
容积激活性氯离子通道蛋白ClC-3表达下调促进H9c2心肌细胞肥大及其机制研究
2021-08-04 09:31陈梦青李艳文李咏琪李春梅
安徽医科大学学报 2021年7期
孙 宇,陈梦青,汪 帅,李艳文,喻 阳,李咏琪,李春梅
心肌肥大的主要特点是心肌细胞体积增大,易引发心律失常,最终导致心力衰竭和心肌梗死等疾病。ClC-3是电压门控氯通道基因家族成员之一,在心脏、脑等组织中广泛表达。许多研究表明,ClC-3的功能具有多效性,包括作为容积调节性氯离子通道的关键组成部分,能调节细胞体积,保护心肌细胞免受低氧、缺血或肥大时细胞体积过度增加的影响。前期研究表明异丙肾上腺素能诱导大鼠原代心肌细胞、H9c2细胞和C57小鼠ClC-3表达降低,且AAV-9介导的ClC-3过表达小鼠能明显抑制异丙肾上腺素诱导的心肌肥厚。该研究利用siRNA转染H9c2心肌细胞构建ClC-3表达下调模型,观察其对心肌细胞的影响及机制,为研究ClC-3在心脏中的功能提供借鉴,同时也为临床心肌肥大的防治提供思路。
1 材料与方法
1.1 细胞与主要试剂
H9c2细胞购自上海中国科学院细胞库。异丙肾上腺素(ISO)购自美国Sigma公司。ClC-3一抗购自英国Abcam公司。siRNA ClC-3及转染试剂盒购自苏州瑞博生物有限公司。Bradford蛋白浓度试剂盒、JC-1线粒体膜电位(mitochondrial membrane potential,MMP)试剂盒、细胞骨架染色α-Tubulin一抗均购自北京碧云天生物技术有限公司。DMEM培养基、FBS胎牛血清、0.25%胰酶均购自上海Gibco公司。TRIzol购自大连TaKaRa生物公司。PCR引物由上海生工生物工程有限公司设计并合成。Prime ScriptTMRT Reagent及SYBR PCR Mix购自日本ToYoBo公司。1.2 主要方法
1
.2
.1
细胞培养及分组 大鼠H9c2心肌细胞株在含10%的胎牛血清的DMEM培养基中培养,每2 d更换1次。在37 ℃、5% CO的培养箱中培养传代,取对数期、生长良好的细胞进行实验。将2×10个细胞接种于6孔板中,待细胞融合度达到60%~70%时进行转染。加入小干扰RNA在培养箱中转染48 h和72 h后,更换为完全培养基继续培养48 h,采用qRT-PCR和Western blot检测转染效率。实验主要分为4组:空白组、siRNA空载对照组、肥大模型组:ISO(10 μmol/L诱导心肌肥大)以及siRNA转染敲低ClC-3组。1
.2
.2
Western blot检测ClC-3蛋白表达 H9c2细胞经100 nmol/L的siRNA转染不同时间后,提取蛋白,并用Bradford法测定蛋白浓度。加入上样缓冲液后,用SDS-PAGE电泳分离蛋白样品,转膜后用奶粉封闭2 h。4 ℃孵育一抗,过夜后室温孵育二抗。采用ECL化学发光法检测。各组蛋白均以GAPDH作为内参校正。1
.2
.3
α-
Tubulin免疫荧光染色 将生长状态良好的H9c2细胞经胰蛋白酶消化后,接种于带有细胞小圆玻片的48孔板中,每孔200 μl,计数后调节细胞浓度约8×10个/孔,培养24 h细胞贴壁后,进行饥饿处理12 h,细胞换液按分组分别处理48 h后,弃去培养基,PBS洗3次,每次5 min,用4%的多聚甲醛固定15 min,弃固定液,PBS洗3次,室温封闭1 h,然后加入α-Tubulin(1∶100)一抗4 ℃过夜。第2天加入羊抗鼠二抗室温避光孵育1 h,DAPI染色,PBS洗3次后封片,荧光显微镜拍照,用Image Pro Plus 6.0进行图像处理和分析。1
.2
.4
总RNA提取和qRT-
PCR检测mRNA表达水平1
.2
.4
.1
mRNA提取 细胞按2×10个/孔的密度接种于6孔板中,每孔给予每组相应处理后加入TRIzol试剂裂解细胞。用细胞刮刮下细胞,收取细胞悬液于1.5 ml EP管中,振荡1 min,使细胞完全裂解。加入氯仿,剧烈振荡15 s,混匀后室温放置3 min,12 000 r/min,4 ℃离心15 min,取上清液加异丙醇,颠倒混匀,室温静置10 min,12 000 r/min,4 ℃离心10 min,弃上清液,加75%的乙醇进行洗涤,7 500 r/min,4 ℃离心5 min,弃上清液,倒扣EP管在洁净的滤纸上,室温静置20 min。加入10 μl的DEPC水冰上溶解,吹打混匀后分光光度计测定RNA浓度和纯度。1
.2
.4
.2
qRT-PCR检测 按逆转录试剂盒(QPS-201)说明书反转录后,以GAPDH为内参,RT-PCR检测心肌组织ANP、BNP、β-MHC的相对表达量。引物序列如下:ANP上游引物5′-TACGAGACAGTGACGGACAA-3′,下游引物5′-GAAGAAGCCCAGGGTGAT-3′;BNP上游引物5′-CAGCTCTCAAAGGACCAAGG-3′,下游引物5′-CGATCCGGTCTATCTTCTGC-3′;β-MHC上游引物5′-GGGTATCCGCATCTGTAGGA-3′,下游引物5′-CCTTTCCGGCTATCAATGAA-3′;ClC-3上游引物5′-AGACATTGCTGCTGACTGGA-3′,下游引物5′-CCCTCTCTTCAAACGTCGTC-3′;GAPDH上游引物5′-AGGAGTAAGAAACCCTGGAC-3′,下游引物5′-CTGGGATGGAATTGTGAG-3′。PCR反应体系为SYBR MIX 10 μl、上游引物 1 μl、下游引物 1 μl、cDNA 2 μl、ddHO 6 μl。根据2法计算各组相对mRNA的含量。1
.2
.5
JC-1染色 接种H9c2细胞于48孔板中,1×10个/孔,待细胞贴壁后按1.2.1进行分组及给药后,按试剂盒说明书进行操作,荧光显微镜下进行拍照,并用Image J软件进行分析。1
.2
.6
容积调节性能力检测 在观察不同渗透压条件下细胞的容积调节性能力(RVD)时,用等渗液(ISO)灌流细胞5 min,改换低渗液灌流20 min,再等渗液灌流5 min。细胞玻片安放在浴槽中,用数字式摄像机连接相差显微镜与计算机,调节好显微镜焦距及亮度后,拍摄图像,采用Image Pro Plus软件进行处理。1
.2
.7
全细胞膜片钳记录ICl电流 该实验使用芯片膜片钳技术检测H9C2细胞膜上氯电流的变化。膜片钳技术用于记录电压钳位条件下一小片膜中的电流,将细胞悬浮液置于芯片上。然后,通过气泵施加压力进行抽吸,将单个细胞吸引并定位在孔上。通过向孔施加负压来实现膜的密封和破裂以建立整个电池构造。在NPC系统中,压力由PatchControl软件控制。使用此软件,可以运行预定义的协议,仪器收集的电流信号由EPC 10 USB膜片钳放大器处理后呈现于显示屏并通过程序记录下来。观察并记录等渗条件下的稳定电流5 min,接着将细胞外环境的等渗液换为低渗液,待细胞电流稳定后,持续记录稳定电流5 min,最后将细胞外环境的低渗液换为等渗液,记录全程的电流变化。
2 结果
2.1 siRNA转染H9c2心肌细胞后ClC-3 mRNA及蛋白的表达
图1显示,与空白组比较,siRNA空载对照组并不能影响细胞内ClC-3 mRNA和蛋白水平,而siRNA转染H9c2细胞48 h后,使细胞内ClC-3 mRNA降低(F
=6.35,P
<0.05)但并没有引起细胞内ClC-3蛋白水平的变化(F
=2.77,P
>0.05);siRNA ClC-3转染72 h后,能诱导细胞内ClC-3 mRNA和蛋白水平都降低(F
=52.84,P
<0.01和F
=70.93,P
<0.01)。结果表明,siRNA转染H9c2细胞72 h后可以成功构建ClC-3表达下调模型。
图1 siRNA ClC-3抑制H9c2细胞ClC-3表达
2.2 ClC-3表达下调对H9c2心肌细胞的影响
该研究分别采用α-tubulin(一种细胞骨架蛋白)进行荧光染色和qRT-PCR的方法来观察ClC-3表达下调对H9c2心肌细胞表面积和细胞内几个常见的心肌肥大标志因子ANP、BNP和β-MHC mRNA的表达水平的影响。如图2所示,ISO(10 μmol/L)刺激H9c2细胞48 h后,心肌细胞的表面积增加[ISO组vs
空白组,(2 130.40 ± 369.78)vs
(877.33 ± 109.88),F
=11.33,P
<0.01,图2A、B],且ANP、BNP和β-MHC mRNA的表达水平降低(F
=9.38、597.9、86.51,均P
<0.01);而与siRNA空载对照组不同,siRNA诱导的ClC-3敲低组的细胞表面积增加[ClC-3敲低组vs
空白组,(2 127.33 ± 109.62)vs
(877.33 ± 109.88),F
=4.93,P
<0.01],ANP、BNP和β-MHC mRNA的表达水平升高(F
=12.8、1 088、63.6,P
<0.01)。
图2 ClC-3表达下调可使H9c2细胞产生心肌肥大
2.3 ClC-3表达下调对H9c2心肌细胞线粒体功能的影响
JC-1是一种广泛用于检测MMP△Ψm的理想荧光探针,红色荧光代表JC-1聚集体(即MMP较高),绿色荧光代表JC-1单体(即MMP较低),绿色荧光与红绿荧光强度的比值越高说明MMP越高;线粒体电子传递链抑制剂CCCP(CON+)被用作阳性对照。如图3所示,与空白组相比,siRNA 空载组不能引起MMP发生变化(F
=3.26,P
>0.05),而与CCCP(CON+)(F
=64.5,P
<0.01)和ISO(F
=30.49,P
<0.01)处理一样,siRNA ClC-3能使MMP下降(F
=280.5,P
<0.01),说明ClC-3表达下调能诱导线粒体结构和功能损伤。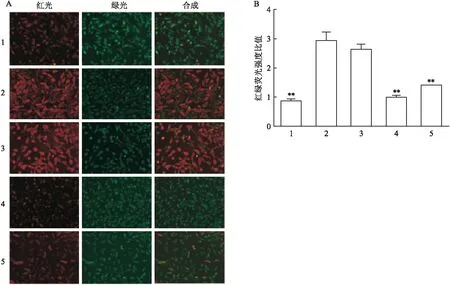
图3 细胞内线粒体膜电位的变化
2.4 ClC-3表达下调对H9c2心肌细胞容积激活性氯离子电流的影响
与空白组相比,空载siRNA处理后不会引起细胞容积激活性氯电流的改变(F
=17.25,P
<0.01),而ISO诱导与ClC-3表达下调都可抑制细胞容积激活性氯离子电流(F
=7.72,P
<0.01)。结果显示,ClC-3可能通过调节容积激活性氯离子电流调控H9c2心肌细胞的表面积大小。见图4。
图4 ClC-3表达下调对H9c2细胞容积激活性氯离子电流的影响
2.5 ClC-3表达下调对H9c2心肌细胞容积调节能力的影响
如图5所示,与空白组相比,ISO诱导H9c2心肌细胞肥大后,H9c2细胞的容积调节性能力下降(F
=1.49,P
<0.01);而空载siRNA不引起细胞容积调节能力的改变(F
=4.59,P
>0.05);siRNA转染敲低ClC-3组与ISO组类似,也可抑制细胞容积调节能力(F
=2.67,P
<0.01)。以上结果表明ClC-3表达下调导致H9c2细胞的肥大可能与其下调细胞的容积调节性能力有关。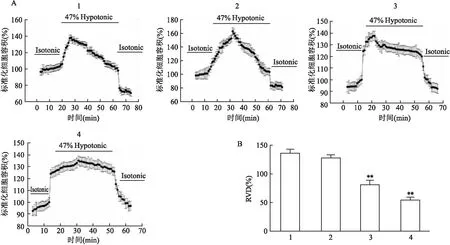
图5 ClC-3表达下调对H9c2细胞RVD的影响
3 讨论
心肌肥大的特征是长期高血压引起的室壁厚度增加,是心血管疾病的主要危险因素,严重威胁到人类健康。它常会引起心脏收缩功能障碍、心肌间质的纤维化以及胎儿基因ANP、BNP、β-MHC的等效应分子的表达增加。ClC-3氯通道是容积调节性氯离子通道基因家族成员之一,在各类动物体内广泛表达。参与到细胞兴奋、迁移、增殖、分化、凋亡和体积稳态的过程中。研究表明ClC-3在主动脉缩窄术诱导的心肌肥厚及心力衰竭中起着重要作用。因此,ClC-3可能具有调控心肌肥大的作用。该研究在前期研究基础上进一步采用siRNA转染H9c2心肌细胞72 h成功构建了ClC-3表达下调的细胞模型。ClC-3下调的同时能诱导H9c2心肌细胞表面积增加,心肌肥大标志性因子ANP、BNP和β-MHC mRNA的表达水平升高,表明ClC-3表达下调能诱导心肌细胞肥大。这与前期研究在大鼠原代心肌细胞、H9c2细胞和C57小鼠以及AAV-9介导的ClC-3过表达小鼠的结果是一致的。同时该研究还通过与计算机连接的显微镜和膜片钳技术检测了细胞RVD能力和细胞容积激活性氯离子电流。结果显示,ClC-3下调降低细胞的RVD能力和细胞的容积激活性氯离子电流,这些可能与其诱导心肌细胞肥大具有一定关系。
心脏是机体的重要器官,线粒体约占心肌细胞质量的30%,心脏空白运转所需的能量约95%来自线粒体。因此,线粒体功能与心脏功能息息相关,线粒体在细胞内信号传导、细胞死亡等方面发挥着重要作用。有报道显示,由于线粒体功能受损导致的能量生成障碍将导致心脏代谢疾病。有研究表明,肥大性心肌病中也观察到心脏线粒体肿胀、嵴断裂、呼吸链复合物活性降低。MMP作为重要的生物能量指标,能够反映线粒体的功能。该研究表明这与ISO处理相似,ClC-3下调也能诱导MMP降低,而导致线粒体功能损伤,这可能也与其能诱导心肌细胞肥大有关。
