
酶预处理对木质纤维素纳米纤丝性能的影响
2021-08-13 09:51贺礼龙,黄璐瑶,高文花,曾劲松,陈克复
大连工业大学学报 2021年4期
贺 礼 龙, 黄 璐 瑶, 高 文 花, 曾 劲 松, 陈 克 复
( 华南理工大学 制浆造纸工程国家重点实验室, 广东 广州 510640 )
0 引 言
纳米纤维素纤丝(CNF)是通过解构植物纤维素纤维而获得的纳米级材料。CNF长度为几百纳米到几微米,直径为5~100 nm。CNF具有高比表面积、高机械强度、低密度、可生物降解性和生物相容性等优越性能[1],已成为纤维素科学研究领域的热点。
通常在生产CNF之前,需去除原材料中的木质素,这必将导致大量的能量和化学品消耗,对环境的影响比较大。相对于纳米纤维素纤丝,木质纤维素纳米纤丝(LCNF)制备成本低,过程污染小[2],其制备方法与CNF几乎一致。目前,有许多方法可用于生产CNF,如机械精制、冷冻破碎、超声处理和微波处理等,其中机械精制工艺已普遍应用于生产,如高压均质化、微流化和研磨化。但是,精制过程中的高能量输入以及精制后产品的异质性强烈限制了其应用[3-4]。为了克服这些挑战,在机械精制之前,可采用化学/生物预处理来“打开/松散/软化”纤维素原材料。尽管各种基于化学的预处理方法都可以实现这一目标,如TEMPO氧化、羧甲基化、磺化等,但是这些化学过程会产生一定的污染物,具有相当大的环境风险,限制了产品的进一步规模化推广应用[5-6]。与化学预处理方法相比,生物处理因其酶负荷低、酶选择性高、反应条件温和而被认为是更具吸引力的方法。
本研究采用内切葡聚糖酶和木聚糖酶进行预处理来制备LCNF,分析了预处理前后组分的变化,并探讨其对LCNF保水值、结晶度、比表面积等性能的影响,以期为高性能LCNF的绿色低能耗制备提供理论基础。
1 实 验
1.1 材料与仪器
材料:漂白化学热磨机械浆(BCTMP),芬欧汇川(常熟)纸业有限公司,游离度300 mL,白度79.0% ISO;商业内切葡聚糖酶(Banzyme 2900),芬欧汇川有限公司;木聚糖酶,麦克林生化科技有限公司;内切葡聚糖酶和木聚糖酶的酶活性分别为7.3和327 IU/g。亚氯酸钠、冰醋酸、丙酮;分析纯。
仪器:MKCA6-2J超微粒研磨仪,日本增幸产业株式会社;DionexICS-5000离子色谱仪,赛默飞世尔科技(中国)有限公司;Multimode-8原子力显微镜,德国Bruker公司;UV-1900紫外分光光度计,日本岛津仪器有限公司;PCD-03粒子电荷探测仪,德国Mütek公司。
1.2 实验方法
1.2.1 原料预处理
将纸浆浸泡在去离子水中12 h,再采用实验室规模的Valley打浆机对纤维进行疏解处理。处理后的纸浆离心脱水并置于4 ℃平衡水分。
1.2.2 浆料脱木质素处理
浆料木质素脱除采用改进的测定综纤维素含量的方法(GB/T 2677.10—1995)。将平衡水分后的20.0 g浆料与650 mL去离子水、5 mL冰醋酸以及6.0 g亚氯酸钠充分混合,移入1 000 mL锥形瓶中,摇匀,扣上25 mL锥形瓶,置于75 ℃恒温水浴中加热2 h,反应1 h后加入一次冰醋酸和亚氯酸钠。定期摇动锥形瓶以确保试剂完全反应。反应结束后,将浆料放入冰水浴中冷却,并用去离子水反复洗涤至滤液不呈酸性为止,再用丙酮洗涤3次,收集浆料并置于4 ℃平衡水分。
1.2.3 酶预处理
将浆料置于50 ℃和200 r/min的条件下在50 mmol/L柠檬酸-柠檬酸钠缓冲液(pH 5.2)中进行酶预处理,底物质量分数为5.0%。预处理期间内切葡聚糖酶的用量为5 mg/g纤维底物,反应时间为1.5 h。反应结束后,将浆料置于95 ℃ 的水浴中10 min使酶灭活,离心收集酶解液用于后续检测分析。酶预处理后的浆料用去离子水洗涤至中性并置于4 ℃平衡水分。与内切葡聚糖酶相比,木聚糖酶的用量为5 IU/g纤维底物,反应时间为0.5 h,其他操作同内切葡聚糖酶的处理。内切葡聚糖酶和木聚糖酶预处理的浆料分别命名为EP和XP,未进行酶预处理的浆料命名为BP。
1.2.4 机械纳米原纤化处理
分别将酶预处理和未经酶预处理的浆料稀释至质量分数为1.0%,经超微粒研磨仪进行研磨制备LCNF。具体工艺:在磨盘间隙为0和-20 μm 下依次研磨5次,转速为1 500 r/min;磨盘间隙调至-50和-80 μm,依次研磨10次,转速为2 000 r/min;磨盘间隙再调至-100 μm研磨15次,转速为2 000 r/min。得到内切葡聚糖酶/机械处理、木聚糖酶/机械处理和纯机械处理的悬浮液分别命名为E-LCNF、X-LCNF和N-LCNF。
1.2.5 测试与表征
1.2.5.1 组分分析
BP、EP和XP样品的木质素含量及酶解液中糖分组成,根据改进文献[7]的分析方法进行。
称取0.3 g干燥至恒重的样品,在30 ℃用3 mL 质量分数72%的H2SO4水解1 h,每10 min 搅动一次,使其水解反应均匀。反应结束后,将反应混合物转移到耐压瓶中,加入84 mL去离子水,并在121 ℃下高压保温1 h。将反应后的液体过滤,收集酸水解溶液,使用紫外-可见分光光度计从205 nm处的吸光度分析计算酸溶性木质素的含量。将剩余的固体残留物在105 ℃干燥至恒重,记录干燥固体的质量,并用于计算酸不溶性木质素的含量。
取9.3 mL去离子水加入耐压瓶中,再加入0.7 mL质量分数72%的H2SO4,最后加入10 mL 收集的酶解液,溶液在121 ℃高压保温1 h。将酶解液和酸解后的酶解液分别通过离子色谱系统分析单糖组分。
1.2.5.2 红外光谱分析
通过傅里叶变换红外光谱分析BP、EP和XP样品的氢键特征参数。使用Peak-Fit软件结合高斯分布函数研究吸收光谱(3 700~3 000 cm-1)以分析氢键含量、能量和不同氢键模型的键长。
EH=(ν0-ν)/Kν0
(1)
式中:EH为氢键能量,kJ/mol;ν0为标准游离羟基频率,3 650 cm-1;ν为样品的羟基频率,cm-1;K为常数,6.7×10-2kJ-1。
(2)
式中:R为键长,10-10m;ν0为标准游离羟基的拉伸振动频率,3 600 cm-1;ν为样品的羟基频率,cm-1。
1.2.5.3 形貌分析
将LCNF配制成质量分数为10-6~10-5的悬浮液,15 ℃超声分散5 min,取一滴超声分散好的溶液滴在云母片上,自然风干后用原子力显微镜观察其形态特征。采用Nano Measurer 1.2软件统计LCNF的直径分布。
1.2.5.4 保水值
保水值反映了纤维孔隙中的总水分,该水分与总孔隙体积密切相关,是纤维素原纤维吸水和溶胀能力或原纤化程度的量度。采用改进的TAPPI T256方法[8]进行测量,质量分数为1.0%的LCNF悬浮液在3 000g的离心力下离心30 min,离心后的样品称重,105 ℃干燥24 h至恒重。保水值由离心后底物中水分的质量与绝干LCNF质量的比值计算得出。
保水值=[(m1-m2)/m2]×100%
(3)
式中:m1和m2分别为离心后底物的质量和恒重后底物的质量,g。
1.2.5.5 X射线衍射测试
LCNF的结晶度通过X射线衍射仪使用Cu-Kα辐射确定。在40 kV、30 mA以2.0°/min在5°~60°内扫描样品。
结晶度指数=[(I200-Iam)/Iam]×100%
(4)
式中:I200为22.5°附近2θ处200晶格平面的衍射强度;Iam为18°附近2θ处非结晶区的衍射强度。
Dhkl=kλ/βcosθ
(5)
式中:Dhkl为晶体在200晶格平面上的平均微晶尺寸,nm;k为Scherrer常数,0.89;λ为Cu-Kα辐射的波长,0.154 18 nm;β为200晶格平面半峰高的宽度,rad;θ为布拉格角度,rad。
1.2.5.6 比表面积
LCNF样品的比表面积采用文献[9]中刚果红染料吸附法测定。用刚果红(底物质量分数5%~30%)处理质量分数0.2%的LCNF水性悬浮液,60 ℃振荡24 h,并在5 000 r/min离心20 min。用紫外-可见分光光度计在495 nm处检测上清液的吸光度,根据刚果红标准曲线计算上清液中的刚果红浓度。
Acell=6.024×1023m1Ad/Mr
(6)
式中:Acell为比表面积,m2/g;m1为纤维吸附的饱和值,g/kg;Ad为染料分子的面积,1.73 nm2;Mr为染料分子的相对分子质量,696.68。
1.2.5.7 悬浮液特性
LCNF样品的表面电荷密度通过粒子电荷探测仪进行测量,标准滴定液为Poly-DADMAC。Zeta电位通过流动电位法Zeta电位仪进行测定。
2 结果与讨论
2.1 酶预处理对纤维结合力的影响
根据红外光谱吸收频率偏移程度以及谱图的面积变化评价酶预处理对浆料纤维氢键特性的影响。分子间和分子内氢键的伸缩振动特征峰位于3 000~3 700 cm-1区域。其中,代表O(2)H…O(6)和O(3)H…O(5)的分子内氢键及代表O(6)H…O(3′)的分子间氢键的特征吸收波数分别在3 455~3 410 cm-1、3 375~3 340 cm-1和3 310~3 230 cm-1。氢键谱带的二阶导数可用于确定不同氢键模型的波数,对图1中吸收峰氢键区域进行分峰拟合,分析不同类型氢键的特征,得到红外分峰拟合图谱(图2)。
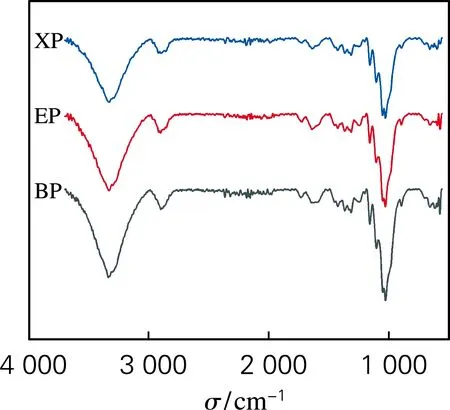
图1 BP、EP与XP的红外光谱
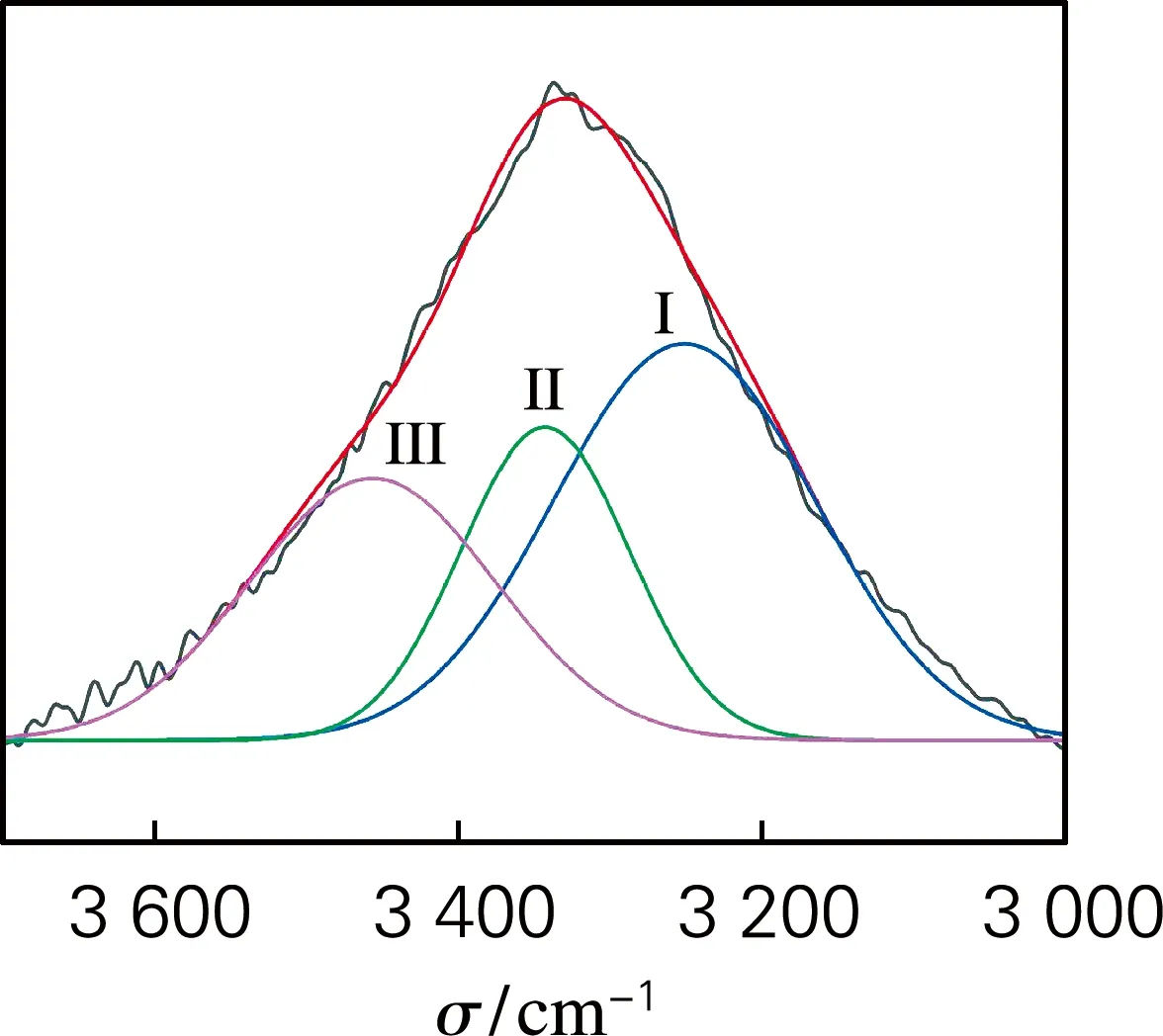
(a) BP
如表1所示,所有分峰拟合的R2均高于0.98,这表明不同氢键模型的拟合结果能够充分反映不同氢键模型的信息。结果表明,酶预处理后,纤维分子间与分子内氢键含量发生变化,纤维的氢键逐渐发生“重组”现象,说明酶预处理可以疏松纤维的结构,有利于后续纳米原纤化。

表1 酶预处理样品红外图谱氢键区域的分峰拟合
2.2 酶预处理对浆料纤维组分的影响
酶预处理过程可以减弱微纤丝之间的氢键作用力,表1结果已得到证实。酶解液中单糖及低聚糖的含量可以反映酶预处理过程对纤维复杂多层壁结构的改变,同时也进一步证实酶预处理过程可以疏松木质纤维,其中,酶解液中低聚糖由酶解液酸解后单糖的含量和组成间接反映。
经内切葡聚糖酶和木聚糖酶预处理后的浆料得率分别为92.25%和83.22%,酶解前后浆料主要化学组分组成如表2所示。由表2可以看出,内切葡聚糖酶预处理后,酶解液糖含量和酶解液酸解后糖含量都很少,说明预处理过程主要以疏松纤维为主,水解的浆料纤维较少。这是可能由于内切葡聚糖酶在纤维的非结晶区域随机水解β-1,4-葡萄糖苷键,使纤维素的非结晶区域变疏松,并较少地将纤维素或半纤维素降解为单糖或

表2 酶解液酸解前后单糖的质量分数
可溶性低聚糖。与内切葡聚糖酶处理的样品相比,经木聚糖酶处理后,酶解液及酶解液酸解液中的木糖含量明显增高。这是由于木聚糖酶能选择性地降解半纤维素中的木聚糖为单糖或可溶性低聚木糖。但该水解大多发生在微纤维之间的半纤维素中,而基层纤维致密结构之间的半纤维素却很难被直接接触或水解[10]。此外,内切葡聚糖酶处理的样品中木质素质量分数为11.21%,与未经酶预处理的样品(11.51%)相差无几,而木聚糖酶处理的样品木质素含量有所下降,其质量分数为6.08%。这是由于在植物纤维中,半纤维素与木质素之间存在化学键,构成木质素-碳水化合物复合体(LCC)[11]。因此,木聚糖酶处理在去除半纤维素成分的同时也会使一部分木质素脱落,并清楚地暴露细胞壁的原纤维网络。
2.3 酶预处理对LCNF形貌与尺寸的影响
由图3可以看出,未经酶预处理的N-LCNF呈现一种树形结构,且“树干”与“树枝”的尺寸差别较大[9],从直方图中能够直观地看到该样品的直径主要分布在20~60 nm,尺寸分布跨度大,相对分散。经过酶预处理的E-LCNF和X-LCNF都呈现相互交织的无规则网络结构。E-LCNF的直径主要分布在15~35 nm,分布相对集中,由于内切葡聚糖酶有效疏松纤维结构,机械研磨能够使纤维剥离出更细小的纤丝;X-LCNF直径主要分布在20~50 nm,分布相对较为集中,因为木聚糖酶能够降解半纤维素,破坏纤维的固有抗降解屏障性,打开纤维网络,有助于纤维纳米原纤化。由于EP的木质素含量高于XP,木质素是一种已知的抗氧化剂,它能稳定机械处理过程中形成的纤维素自由基。纤维素自由基具有极强的反应性,可以参与复合反应(交联),在木质素含量较低的情况下会不利于原纤化,在木质素含量较高的情况下,木质素清除自由基的能力导致纤维素的交联不那么明显,这使得纤维可以更好地解构[20],因此E-LCNF的直径小于X-LCNF。

图3 LCNF的AFM形貌结构及其直径分布
2.4 酶预处理对LCNF性能的影响
如表3所示,E-LCNF样品获得了最高的保水值564%,表明该样品在内切葡聚糖酶/机械研磨处理后发生显著的原纤化,纤维裸露的羟基增加,结合水分子能力增强。由于内切葡聚糖酶可以高效率和选择性地水解纤维中的非结晶区,从而使样品产生较小的纤维碎片和增加纤维素纤维在水中的溶胀度。此外,内切葡聚糖酶可能会破坏微纤维之间的氢键连接[12-13],这对从纤维表面解离出纤丝具有积极影响,也意味着可以在纤维上建立更多的水可渗透的空腔[14],从而导致保水值的增加,同时也提高了样品的比表面积。

表3 酶预处理制备所得LCNF的性能
X-LCNF的保水值显著提高,表明木聚糖酶预处理有助于“打开/松散/软化”纤维素原材料。半纤维素组合物采取高度分支的多态多糖的形式,通过氢键结合相邻的纤维素微原纤维来增强细胞壁强度,但其分支数量和组成的变化可以显著改变细胞壁强度和微观结构[15]。木聚糖酶可以使纸浆纤维中的半纤维素结构自然地解构,有益于纤维原纤化和微原纤或纳米原纤的形成[16]。
因此,木聚糖酶预处理可以破坏纤维羟基之间的分子间和分子内氢键,随后的机械原纤化增加了原纤维的内表面和外表面面积,使水分子更容易渗透到原纤维之间以增加保水值。然而,由于木聚糖酶降解了部分半纤维素,半纤维素可以抑制纤维聚合,促进纤维形成开放的、多孔的结构[17-18],且半纤维素比纤维素具有更高的水可及性[19],因此X-LCNF的保水值稍低于E-LCNF。
纤维内部的结晶区和非结晶区之间存在相互干扰。从图4中可以看出,经过酶预处理和未经酶预处理的样品在XRD谱图上相差不大,说明酶预处理过程中纤维内部结晶区域的晶型微观结构变化不显著。

图4 LCNF的X射线衍射谱图
内切葡聚糖酶预处理样品的结晶度和微晶尺寸略有增加。通过木聚糖酶预处理制备X-LCNF的结晶度增加较为明显(表3),结晶纤维素被包埋在非结晶组分中,木聚糖酶处理后,附着在纤维表面的半纤维素被去除,提高了LCNF的结晶度。
如表3所示,E-LCNF悬浮液的Zeta电位最高,说明内切葡聚糖酶酶解结合机械研磨制备的木质纤维素纳米纤丝有着良好的分散能力。这是由于内切葡聚糖酶能随机切割纤维使其变短,并破坏氢键减弱纤维内部纤丝的作用力,在机械力的作用下更易剥离出直径较小且尺寸分布均匀、比表面积大的纤维。此外,半纤维素表面带有负电荷,能够在纤维间产生排斥力[10],且侧链基团也有助于纳米纤维之间的空间排斥,因此半纤维素的存在促进了LCNF悬浮液的胶体稳定性。由于在预处理过程中木聚糖酶降解了部分半纤维素,静电排斥的作用减弱,但同时木聚糖酶酶解打开了纤维网络,在后续机械作用下原纤化程度较高,产生的纳米纤维尺寸较为均匀,综合作用下,X-LCNF的Zeta电位稍低于E-LCNF,但电荷密度最低。N-LCNF中虽然半纤维素含量高,能够提供较强的静电排斥力,但由于未经酶预处理,使得N-LCNF在机械研磨后的尺寸较大且分布不均匀,这可能导致静电排斥力不足以抵抗其倾向于凝聚的趋势,故而N-LCNF的Zeta电位在三者中最低。正如图5所示,可以直观地看到,N-LCNF悬浮液静置后会絮聚沉降,分散性较差,而E-LCNF与X-LCNF由于Zeta电位绝对值都大于40 mV,表现出良好的分散性。

图5 静置5 d的LCNF分散液
3 结 论
采用酶预处理结合机械研磨的方法制备了不同特性的LCNF样品,探究了酶预处理过程对LCNF性能的影响。结果表明,内切葡聚糖酶在预处理过程中对浆料的组分改变较为细微,得率高,随机水解非结晶区纤维疏松了木质纤维微观结构,同时也减弱了纤维间的作用力,在机械力剥离与切断的作用下原纤化明显,尺寸小且分布均匀。因此,E-LCNF的保水值、比表面积以及Zeta电位最高。木聚糖酶预处理时降解了半纤维素,同时也使部分木质素脱落,导致得率相对较低但软化了纤维原料,纳米原纤化后所得样品X-LCNF 具有最高的结晶度,保水值、比表面积以及Zeta电位均只稍低于E-LCNF。机械法所得样品N-LCNF由于未经预处理,其尺寸大且分布很不均匀,保水值、比表面积以及Zeta电位最低。总之,内切葡聚糖酶可以疏松木质纤维结构,进而促进木质纤维纳米原纤化,提高LCNF性能;木聚糖酶可以降解半纤维素并脱落部分木质素,“打开/松散/软化”了木质纤维原材料,改善了LCNF性能。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
今日农业(2022年13期)2022-09-15
食品安全导刊·中旬刊(2022年3期)2022-04-15
——谷物β-葡聚糖的最新研究进展
粮油食品科技(2022年2期)2022-03-25
中国畜牧杂志(2022年3期)2022-03-16
食用菌(2022年1期)2022-02-14
乡村科技(2021年12期)2021-09-06
北华大学学报(自然科学版)(2021年3期)2021-07-13
中国饲料(2021年2期)2021-02-23
中国科技纵横(2020年16期)2020-11-28
