细胞核)。分析发现ZmARFs基因多富集在GO:0003677-DNA 结合、GO:0005515 -蛋白结合、GO:0006355-转录调控、GO:0009734-激活生长素信号通路、GO:0005634-细胞核等路径上,说明玉米ARFs家族多个成员主要参与转录调控和生长素信号传导路径。此外,发现ZmARF7 还参与根发育和分生组织发育;而ZmARF6 和ZmARF20 基因主要参与叶片衰老、转录负调控和细胞分裂。
2.6 玉米ARFs 基因的组织表达分析
通过分析ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因在根、茎、叶、雌穗、雄穗、花药、胚和胚乳组织中的表达(图5),发现4 个基因均在雄穗中高表达,而在花药中表达量最低,其中差别最大的是ZmARF22(160 倍),差别最小的是ZmARF1(50 倍);4个基因在胚中的表达量均高于在胚乳中的表达量(3.5~22 倍)。此外,ZmARF1 在茎和雌穗中相对表达量也较高,其他3 个基因在根中相对表达量较高。
2.7 玉米ARFs 基因响应非生物和激素诱导的表达分析
由图6 可知,ZmARF1、ZmARF6 和ZmARF13 基因均强烈响应外源ABA 处理,在ABA 处理后的2、4、12和24 h 表达量均显著上调表达,其中ZmARF1 和ZmARF22 在处理2 h 达到最高峰,分别是对照的10.8和5.4 倍;ZmARF6 和ZmARF13 在处理4 h 达到最高峰,分别是对照的15.9 和12.3 倍。高温胁迫下,ZmARF1、ZmARF6 和ZmARF13 在4 个胁迫时间点均显著上调表达,且在胁迫初期2 h 时表达量达到最大值,分别是对照的2.7、4.6 和6.7 倍;而ZmARF22 基因在胁迫后表达量降低。NaCl 胁迫时,除ZmARF22外其他3 个基因的表达量均高于对照,ZmARF6 基因在胁迫2 h 时表达达到最高峰,是对照的2.6 倍;ZmARF1、ZmARF13 和ZmARF22 基因在胁迫12 h 时表达达到最高峰,分别是对照的4.6、7.8 和2.4 倍。PEG 胁迫下,4 个基因的表达均高于对照,达到显著水平,且都在胁迫4 h 达到表达最高峰,分别是对照的11.5、6.4、12.6 和3.2 倍。
2.8 玉米ARFs 蛋白的亚细胞定位分析
为了探究玉米ARFs 蛋白的作用机制,本研究对ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因编码的蛋白进行亚细胞定位,结果如图7 所示。对照35S:GFP 在烟草的细胞膜和细胞质核中均有表达,而35S:ZmARF1-GFP、 35S: ZmARF6-GFP、 35S: ZmARF13-GFP 和35S:ZmARF22-GFP 融合蛋白只在细胞核发出绿色荧光信号,说明ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因编码的蛋白位于细胞核,属于核蛋白,具有转录因子的一般特性。
3 讨论
植物特有的转录因子ARF 已经在多个物种的全基因水平上被鉴定到,其可通过结合下游功能基因启动子上的顺式作用元件来调控基因的表达。本研究通过全基因组水平,以ARF 转录因子特有的DNA 结合结构域B3 和ARF(Auxin_resp),从玉米中鉴定出了36个ARF 家族成员,可能是玉米ARF 发生了基因复制,或鉴定方法不同等原因导致不同物种间家族成员不一致。总体而言,不同物种间ARF 家族成员数目存在差异,这一现象可能与全基因组的复制相关。本研究发现玉米ARFs 蛋白的氨基酸长度、分子量、等电点及α-螺旋、β-转角、无规则卷曲、延伸链均存在很大的差异,可能与家族成员在生长发育过程中的功能机制不同有关。此外,本研究中玉米36 个ARFs家族成员广泛而非均匀分布于玉米9 条染色体上,ZmARFs基因家族未出现基因簇,这与拟南芥和水稻研究结果一致[2-3]。但在玉米1 号染色上的ZmARF4 和ZmARF5,2 号染色染色体的ZmARF7 和ZmARF8、ZmARF9 和ZmARF10,10 号染色体的ZmARF34 和ZmARF35 所在的物理位点较近,可能与基因家族的基因复制有关,这些位点相近的基因或许在功能上较为相似,这一结果为探究玉米ARFs基因家族的作用机制提供了理论参考。
本研究发现,玉米、高粱、水稻和拟南芥的ARF 蛋白成员分为Ⅰ~Ⅳ四大类,四个分支分别包含5、14、11和6 个玉米ARFs 蛋白,说明玉米ARFs 蛋白家族成员间存在较为广泛的多样性,可能与B3、ARF 和AUX_IAA 结构域的保守性有关。此外,玉米ARFs 蛋白家族亲缘关系与高粱最近、水稻次之、拟南芥最远,这可能与玉米和高粱属于单子叶C4植物,水稻为单子叶C3植物,拟南芥为双子叶C3植物有关,同时暗示高粱与玉米可能存在相同的分化,但玉米内又发生了基因复制。基因复制分为基因内复制和基因间复制,是植物进化过程中普遍存在的一种现象,是基因扩张和物种进化的一个重要方式[19-20]。本研究发现旁系基因对ZmARF4 和ZmARF5 位于第1 号染色体,ZmARF13和ZmARF30 分别位于第3 和第8 号染色体,前者属于基因内复制,后者属于基因间复制。结果表明玉米ARFs家族通过基因内复制和基因间复制来增加家族成员的数目,促进玉米的进化,同时暗示不同类型的ARFs基因家族成员存在不同的进化途径和速度。
不同组织和器官的表达模式分析在很大程度上可以预测基因涉及的生命过程和分子功能。本研究发现ZmARF1、ZmARF6、ZmARF13 和ZmARF22 在玉米根、茎、叶、雄穗、雌穗、花粉及果实等不同器官中均表达,属于组成型基因,在番茄和苹果等作物中也获得了相似的结果[21-22]。这4 个基因均在雄穗分支和胚中表达量较高,在花粉中表达量较低,但ZmARF1 在茎中相对表达量较高,其他3 个基因在根中相对表达量较高。说明玉米ARFs基因参与玉米生长发育过程,但在不同器官生长和发育中的作用既存在相似性又具有不同的生物学功能。水稻和拟南芥中多数ARFs基因存在组织特异表达,同时对不同的非生物胁迫选择表达或者不表达[23]。基因启动子顺式调控元件的分析可以预测基因潜在的分子功能和机制,本研究发现玉米ARFs 基因家族成员的启动子区域含有多种响应元件,如干旱、氧化、低温、防御和应激响应元件、生长素、脱落酸、茉莉酸甲酯、赤霉素、水杨酸植物激素类响应元件等,表明ZmARFs基因的表达受多种因素调控,参与植物的逆境胁迫和生长发育等。
非生物胁迫(高温、干旱和盐)和激素诱导(ABA)的结果表明,ZmARF1、ZmARF6、ZmARF13 基因在高温干旱、盐和ABA 诱导时表达模式一致,均显著上调表达。这些基因的启动子区域均含有干旱响应元件、脱落酸响应元件等,启动子顺式调控元件和荧光定量表达模式的分析证实了二者结果的可靠性。其中,ZmARF1 和ZmARF13 基因在干旱、高温、盐胁迫及ABA 诱导时上调表达的幅度最大,推断这2 个基因可能是参与非生物胁迫和激素诱导应答的主要基因,结果为进一步研究抗逆基因提供储备资源。但4 个基因的表达最高峰所在的胁迫点不同,结合ZmARF22 基因在高温胁迫的4 个时间点的表达模式与其他基因的模式相反的结果,推测玉米ARFs基因响应非生物胁迫和激素诱导的表达模式存在差异,且具有阶段性,或许响应机制存在差异。其他物种也有报道ARFs基因参与非生物胁迫的生物学过程[24-27]。
基因表达后翻译成蛋白质,蛋白质发挥生物学功能,因此了解蛋白质的位置有助于研究基因的功能及作用机制[28]。亚细胞定位结果显示ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因编码的蛋白质,定位于细胞核,属于核蛋白,这为进一步研究这4 个基因的作用机制提供了理论参考。关于ZmARF基因家族响应逆境胁迫的机制还需要后续进一步的研究。
4 结论
本研究利用生物信息学方法,在全基因组水平鉴定了玉米36 个ARF 基因家族成员,发现玉米与高粱的亲缘关系较近,且玉米发生了基因内复制和基因间复制。6 个ZmARFs基因启动子区域含有大量的响应非生物胁迫及激素类元件,主要参与转录调控和生长素信号传导路径。表达模式分析发现ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因均在雄穗分支和胚中表达量较高,在花粉中表达量较低。ZmARF1、ZmARF6、ZmARF13 和ZmARF22 基因表达量受干旱、盐和ABA 诱导,且4 个基因编码的蛋白质属于核蛋白,为后续机制研究奠定了基础。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
中国蔬菜(2021年3期)2021-12-11
小天使·六年级语数英综合(2020年3期)2020-12-15
实用肿瘤学杂志(2020年4期)2020-12-08
三农资讯半月报(2020年14期)2020-08-09
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
