
转录因子NRF3结构、功能及其表达调控的研究进展*
2021-09-01 08:30吴宇财钱金钦何宇辉李学松巩艳青
中国病理生理杂志 2021年8期
吴宇财, 钱金钦, 何宇辉, 李学松, 巩艳青
[北京大学第一医院泌尿外科,北京大学泌尿外科研究所,国家泌尿男生殖系肿瘤研究中心,泌尿生殖系疾病(男)分子诊治北京市重点实验室,北京100034]
“ 帽 领 ”(Cap'n'collar,CNC)家 族 包 括 果 蝇CNC1,线虫Skn-1,脊椎动物核因子E2样蛋白1(nu⁃clear factor E2-like protein 1,NFE2L1)/核因子E2相关因子1(nuclear factor E2-related factor 1,NRF1)、NFE2L2/NRF2和NFE2L3/NRF3,以及相似性较低的BACH1和BACH2蛋白[1]。在信号传递过程中,CNC因子与小Maf家族蛋白形成异源二聚体,以进一步识别并结合多种反应元件如NFE2、Maf识别元件(Maf recognition element,MARE)、抗氧化反应元件(antioxidant response element,ARE)、应激反应元件(stress response element,STRE)和亲电子反应元件(electrophile response element,EpRE),进而调控靶基因的表达[2]。其中研究最为广泛的是NRF2,其在细胞氧化应激、合成代谢以及肿瘤预防与治疗等方面发挥重要作用[3-4]。NRF3作为NRF2的同源基因,其蛋白的功能和调控目前还未得到深入的阐明;明确NRF3表达的调控因子及其调控的靶基因,寻找调控其转录因子活性的通路分子,对于全面了解它的功能和调节机制至关重要。
1 NRF3蛋白结构和表达
脊椎动物NRF转录因子家族由3个密切相关的因子NRF1、NRF2和NRF3组成,三者的结构见图1。它们在C端区域都具有CNC结构域和碱性亮氨酸拉链(basic leucine zipper,bZIP)结构域,中间含有1~2个调节转录激活的酸性区域(AD1、AD2或AD2L);NRF1和NRF3的N末端结构域(N-terminal domain,NTD)带有跨膜区域,可以将蛋白锚定到内质网膜上,此外两者还都包含一个富含Asn/Ser/Thr(NST)的可发生N-糖基化的结构域[5]。不同物种之间的生物信息学分析表明,NRF3结构中的NTD、CNC和bZIP等重要结构域在进化上具有高度保守性。人类NRF3基因位于染色体7p14~15,编码的蛋白质分子量为73 kD,包含7个结构域,即NTD、PEST、TAD、Neh6L、CNC、bZIP和Neh3L,其中NTD区可进一步细分为N末端同源盒1(N-terminal homology box 1,NHB1)和NHB2,两者对NRF3的翻译后修饰起重要作用[1,6]。NRF3通过NHB1信号序列靶向输送到内质网中进行糖基化修饰,NHB2可能参与控制NRF3活性以及其在内质网内的翻译后加工过程[7]。分子生物学研究表明,NRF3有A、B、C三种形式:B形式主要存在于细胞质中,可被靶向运输到内质网并进行N-糖基化修饰而转化为A形式;而C形式主要存在于细胞核中,与sMaf蛋白形成异二聚体,通过与特异性DNA结合位点结合而激活靶基因的转录[1]。Nouhi等[8]的研究表明,C形式可能是NRF3的截短形式,因其N端缺乏NHB1结构域。
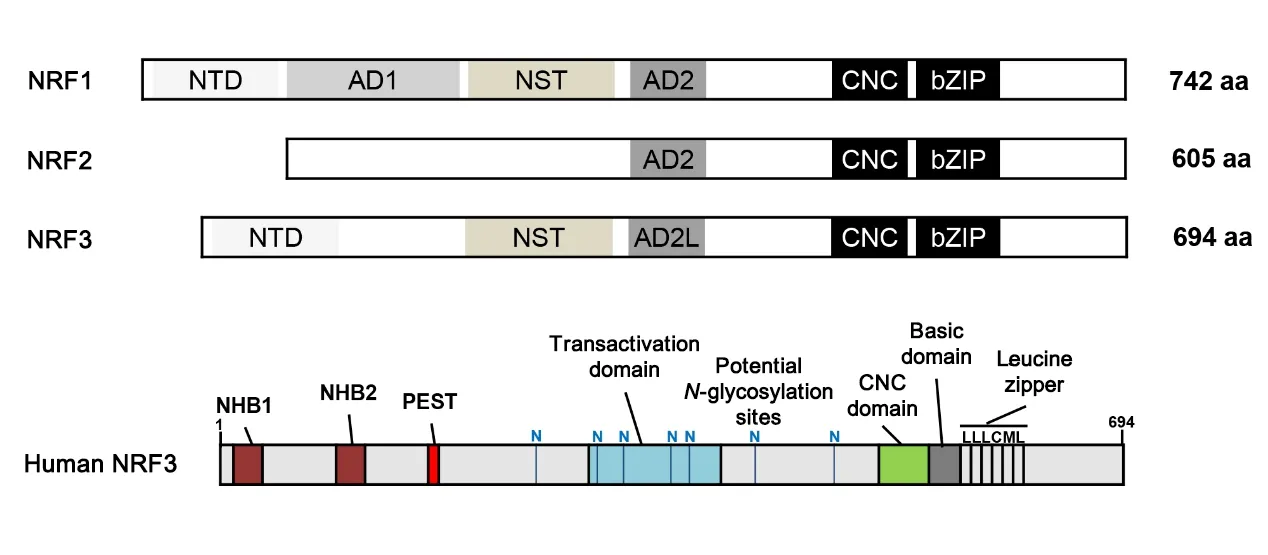
Figure 1.Schematic structures of NRF1,NRF2 and NRF3.These proteins share Cap'n'collar(CNC)and a basic leucine zipper(bZIP)domains in their C-terminus region.They contain one or two acidic domains(AD1,AD2 or AD2L)that regulate the transcriptional activation.The N-terminal domain(NTD)of NRF1 and NRF3 has a transmembrane region that can an⁃chor proteins to the endoplasmic reticulum membrane.In addition,both of them contain an Asn/Ser/Thr-rich(NST)do⁃main,which isa target for N-glycosylation.图1 NRF1、NRF2和NRF3的结构示意图
NRF3在组织和细胞中的表达水平已有研究。人类NRF3转录本从妊娠第12周至足月期间存在于胎盘绒毛中,特别是在滋养层细胞中高度表达,但在成纤维细胞中不表达。与组织中一致,NRF3转录本和蛋白在来源于人胎盘滋养层肿瘤的BeWo和JAR细胞系中高表达。细胞因子TNF-α能促进胎盘细胞中NRF3的mRNA和蛋白表达,提示NRF3可能参与TNF-α介导的信号通路[9]。NRF3在人其他组织中的表达程度不同,如在心脏、脑、肺、肾脏、胰腺、结肠、胸腺和脾脏有表达,而在睾丸、前列腺、骨骼肌和卵巢内几乎不表达。
2 NRF3的表达调节机制
2.1 NRF3的翻译后修饰 在生理条件下NRF3表达受到严格的调控,其在细胞内主要是通过泛素-蛋白酶体途径降解,以维持稳定的水平,此过程可能涉及多个信号分子参与[8]。Kannan等[10]的研究首次提出含F框/WD重复域蛋白7(F-box/WD repeat-con⁃taining protein 7,FBW7)和糖原合成酶激酶3(glyco⁃gen synthase kinase 3,GSK3)共同介导NRF3的降解。FBW7基因编码的蛋白属于F-box蛋白家族的一员,为SCF型泛素连接酶识别目标蛋白的关键组分,可介导包括癌蛋白在内的许多重要蛋白质的泛素化,如cyclin E、c-Myc、Mcl-1、mTOR等,进而经26S蛋白酶体降解[10]。GSK3是一种丝氨酸蛋白激酶,可通过磷酸化FBW7的结合位点而增加FBW7与其底物之间的亲和力。此外,有学者提出NRF3在细胞质和细胞核中通过不同的途径降解[11]。生理条件下NRF3在胞质内是通过羟甲基戊二酰辅酶A还原酶降解蛋白1(3-hydroxy-3-methylglutaryl-coenzyme A reductase degra⁃dation protein 1,HRD1;定位于内质网膜上的泛素连接酶,介导内质网中底物的跨膜转位[12])和含缬酪肽蛋白(valosin-containing protein,VCP;也称为p97,是一种高度保守的ATP酶,通过水解ATP将多肽从膜上分离出来)进行降解,在细胞核中则通过含β-转导蛋白重复域蛋白(β-transducin repeat-containing pro⁃tein,β-TrCP;F-box蛋白家族的成员,是SCF泛素连接酶E3的关键组分[13])促进其泛素化降解[11]。
2.2 NRF3的核转位 研究表明,NRF2对于机体抵抗氧化应激具有重要作用,生理情况下细胞质中的NRF2不断被Cul3-Keap1泛素连接酶复合体泛素化,并经蛋白酶体途径迅速降解,氧化应激会抑制NRF2在胞质中的降解,促使未降解的NRF2转位进入细胞核结合ARE而调控靶基因的表达[14-15]。当抑制NRF3胞质降解关键基因HRD1和VCP的表达时,NRF3在细胞质积聚,但细胞核内NRF3的含量却未见增加,显示NRF3入核与其细胞质内泛素化降解无关,其核转位机制不同于NRF2[11]。有研究表明,天冬氨酰蛋白酶DDI2(DNA-damage inducible 1 homolog 2)的N末端含有泛素样(ubiquitin-like,UBL)结构域,能以较弱的亲和力结合泛素,已知人体内DDI2的UBL结构域参与NRF1核转位前的加工,DDI2的表达下调抑制了NRF1的核转位,并伴有胞质内NRF1的增加[5,16-17]。Chowdhury等[11]证实NRF3进入细胞核内的过程与此相似,DDI2识别多聚泛素化的NRF3并对其N末端NHB2结构域中的AWLVH基序进行切割,从而促进NRF3的核转位;而对于NRF3哪些位点的泛素化介导了DDI2的识别切割,目前未有报道。
2.3 NRF3调控的靶基因 鉴定NRF3调控的靶基因有助于深入了解其在生物体内所发挥的功能。在氧化应激环境下,NRF2进入细胞核内与ARE相互作用,启动下游编码抗氧化蛋白和II相解毒酶的基因表达,发挥细胞保护功能[18]。尽管NRF3的生物学特征在很多方面与NRF2类似,但研究发现其调控作用与NRF2相反:体外细胞实验的结果表明,在氧化剂叔丁基对苯二酚(tert-butylhydroquinone,t-BHQ)的诱导下,NRF3负性调控其下游基因NQO1的表达,使细胞难以抵抗氧化应激损伤[19]。此外,NRF3还可通过结合Prdx6启动子区域内的ARE,进而实现对Prdx6基因的负性调控[20]。有学者初步分析了NRF1、NRF2和NRF3所调控基因的异同,发现了31个共同调控的基因,其中27个基因的调控机制相似;同时也发现了22个仅由NRF3调控的基因[21]。葡萄糖转运蛋白1(glucose transporter 1,GLUT1;由SLC2A1基因编码)受NRF3的正向调控,这可能与癌细胞的代谢重编程过程相关[22]。尽管有报道称UHMK1(U2AF homology motif kinase 1)是NRF3的靶基因[11],但是UHMK1上不含有功能的ARE位点,故其可能是被NRF3间接调节[23]。
近来在干细胞分化研究领域发现,NRF3可通过直接结合到平滑肌细胞特异性基因启动子区域内的CArG盒,来调节平滑肌细胞特异性基因SMαA(smooth muscleα-actin)和SM22α(smooth muscle pro⁃tein 22α)的表达[24]。体外实验研究表明,NRF3还可以直接与Pla2g7(phospholipase A2group VII)基因启动子结合,激活Pla2g7基因表达,进而促进干细胞向平滑肌细胞分化[25]。
此外,有学者认为NRF3可以负调控NRF2的表达,NRF2启动子的近端区域有一个近似于ARE的功能序列,NRF3通过结合此序列来调控NRF2基因的表达[26-27]。目前对于NRF3调控的靶基因尚不十分明确,这也是NRF3的功能复杂、多样,且不同于NRF2的原因之一。
3 NRF3的生物学功能
3.1 NRF3在肿瘤发生发展中的作用 人类癌症基因组计划已经将NRF3归入127个显著突变基因谱中,认为NRF3其能显著诱导癌症的发生[28]。基因芯片数据分析发现NRF3在霍奇金淋巴瘤和套细胞淋巴瘤等多种淋巴肿瘤中高表达[29-30]。除血液系统恶性肿瘤外,NRF3在结直肠癌、乳腺癌、甲状腺癌、肝癌和睾丸癌等肿瘤组织中均有报道出现高表达[31-36]。NRF3的表达量可能与某些肿瘤的预后相关。Wang等[37]的一项回顾性研究表明,与癌旁组织相比,胰腺癌组织中NRF3的mRNA和蛋白表达量均显著增加,NRF3表达量高的患者更易发生淋巴结转移,且肿瘤TNM分期更高,预后更差;多变量分析表明NRF3和肿瘤病理分级可作为胰腺癌患者总体生存率的独立预测因子。近来有研究显示NRF2可通过结合ARE诱导多种抗氧化蛋白和II相解毒酶的合成,从而增强尿路上皮癌细胞对顺铂的耐药性[38]。ARE也是NRF3的结合位点,关于NRF3是否在肿瘤患者发生化疗耐药过程中起作用目前则尚未有相关报道。
NRF3在不同肿瘤发生中的作用有所差别。例如,NRF3表达与肝癌的肿瘤分级和病理T分期呈显著正相关,敲减NRF3可以抑制肝癌细胞的侵袭和转移[36]。然而Chevillard等[39]的动物实验通过管饲法让小鼠暴露在化学致癌物苯并芘下,与野生型小鼠相比,NRF3缺失型小鼠对苯并芘更加敏感,30周后死亡率显著增加(死亡率分别是6%和32%),组织病理学分析显示NRF3缺失型小鼠更易患T淋巴母细胞性淋巴瘤(67%),这个结果表明NRF3能抑制某些血液系统恶性肿瘤的发生。Sun等[33]观察到NRF3的表达在乳腺癌中被显著抑制,并与肿瘤的转移与分期呈现负相关;过表达NRF3能显著抑制肿瘤恶性增殖能力并延缓上皮-间充质转化过程。Zhou等[40]发现NRF3在结直肠癌中处于低表达状态,且NRF3的低表达与更差的预后显著相关;过表达NRF3后结直肠癌细胞的增殖、转移和侵袭能力均显著降低。因此,NRF3对肿瘤发生(促进或抑制)的作用可能与肿瘤的类型及肿瘤所处的微环境有关。
目前NRF3在肿瘤发生过程中的确切机制尚未完全阐明,此过程可能涉及多种信号通路参与。UHMK1是一种丝氨酸/苏氨酸蛋白激酶,在有丝分裂原的激活下其表达上调并使细胞周期蛋白依赖性激酶抑制剂p27Kip1磷酸化,磷酸化的p27Kip1从核内迁移到细胞质中,使细胞周期正常进行,UHMK1活性异常增高能促进肿瘤的发展[41]。Chowdhury等[11]通过敲减NRF3后观察到UHMK1的mRNA和蛋白表达降低,证实了NRF3通过诱导UHMK1的表达来促进结肠癌细胞增殖。此外,NRF3可降低下游抗氧化基因的表达,使细胞代谢过程产生的活性氧增加,继而改变和破坏细胞内生物大分子结构,从而促进肿瘤发生[42]。NRF3还可以通过调节细胞内血管内皮生长因子A(vascular endothelial growth factor A,VEGFA;肿瘤血管生成的关键因子)的水平来促进肿瘤进展[37]。Aono等[22]则提出了β-catenin/T细胞因子(T-cell factor,TCF)-NRF3轴促进肿瘤生成,其中APC(adenomatous polyposis coli)基因缺失是重要的第一步,其缺失导致β-catenin及TCF4介导的相关基因的激活及表达,如NRF3,进而上调GLUT1基因表达水平和促进肿瘤细胞增殖。Sun等[33]探究了NRF3抗乳腺癌的潜在机制,结果表明NRF3通过抑制AKT的磷酸化继而影响ID3(inhibitor of DNA binding 3,HLH protein)的表达,从而发挥抗癌作用。Zhou等[40]在结直肠癌中观察到NRF3的缺失促进表皮生长因子受体(epidermal growth factor receptor,EGFR)和p38的磷酸化激活及表达增加,同时促进EGFR和p38下游分子(AKT、ATF2、CHOP等)的表达,进而增强细胞的增殖能力,促进结直肠癌发生发展。Waku等[43]观察到肿瘤对NRF3的依赖性,并提出了NRF3-蛋白酶体成熟蛋白(proteasome maturation protein,POMP)-20S蛋白酶体轴介导的非泛素依赖蛋白质降解;NRF3通过诱导POMP分子伴侣的表达,激活非泛素依赖的20S蛋白酶体装配,降解肿瘤抑制因子p53和Rb,从而促进肿瘤的生长及恶性进展。
3.2 NRF3抑制炎症的发生 已有研究表明NRF3可能与炎症发生有关。Kitaya等[44]在研究人子宫微血管内皮细胞内干扰素γ(interferon-γ,IFN-γ)调节的靶基因时发现IFN-γ处理能上调NRF3基因的表达,发挥抗炎作用;动物实验研究也表明NRF3能抑制炎症的发生。有研究用酚类抗氧化剂丁羟甲苯处理野生型和NRF3缺陷型小鼠,可引起小鼠广泛肺泡损伤和血管周围炎症细胞浸润,主要表现为呼吸困难和体重下降;与野生型小鼠相比,NRF3缺陷型小鼠体重下降更加显著,且处理后的野生型小鼠肺组织中NRF3的表达量降低[45]。Braun等[46]研究了NRF2敲除小鼠的创伤修复过程,发现在创伤修复早期,参与创面愈合的各种关键分子的表达明显减少,修复后期炎症持续时间延长,然而NRF2敲除小鼠并没有表现出明显的组织学异常;与野生型小鼠相比,NRF2敲除小鼠在受伤前及伤后1 d的皮肤中NRF3的mRNA表达上调,其可能是NRF2无表达的代偿机制,且NRF2和NRF3均可在角质细胞生长因子(ke⁃ratinocyte growth factor,KGF)的作用下表达上调。
3.3 NRF3促进干细胞分化 近年来的研究还发现NRF3在胚胎干细胞向平滑肌细胞分化过程中起重要作用[24]。在平滑肌细胞分化过程中NRF3的mRNA和蛋白表达上调,到分化第4天时达到高峰,在干细胞分化过程中NRF3易位到细胞核内,通过募集SRF/心肌素复合物,或直接结合平滑肌特异性基因的启动子区域,启动α-SMA和SM22α基因的表达[24]。此外,NRF3通过增加NADPH氧化酶4(NADPH oxidase 4,NOX4)、下调抗氧化酶的表达,使超氧化物的产生增加,促进平滑肌分化[24]。因此,NRF3是干细胞分化的重要调节器。Xiao等[25]在探索Pla2g7与干细胞分化的关系时,发现NRF3是Pla2g7的上游调控基因,NRF3被内质网应激激活后,通过转录激活调节Pla2g7基因表达,使Pla2g7表达上调,产生超氧化物,进而介导血清应答因子(serum response factor,SRF)与平滑肌特异性基因启动子的结合,促进平滑肌特异性基因表达。目前,NRF3在干细胞分化中的调控机制尚未完全明确,现有的研究大多来自体外实验,需要后续更多的体内研究进行验证。
4 结语与展望
NRF3是一种受严格调控和转录翻译后修饰的转录因子。糖基化修饰是其重要的修饰方式,但NRF3蛋白是否存在其他转录后修饰对其功能的调节,如磷酸化、类泛素化和棕榈酰化修饰等尚待研究。NRF3作为干性标志基因在干细胞分化早期上调,但是其具体作用尚未明确。虽然有研究报道了NRF3可能调控的靶基因,如Prdx6、Nqo1、α-SMA、SM22α、NOX4等,但仍缺乏它们体内相关性的足够证据。基因表达谱芯片研究显示NRF3转录本在人类多种癌症中表达上调,但仅反映了其mRNA表达水平;对这些组织中诱导表达的转录本是编码有功能还是突变的转录因子,以及亚细胞内调控和在肿瘤发生中的具体作用机制尚未十分明确;NRF3及其相关通路将可能成为癌症患者治疗的新的潜在靶点。作为一直被忽视的CNC家族转录因子,NRF3的研究正逐步进入公众视野,因此明确NRF3表达的调控因子及其调控的靶基因,以及调控其转录因子活性的相关通路和机制,对于全面了解其在人类生理活动或疾病中的功能和调节具有重要研究意义。
猜你喜欢
医学综述(2020年11期)2020-02-16
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
广州大学学报(自然科学版)(2019年1期)2019-05-07
作文教学研究(2016年1期)2016-07-05
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
医学研究杂志(2015年8期)2015-06-22
中国医学科学院学报(2015年5期)2015-03-01
